G12iim_BAB IV Pembahasan
advertisement

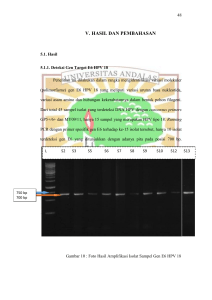
Analisis Varian Nukleotida Pada ekson 2, satu dari enam titik varian termasuk substitusi transversi. Varian tersebut yaitu C1G yang menyandikan asam amino leusin. Lima titik varian lainnya termasuk substitusi transisi yaitu C41T (Ala), G239A (Arg), G266A (Arg), A268G (Thr), dan A331G (Met). Pada ekson 3, satu titik varian termasuk substitusi tranversi A330C (Ile), sedangkan lima varian lainnya yaitu G75A, G77A, T261C, C273T, dan T336C (Tyr) termasuk substitusi transisi. Analisis lebih lanjut terhadap varian tersebut menunjukkan bahwa empat dari 12 total varian merupakan varian yang hanya terjadi pada sapi madura. Varian tersebut yaitu G75A, G77A, T261C, dan C273T. Varian G75A merupakan varian yang terjadi pada posisi nukleotida 75 basa G terhadap basa A (Gambar 2A). Varian tersebut bersifat nonsense karena tetap sebagai asam glutamat. Varian G77A terjadi pada posisi nukleotida 77 basa G terhadap basa A (Gambar 2A). Varian ini bersifat missense karena menyebabkan perubahan histidin menjadi arginin. Lebih lanjut, varian T261C terjadi pada posisi nukleotida 261 basa T terhadap basa C (Gambar 2B). Varian ini tetap menyandikan asam amino alanin. Kemudian varian C273T terjadi pada posisi nukleotida 273 basa C terhadap basa T yang tetap menyandikan sistein (Gambar 2C). Delapan varian lainnya merupakan varian yang memiliki kesamaan basa nukleotida terhadap beberapa genus Bos pembanding. Varian C1G pada ekson 2 memiliki kesamaan nukleotida basa C sebesar 48% terhadap genus Bos pembanding, varian C41T memiliki kesamaan nukleotida basa C sebesar 64%, varian G239A memiliki kesamaan nukleotida basa G sebesar 88%, varian G266A memiliki kesamaan nukleotida basa G sebesar 92%, varian A268G memiliki kesamaan nukleotida basa A sebesar 96%, dan varian A331G memiliki kesamaan nukleotida basa A sebesar 92%. Sedangkan dua varian yang terdapat pada ekson 3 yaitu varian A330C memiliki kesamaan nukleotida basa A sebesar 72,7% dan varian T336C memiliki kesamaan nukleotida basa T sebesar 36,4%. Kesamaan nukleotida basa tersebut sebagian besar terjadi pada Bos taurus dan Bos indicus pembanding. PEMBAHASAN Amplifikasi dan visualisasi untuk ekson 2 menghasilkan pita tunggal yang tebal dan beberapa pita tambahan yang samar-samar (Gambar 1A). Pita tambahan tersebut merupakan pengotor yang terdeteksi dalam gel poliakrilamid setelah pewarnaan perak dan tidak sampai mengganggu runutan amplikon. Hal ini didukung dengan puncak kromatogram yang jelas dengan jarak/spasi antar puncak yang sebagian besar sama (Gambar 2). Pewarnaan sensitif perak yang digunakan untuk mendeteksi pita-pita DNA dalam gel poliakrilamid sangat sesuai dengan volume dan konsentrasi sampel yang sangat kecil yaitu 2 μl [10 ng] (Avise 1994). Hal tersebut dikarenakan pewarnaan sensitif perak merupakan metode dengan sensitifitas yang sangat tinggi dan cepat dalam mendeteksi pita-pita DNA yang sangat kecil dengan konsentrasi berkisar 10 ng (Byun et al. 2009). Hasil runutan dua amplikon dari arah forward dan reverse dengan hasil pensejajaran ruas gen miostatin ekson 2 dan ekson 3 menunjukkan adanya perbedaan panjang nukleotida. Perbedaan yang terdapat pada ekson 2 berasal dari bagian akhir intron 1 sepanjang 158 nt dan bagian awal intron 2 sepanjang 127 nt (Lampiran 1). Sedangkan pada ekson 3, perbedaan berasal dari bagian akhir intron 2 sepanjang 108 nt dan bagian awal terminator sepanjang 125 nt (Lampiran 1). Pada bagian intron-intron tersebut tidak dilakukan analisis lebih lanjut karena ruas intron tidak berperan dalam proses penyandian asam amino (translasi) dalam hal ini ekspresi dari gen miostatin. Analisis runutan gen miostatin menunjukkan bahwa ekson 2 dan ekson 3 masing-masing menghasilkan enam varian berturut-turut yaitu C1G, C41T, G239A, G266A, A268G, A331G,G75A, G77A, T261C, C273T, A330C dan T336C. Varian tersebut ditunjukkan oleh adanya dua puncak pada kromatogram yang saling tumpang tindih. Puncak kromatogram pada varian ekson 3, bukanlah sebagai pengotor namun lebih mengarah kepada varian (Gambar 2). Namun lain halnya dengan beberapa puncak yang terdapat pada ekson 2 selain enam puncak yang menunjukkan varian ekson 2. Banyaknya puncak pada latar belakang kromatogram (tidak dilampirkan) diakibatkan pengotor yang terbawa saat amplifikasi sampel. Hal ini mengakibatkan runutan ekson 2 diverifikasi lebih lanjut terhadap runutan amplikon dari arah primer forward dan reverse untuk memastikan keakuratan runutan tersebut. Adanya dua puncak pada kromatogram merupakan sebuah data kualitatif yang 4 diharapkan, karena menunjukkan adanya varian pada