Respon Anatomi Daun Dan Parameter Fotosintesis
advertisement

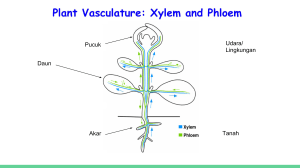
PENDAHULUAN Latar Belakang Peningkatan suhu udara atmosfir diduga akan sangat mempengaruhi iklim global dunia, seperti kemungkinan meningkatnya frekuensi dan tingkat kekeringan di beberapa belahan bumi khususnya Asia dan Afrika (Pitelka dan Rojas 2001). Keadaan ini mempengaruhi pertumbuhan dan menyebabkan penurunan produksi tumbuhan (Hamim 2005). Apalagi di daerah-daerah pertanian padi tadah hujan, keadaan kekeringan yang berkepanjangan akan menurunkan produksi beras, sehingga dapat menimbulkan kerawanan pangan. Kekeringan merupakan salah satu faktor lingkungan yang besar pengaruhnya terhadap penurunan produksi tumbuhan. Bahkan kekeringan merupakan penyebab terbesar penurunan produksi pangan dunia bila dibandingkan dengan faktor lingkungan lain (Boyer 1985). Selain menurunkan laju fotosintesis, kekeringan juga menyebabkan penurunan laju pertumbuhan akibat rendahnya potensial air dan turgor tumbuhan (Tezara et al. 2002). Secara fisiologis ada perbedaan yang menyolok antara tumbuhan C3 dan C4 dalam menghadapi cekaman kekeringan. Pada tumbuhan C3, ketika stomata menutup sebagai akibat kekeringan, fotosintesis neto akan menurun dengan cepat, sedangkan fotorespirasi akan meningkat (Drake et al. 1997). Hal ini berkaitan dengan karakteristik dari enzim fotosintetik, RuBP karboksilase (Rubisco) yang selain mengikat CO2 juga dapat berikatan dengan oksigen dalam proses fotorespirasi, dan keadaan ini terjadi khususnya ketika rasio CO2/O2 menurun akibat penutupan stomata (Hamim 2005). Tumbuhan C4 memiliki metabolisme reduksi CO2 yang berbeda dengan C3. Pada tumbuhan C4, CO2 diikat oleh enzim PEP karboksilase di ruang mesofil daun. Enzim PEP karboksilase dapat mengikat CO2 membentuk senyawa berkarbon empat, namun PEP karboksilase tidak dapat mengikat O2 (Kanai dan Edwards 1999). Senyawa berkarbon empat yang terbentuk di ruang mesofil selanjutnya dipompa ke sel seludang pembuluh. Hal ini menyebabkan sel seludang pembuluh memiliki kadar CO2 lebih tinggi, dan disini CO2 akan diikat oleh rubisko membentuk gula. Tingginya konsentrasi CO2 di sel seludang 2 pembuluh menyebabkan fotorespirasi pada tumbuhan C4 sangat rendah. Hal tersebut memungkinkan tumbuhan C4 relatif lebih tahan terhadap kekeringan dibanding tumbuhan C3 (Voznensenskaya et al. 2003). Pada kondisi intensitas cahaya yang tinggi, kekeringan mungkin juga bisa mengakibatkan terjadinya cekaman oksidatif karena dalam keadaan intensitas cahaya tinggi akan terjadi kelebihan energi dari proses reaksi terang, sedangkan laju reduksi CO2 fotosintesis menurun drastis akibat penutupan stomata selama cekaman (Violita 2007). Hamim (2004) menyatakan bahwa pada tahap awal kekeringan menyebabkan berkurangnya pembukaan stomata untuk meminimalisir kehilangan air di bawah kondisi cahaya berlebihan. Peristiwa ini mengakibatkan terjadinya penurunan konsentrasi CO2 intrasel, sehingga tumbuhan mengalami kelebihan reduksi pada transfer elektron fotosintesis (Berkowitz 1998). Kelebihan reduksi ini terjadi