BAB II Tinjauan Pustaka
advertisement

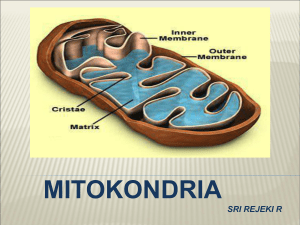
BAB II Tinjauan Pustaka Pada Bab II diberikan penjelasan mengenai beberapa hal yang berkaitan dengan penelitian ini, yaitu mengenai mitokondria, fungsi mitokondria, genom DNA mitokondria manusia, sifat genetik mitokondria, laju mutasi mtDNA, haplogroup, analisis variasi mtDNA manusia dengan metode RFLP, program Mito Mutation Analyzer (MMA) dan basis data. II.1 Mitokondria Asal-usul mitokondria diduga merupakan bakteri yang mirip dengan Rickettsia yang hidup bebas kemudian ditelan oleh nenek moyang sel eukariot membentuk endosimbiosis sekitar satu milyar tahun yang lalu. Sel inang menyediakan nutrien yang kaya energi bagi mitokondria, sedangkan sel mitokondria menjadi alat pengubah nutrien tersebut menjadi energi menggunakan oksigen. Ketika komposisi atmosfer purba bergeser dari yang kaya hidrogen menjadi kaya oksigen dengan kehadiran fotosintesis maka sistem simbiosis ini menjadi paling efektif (Albert et al., 1994) Mitokondria merupakan salah satu organel yang terdapat pada sel eukariot, berbentuk elips dengan diameter ~ 0,5 µm dan panjangnya 0,5-1,0 µm. Mitokondria memiliki dua membran yaitu membran dalam dan membran luar. Membran luar mengandung sejumlah protein transport yang disebut protein porin yang mampu menyaring ion-ion atau molekul-molekul kecil berukuran 5 kDa atau lebih lebih kecil dari itu. Selain itu membran luar juga mengandung enzim yang terlibat dalam biosintesis lipid ke matriks untuk menjalani proses β-oksidasi menghasilkan asetil CoA. Membran dalam memiliki struktur yang berlipat-lipat membentuk suatu krista. Struktur krista ini meningkatkan luas permukaan membran dalam sehingga meningkatkan kemampuannya dalam memproduksi ATP. Membran dalam mengandung protein yang terlibat dalam reaksi fosforilasi oksidatif, ATP sintetase yang berfungsi membentuk ATP pada matriks mitokondria, serta protein transport yang mengatur keluar masuknya metabolit dari matriks melalui membran. Matriks mitokondria merupakan tempat berlangsungnya reaksi-reaksi penting bagi sel, seperti siklus Krebs, reaksi oksidasi dan reaksi β-oksidasi, oleh sebab itu sebagian besar protein mitokondria terdapat pada matriksnya. Enzim-enzim yang terlibat pada reaksi-reaksi di atas dan enzim komplek piruvat dehidrogenase berperan penting dalam mengubah piruvat menjadi asetil CoA. Di dalam matriks mitokondria juga terdapat materi genetik yang dikenal dengan DNA mitokondria (mtDNA), ribosom, Adenosine triphosphate (ATP), Adenosine diphosphate (ADP), fosfat inorganik serta ion magnesium, kalsium dan kalium (Boyer et al., 1997). II.2 Fungsi Mitokondria Mitokondria berperan dalam proses metabolisme karena merupakan tempat terjadinya reaksi oksidasi, reaksi β-oksidasi dan siklus Krebs. Asam lemak hasil degradasi lipid di sitosol akan dioksidasi menjadi asetil CoA melalui reaksi βoksidasi dalam matriks mitokondria. Asam lemak di sitosol harus berikatan terlebih dahulu dengan CoA, membentuk asil CoA dengan bantuan asil CoA sintetase yang terdapat pada membran luar. Setelah masuk ke ruang antar membran, asam lemak tersebut akan terlepas dari CoA, kemudian berikatan dengan karnitin dengan membentuk karnitin-asil, dengan bantuan enzim karnitin-asil transferase I agar dapat masuk ke dalam matriks mitokondria. Selanjutnya asam lemak akan mengalami proses β-oksidasi menghasilkan asetil CoA yang kemudian akan masuk ke siklus Krebs. Asam amino hasil degradasi protein akan masuk ke siklus Krebs melalui proses konversi menjadi salah satu intermediet siklus Krebs. Aspartat