ekson 2 dan ekson 3. Puncakpuncak tersebut merupakan hasil dari pembacaan mesin perunutan PCR dan elektroforesis terhadap sampel yang dijadikan dalam satu tabung (DNA pooling) yang sesuai dengan masing-masing ekson. Teknik DNA pooling yang digunakan dalam penelitian ini merupakan teknik yang dapat mendeteksi varian pada suatu lokus namun varian tersebut tidak dapat diketahui terjadi pada sampel yang mana. Ini karena sampel-sampel tersebut dijadikan dalam satu tabung (pooling) sehingga teknik ini sangat hemat sebagai penanda molekuler dan proses screening pada sampel dapat terjadi secara cepat dan banyak sampel. Secara umum, teknik DNA pooling untuk perunutan merupakan teknik dalam mendeteksi delesi, insersi dan substitusi nukleotida (dikenal sebagai Single Nucleotide Polymorphisms - SNPs). Hasil perunutan amplikon ekson 2 (Lampiran 4) dan ekson 3 (Lampiran 5) menunjukkan bahwa tidak ditemukan adanya varian yang menyebabkan terjadinya pertambahan massa dan jumlah serat otot rangka (dikenal double muscling atau muscular hypertrophy). McPherron dan Lee (1997) melaporkan adanya delesi 11 pb pada ekson 3 diantara nukleotida 821-831 dikenal dengan nt821(del11), mengakibatkan stop kodon menjadi tidak dikenali sehingga protein miostatin tidak dapat diekspresikan. Sedangkan Grobet et al. (1998) melaporkan beberapa varian yang menyebabkan double muscling terjadi pada ekson 2 yaitu delesi tujuh pb dan digantikan dengan insersi 10 pb pada posisi nukleotida ke-419 dikenal sebagai nt419(del7-ins10), transisi basa C menjadi basa T pada posisi nukleotida ke-610 (Q204X), dan transversi basa G menjadi basa T pada posisi nukleotida ke-676 (E226X); serta substitusi transisi basa G menjadi basa A pada posisi nukleotida ke-938 (C313Y) menyebabkan asam amino sistein (Cys) berubah menjadi treonin (Thr). Varian yang tidak menyebabkan double muscling juga telah dilaporkan oleh Dunner et al. (2002) pada ekson 2 yaitu substitusi transisi basa G menjadi basa A pada posisi nukleotida ke-544 (D182) yang mengakibatkan perubahan asam aspartat (Asp) menjadi asparagin (Asn) dan substitusi transisi basa G menjadi basa A pada posisi nukleotida ke-387 dikenal dengan nt387(G-A). Regulasi miostatin secara umum diawali dengan pembentukan dan ekspresi sebagai premiostatin yang belum aktif. Miostatin menjadi aktif pada saat pasca-translasi dan kemudian berikatan dengan follistatin. Kompleks miostatin dan follistatin ini akan membentuk situs aktif jika berikatan dengan activin reseptor IIB di jaringan otot. Reseptor tersebut akan memberikan sinyal kerja ke protein yang mengatur pengendalian otot (miogenin), sehingga perkembangan jaringan otot menjadi normal. Mutasi pada gen miostatin menyebabkan perubahan jumlah (hiperplasia) dan/atau ukuran serat otot (hipertropi) maupun perubahan ikatan situs aktif dengan activin IIB sehingga menyebabkan fenomena double muscling (McNally 2004). Pada penelitian ini, amplifikasi dan perunutan gen miostatin menunjukkan bahwa empat varian pada ekson 3 hanya terdapat pada sapi madura, yaitu G75A, G77A, T261C, dan C273T (Tabel 1). Meskipun varian berbeda berdasarkan basa tersebut nukleotidanya, telaah lebih lanjut berdasarkan asam amino yang disandikan ternyata tiga varian, yaitu G75A, T261C, dan C273T memiliki asam amino yang sama dengan genus Bos pembanding berturut-turut yaitu asam glutamat (Glu), alanin (Ala), dan sistein (Cys). Sedangkan pada varian G77A memiliki perbedaan asam amino karena terjadi perubahan dari histidin (His) menjadi arginin (Arg). Arginin (Arg) dan histidin (His) merupakan asam amino yang bersifat basa dan mempunyai gugus amino pada rantai sampingnya yang bermuatan positif serta bersifat hidofilik. Adanya kesamaan asam amino pada tiga varian ekson 3dikarenakan posisi nukleotida basa pada tiga varian tersebut terjadi pada basa ketiga (Lampiran 5) sehingga asam amino yang disandikannya tetap. Sedangkan posisi nukleotida basa pada varian G77A terjadi pada basa kedua sehingga terjadi perbedaan asam amino yang disandikan. Asam amino yang berubah tidak hanya terjadi pada perubahan nukleotida basa di posisi kedua setiap kodonnya namun juga dapat terjadi pada basa kesatu seperti pada varian C1G, A268G dan A330C (Lampiran 4). Selain kesamaan berdasarkan nukleotida basa, lima varian ekson 2 juga memiliki kesamaan berdasarkan sifat asam amino terhadap genus Bos pembandingnya. Varian C1G, C41T, dan A331G berturut-turut menyandikan asam amino leusin (Leu), alanin (Ala), dan metionin (Met). Sedangkan genus Bos pembandingnya menyandikan asam amino valin (Val). Keempat asam amino tersebut merupakan asam amino dengan rantai samping nonpolar dan bersifat hidrofobik. 5