dan asparagin akan dikonversi menjadi oksaloasetat, fenil alanin dan tirosin menjadi fumarat, isoleusin, metionin dan valin menjadi suksinil CoA, arginin, histidin, glutamin dan prolin diubah dahulu menjadi glutamat sebelum masuk ke siklus Krebs melalui αketoglutarat; alanin, glisin, serin, dan sistein mengalami dua macam konversi menjadi piruvat dan dari piruvat menjadi asetil CoA sebelum akhirnya sitrat yang merupakan intermediet siklus Krebs, begitu juga dengan leusin, lisin dan triptofan yang akan diubah menjadi asetoasetil CoA dan kemudian asetil CoA sebelum menjadi sitrat. Glikolisis yang merupakan tahapan dalam metabolisme karbohidrat menghasilkan piruvat di sitosol yang akan di transport menuju matriks mitokondria. Piruvat ini akan di ubah menjadi asetil CoA dan selanjutnya masuk ke dalam siklus Krebs. Reaksi-reaksi yang berlangsung dalam siklus Krebs akan menghasilkan molekul berenergi tinggi NADH dan FADH2. Selain dihasilkan dalam matriks mitokondria melalui reaksi siklus Krebs, NADH juga dihasilkan di sitosol melalui proses glikolisis. NADH di sitosol ini akan ditransport ke dalam matriks mitokondria melalui malat-aspartate shuttle (Boyer et al., 1977) Fungsi utama dari mitokondria adalah menghasilkan adenosine triphosphate (ATP). Energi yang dihasilkan ini selanjutnya oleh sel hidup digunakan sebagai “bahan bakar’ untuk menjalankan berbagai fungsi yang dibutuhkan agar dapat bertahan hidup. Proses pembentukan energi atau dikenal dengan sebagai fosforilasi oksidatif (OXPHOS) terdiri atas lima tahapan reaksi enzimatis, yang melibatkan kompleks enzim yang terdapat pada membran bagian dalam mitokondria, dimana molekul berenergi tinggi NADH dan FADH2 yang dihasilkan dari penguraian karbohidrat, lipid dan protein akan dijadikan substrat untuk selanjutnya diubah manjadi ATP. Jumlah mitokondria tiap sel tergantung jenis sel dan jenis organisme. Mitokondria ditemukan dalam jumlah banyak pada sel yang memiliki aktivitas metabolisme tinggi yaitu sel-sel kontraktil seperti sel sperma pada bagian ekornya, sel otot jantung dan sel-sel yang aktif membelah seperti sel epitelium, akar rambut dan epidermis kulit (Thorpe,1984) II.3 Genom DNA Mitokondria Manusia DNA mitokondria manusia telah ditentukan urutannya secara lengkap oleh Arderson pada tahun 1981. Urutan ini disebut Cambridge Reference Sequence (CRS), dan menjadi rujukan standar dalam menginterpretasikan variasi urutan nukleotida terutama polimorfisme mtDNA manusia yang berhubungan dengan studi antropologi maupun penentuan mutasi yang berkaitan dengan penyakit genetika (Marzuki et al., 1991). Bentuk mtDNA adalah sirkular terdiri atas untai H (heavy) memiliki basa G lebih banyak dan untai L (light ) memiliki basa C lebih banyak (Wallace et al., 1997) Ukuran genom mitokondria manusia hampir sama dengan tikus dan sapi tetapi jauh lebih kecil dari genom ragi (84kb) dan tumbuhan (~100kb) (Stracham et al., 1999). mtDNA manusia tersusun atas 16.569-bp. Genom mtDNA mengandung 37 gen yang terdiri atas 13 gen penyandi protein yang terlibat dalam fosforilasi oksidatif, terdiri atas tujuh sub unit komplek I NADH dehidrogenase yaitu ND1, ND2, ND3, ND4, ND5, ND6 dan satu unit kompleks III sitokrom b: cyt b tiga sub unit kompleks IV sitokrom oksidase terdiri atas COI, COII, COIII, dan dua sub unit kompleks V ATP sintetase yaitu ATP 6 dan ATP 8 dari 70 sub unit kompleks enzim respirasi dan daerah D-Loop (Anderson et al., (1981), Horai et al., (1995) dan Marzuki et al., 1991) 22 gen menyandi tRNA (satu gen tRNA untuk masing-masing asam amino dan dua tRNA ekstra : tRNAleu dan tRNAser), dan 2 gen menyandi rRNA (12S rRNA dan 16S rRNA) (Arderson et al., 1981). Gambar organisasi dan struktur fungsional dari mtDNA manusia dapat dilihat pada Gambar II.1 Gambar II.1 Struktur dan organisasi fungsional mtDNA manusia. Genom mitokondria manusia berukuran 16.569 pb yang mengandung 37 gen terdiri atas 13 gen penyandi protein, 22 gen penyandi tRNA dan 2 gen penyandi rRNA (Bermisheva et al., 2003) Reaksi fosforilasi oksidatif dalam mitokondria meghasilkan kurang leabih 90% energi pada organ dan sistem jaringan. Proses fosforilasi oksidatif menghasilkan berbagai metabolit berupa radikal bebas, yang berpotensi merusak DNA. Dalam kondisi normal radikal bebas akan dieliminasi oleh dismutase, katalase dan peroksidase. Namun mekanisme pertahanan ini berkurang fungsinya dengan bertambahnya umur. Organisasi genom mitokondria sangat berbeda dengan genom inti yaitu sederhana, sangat kompak, tidak adanya intron, gen-gen yang berdekatan secara umum overlap satu basa sehingga basa terakhir dari suatu gen merupakan basa pertama untuk gen berikutnya (Lewin, 1997). Kodon terminasi yang tidak lengkap pada proses transkripsi mtDNA dapat disempurnakan dengan poliadenilasi mRNA dengan enzim penambah poli A, sehingga diperoleh kodon stop UAA pada ujung 3’ (Mantoya et al., 1981 dan Brown, 1988). Tidak seperti DNA inti, mtDNA tidak dibungkus oleh protein histon sehingga memudahkan mtDNA untuk mengalami mutasi. II.4 Sifat Genetik Mitokondria Mitokondria memiliki sifat genetik sendiri yang disebut DNA mitokondria (mtDNA) yang terletak pada matrik semi cair di bagian paling dalam mitokondria. Satu mitokondria dapat mengandung puluhan mtDNA. Sistem genetik mitokondria mirip dengan bakteri yaitu berupa molekul sirkuler seperti plasmid atau kromosom bakteri, sehingga tahan terhadap eksonuklease. Kesamaan adalah terdapat sedikit DNA yang tidak menyandi protein dan gen biasanya dipaketkan secara padat dalam satu kromosom dengan sedikit intron. Hal ini berbeda sekali dengan DNA inti sel eukariot yang tersebar dalam kromosom dan memiliki banyak intron. Tipe ribosom dalam mtDNA mirip dengan sel prokariot yaitu 70S sedangkan DNA inti sel eukariot memiliki tipe ribosom 80S. Tiga hal pokok yang mendasari perbedaan antara sistem genetik inti dengan sistem genetik mitokondria yaitu tingkat polimorfisme yang tinggi yang ditunjukkan dengan laju mutasi yang tinggi bila dibandingkan dengan laju mutasi yang terjadi pada DNA inti, pola pewarisannya yang spesifik yaitu melalui garis keturunan ibu tanpa disertai dengan adanya rekombinasi mtDNA dari garis keturunan ayah, memiliki sistem kode genetik yang berbeda dengan sistem kode genetik DNA inti (kode genetik universal ) (Anderson et al., 1981 dan Lewin et al., 1997). Perbedaan kodon mtDNA manusia dengan kodon universal diperlihatkan pada Tabel II.1 Tabel II.1 Perbedaan kodon mtDNA manusia dengan kodon universal. Terdapat perbedaan antara kodon universal dengan kodon mtDNA manusia (Fox, 1987). Kodon Universal Mamalia UGA AUA AUU AGG AGA Stop Isoleusin Isoleusin Arginin Arginin Triptofan Metionin Metionin Stop Stop II.5 Laju Mutasi mtDNA DNA mitokondria bersifat unik, berbeda dengan DNA inti karena mtDNA diwariskan melalui garis keturunan ibu. Sel telur memiliki jumlah kopi mtDNA yang tinggi (≥100000) sementara sel sperma memiliki jumlah kopi yang rendah (1001500) (Chen et al., 1995dan Manfredi et al., 1997 ) dan terdapat paling banyak pada ekornya. Pada saat terjadi pembuahan pada sel telur, bagian ekor sperma dilepaskan sehingga hanya sedikit atau hampir tidak ada mtDNA yang masuk ke dalam sel telur. Karena tidak terjadi rekombinasi, maka mtDNA bersifat haploid, diturunkan dari ibu ke seluruh keturunannya. DNA mitokondria juga bersifat unik dan berbeda dengan DNA inti karena memiliki laju mutasi yang tinggi yaitu sekitar 5-10 kali DNA inti (Wallace et al., 1992). Tingginya laju mutasi mtDNA disebabkan oleh enzim DNA polymerase γ yang digunakan dalam proses replikasi mtDNA yang tidak memiliki mekanisme reparasi yang efisien (prooffreading) sehingga tidak dapat mengoreksi kesalahan-kesalahan selama proses replikasi (Watson et al., 1997). Selain itu mtDNA tidak memiliki protein pelindung seperti histon dan terletak berdekatan dengan membran dalam mitokondria, tempat berlangsungnya reaksi fosforilasi oksidatif yang menghasilkan radikal bebas sebagai produk samping (Richter, 1988). Perbedaan sifat mtDNA dengan DNA inti ditunjukkan pada Tabel II.2 Tabel II.2 Perbedaan karakteristik DNA inti dan DNA mitokondria manusia.Terdapat perbedaan antara DNA inti dengan mtDNA manusia dan perbedaan tersebut karakteristik (Sudoyo, 2003) Karakteristik DNA inti DNA mitokondria Ukuran 3x109 pb 16569 pb Kopi /sel 1 Bisa >1000 Struktur Linear Sirkular Pewarisan Paternal & maternal Maternal Rekombinasi Ya Tidak Laju mutasi Rendah Tinggi DNA polymerase yang dimiliki oleh DNA mitokondria adalah polymerase γ, yang tidak mempunyai aktivitas proofreading (eksonuklease). Tidak adanya aktivitas ini menyebabkan mtDNA tidak memiliki sistem reparasi yang dapat menghilangkan kesalahan replikasi. Replikasi mtDNA yang tidak selalu akurat ini akan menyebabkan mutasi mudah terjadi. Mutasi ini akan diturunkan dari satu generasi ke generasi selanjutnya sehingga makin jauh hubungan kekerabatan antara dua individu, makin besar pula jumlah perbedaan mutasi. Variasi basa atau polimorfisme yang disebabkan oleh mutasi ini disebut dengan Single nucleotide Polymorfism (SNP). SNP yang dapat terjadi pada daerah pengode maupun daerah yang tidak mengode, misalnya pada daerah D-Loop, dapat digunakan untuk membedakan satu individu dengan individu lain. Polimorfisme pada daerah D-Loop lebih tinggi daripada polimorfisme pada daerah pengode. Hal ini disebabkan karena laju mutasinya yang lebih tinggi. II.6 Haplogroup Haplogroup adalah cara pengelompokan berdasarkan perbedaan panjang pendeknya fragmen mtDNA yang dipotong oleh enzim restriksi. Pengelompokan haplogroup berdasarkan enzim restriksi juga telah berhasil mengelompokkan suku-suku bangsa atas haplogroup tertentu seperti haplogroup L (L1, L2, L3) termasuk kedalam group garis keturunan sub Sahara Afrika. Haplogroup H, I, J, K,T, U, V, W hampir semua mtDNA nya berasal dari Eropa, Afrika Utara, Caucasia dan Asia Barat, sedangkan haplogroup A, B, C, D, E, F, G adalah mayoritas garis keturunan dari Asia, Oceania dan Amerika asli (Nicole et al., 2001 ). Haplogroup DNA mitokondria manusia dalam bentuk huruf yaitu haplogroup A, B, C, D, E, F, G, H, I, J, K, L, T, U, V, W, dan X. Pembagian haplogroup mtDNA berdasarkan populasi genetik dan letak georafis antara lain, Eurasia Barat di kelompokkan dalam haplogroup H, T, U, V, X, K, N, I, J. Sub Afrika Sahara atas haplogroup L, sedangkan Asia Timur termasuk ke haplogroup A, B, C, D, E, F, G (C, D, E dan G termasuk ke haplogroup M) dan Amerika asli termasuk ke dalam haplogorup A, B, C, D (Bermisheva, et al., 2003) II.7 Analisis Variasi mtDNA Manusia dengan Metode RFLP Penelitian mengenai variasi mtDNA manusia dilakukan dengan analisis restriksi menggunakan metode RFLP. RFLP suatu metode analisis urutan DNA yang di potong oleh enzim restriksi yang diketahui, dengan mengetahui perbedaan panjang fragmen yang dipotong oleh enzim restriksi tidak sama, berarti enzim restriksi memotong DNA pada lokasi yang tidak saling berhubungan. Perbedaan dan kemiripan ini dapat digunakan untuk mendiferensiasi jenis, suku bangsa dan strain antara satu individu dengan individu lain. Enzim restriksi endonuklease yang digunakan dalam metode RFLP antara lain adalah HaeIII, AluI, HincII, HhaI, HaeII, HpaI, RsaI, MboI, AvaII, BamHI, NIaIII, Hinf I, TaqI dan AccI (Horai et al., 1986) Berbagai penelitian telah dilakukan terhadap polimorfisme mtDNA manusia. Variasi urutan nukleotida mtDNA telah ditentukan menggunakan enzim restriksi, dari 140 manusia Afrika yang menunjukkan bahwa lebih dari 76% mtDNA manusia Afrika dibentuk dalam satu kelompok besar yaitu haplogroup L, yang didefinisikan sebagai manusia Afrika spesifik pada 3592 pb untuk sisi restriksi Hpa I. Mutasi tambahan sebagai sub bagian haplogroup L yaitu, adanya insersi 12 pb homoplasmi pada daerah antara gen tRNAtyr dan gen COI yang diamati pada 17.6% suku Agmies dari Republik Afrika tengah, 25% lainnya dari suku tersebut mempunyai delesi 9 pb antara gen COII dan tRNAlys (yang digunakan untuk menentukan hubungan genetika antara manusia Asia dan Pasifik). Sub haplogroup lainnya diamati pada manusia Senegal yang mirip dengan manusia Eropa dan Asia. Perbandingan polimorfisme antara populasi Afrika dan non Afrika menunjukkan bahwa populasi Afrika mempunyai populasi yang lebih besar. Hasil penelitian ini dapat mendukung suatu hipotesis bahwa manusia Afrika, merupakan nenek moyangnya manusia yang mewakili manusia purbakala dan manusia moderen. Usia populasi Afrika total sekitar 101000-133000 tahun yang lalu, sedangkan usia haplogroup L sekitar 98000-130000 tahun yang lalu ( Chen et al., 1995) Analisis delesi 9 pb pada daerah antara gen COII dan tRNAlys dapat digunakan untuk mengindentifikasi hubungan genetik antara manusia Asia Tenggara dengan Polinesia. Delesi 9 pb tersebut ditemukan pada 74 manusia Indonesia, Papua New Guinea, Amerika Samoa (Redd et al., 1995). Sembilan di populasi Asia Tenggara (Taiwan selatan, Filipina dan Indonesia), tetapi tidak ditemukan pada manusia India, Bangladesh dan Pakistan (Melton et al., 1995) II.8 Program Mito Mutation Analyzer (MMA) Program Mito Mutation Analyzer (MMA) adalah algoritma pemograman yang dirancang untuk menganalisis dan menampilkan jumlah, jenis dan posisi mutasi satu atau lebih sampel urutan lengkap mtDNA manusia sekaligus. Algoritma tersebut dituliskan dalam bahasa C++. Program MMA membaca urutan lengkap mtDNA dalam file berformat. txt dan .seq, kemudian menuliskan hasil analisis, berupa panjang basa urutan lengkap, jumlah, jenis, dan posisi mutasi dalam file yang sama (Yolita, 2006) II.9 Basis Data FBI telah mengumpulkan data-data urutan nukleotida DNA mitokondria manusia di dunia yang tersimpan dalam software mtDNA Population. Data-data ini tersimpan dalam beberapa situs yaitu [www. Mitomap.org] dan Mitbase, Genbank dari National Institute of Health (NIH), National Center for Biotecnology Information (NCBI), DNA Data Bank of Japan (DDBJ) dan sekuen mtDNA disamping DNA inti berbagai organisme (Baxevanis, 2001). Data ini dapat diakses secara gratis. Data mtDNA FBI dikelompokkan menjadi dua kelompok yaitu data forensik dan data publik. Sesuai dengan kelompoknya, data forensik digunakan dalam identifikasi forensik yang menggunakan mtDNA, sedangkan data publik mencakup urutan nukleotida mtDNA yang dikumpulkan, diorganisasikan dalam literatur ilmiah. dikelompokkan, diformat dan