karena pembentukan NADPH pada reaksi terang tidak diimbangi oleh pemakaian NADPH pada reaksi gelap karena penurunan konsentrasi CO2 intrasel. Hal ini mengakibatkan terbentuknya spesies oksigen reaktif (ROS) yang diawali dengan peningkatan jumlah elektron pada transpor elektron fotosintesis oleh oksigen. Proses selanjutnya akan terbentuk berbagai bentuk senyawa ROS seperti; superoksida (O2-), singlet oksigen (.O2), radikal hidroksil (OH-) dan hidrogen peroksida (H2O2) (Mc Kersie dan Leshem 1994). Senyawa ROS ini akan dapat menimbulkan kerusakan pada tumbuhan (Aroca et al. 2001). Jika hal ini dibiarkan, maka lama kelamaan tumbuhan akan mati (Apel dan Hirt 2004). Dalam menghadapi cekaman oksidatif yang di antaranya ditimbulkan oleh cekaman kekeringan, dapat diatasi oleh tumbuhan melalui mekanisme adaptasi, baik yang berjalan secara enzimatik maupun non enzimatik. Salah satu mekanisme penyelamatan secara non enzimatik terhadap cekaman oksidatif ini yaitu dengan mengakumulasi senyawa antioksidan seperti asam askorbat (ASA). ASA merupakan asam organik yang terdapat di dalam kloroplas seperti halnya klorofil yang berfungsi menetralisir radikal oksigen yang terbentuk akibat cekaman oksidatif (Dina 2001). Selain mekanisme fisiologis, tumbuhan juga memiliki kemampuan adaptasi secara morfologis dan anatomis. Pada keadaan cekaman kekeringan 3 terdapat dua mekanisme utama yang mungkin terjadi pada tumbuhan, yaitu: (a) tumbuhan berusaha menghindari cekaman, baik dengan cara melakukan perubahan struktur morfologi dan anatomi, maupun dengan meningkatkan efisiensi penggunaan air dengan cara mengatur laju transpirasi, dan (b) meningkatkan toleransi terhadap cekaman kekeringan melalui perubahan kimia sel (Meyer dan Boyer 1981) Pada dasarnya tumbuhan memiliki mekanisme tertentu dalam menghadapi cekaman kekeringan. Informasi tentang mekanisme tumbuhan C3 dan C4 dilihat dari aspek fisiologis dan anatomis khususnya yang menyangkut perbedaan kaakteristik antara tumbuhan C3 dan C4 masih kurang. Oleh karenanya kajiankajian tentang perubahan struktur anatomi, laju fotosintesis dan perubahan fisiologi dari kedua kelompok tumbuhan tersebut saat mendapat cekaman kekeringan khususnya untuk jenis tumbuhan C3 dan C4 yang ada di Indonesia masih perlu dilakukan. Tujuan 1 Membandingkan laju fotosintesis dari parameter yang diukur yaitu: efisiensi maksimum fotosintesis (Fv/Fm), pelepasan energi untuk reaksi fotokimia (qP), pelepasan energi untuk reaksi non fotokimia (qN), dan quatum hasil fotosintesis (qY) terhadap perlakuan cekaman kekeringan pada tumbuhan C3 dan C4. 2 Mengamati respon anatomi daun pada tumbuhan C3 (padi gogo dan caisim) dan C4 (echinochloa dan bayam) pada perlakuan cekaman kekeringan. 3 Mengamati akumulasi Asam askorbat (ASA) pada tumbuhan C3 dan C4 yang mendapat cekaman kekeringan. Hipotesis 1 Terdapat perbedaan laju fotosintesis dari parameter yang diukur yakni; Fv/Fm, qP, qN, dan qY terhadap perlakuan cekaman kekeringan pada tumbuhan C3 dan C4. 2 Terdapat perbedaan respon anatomi daun terhadap cekaman kekeringan pada tumbuhan C3 dan C4. 4 3 Peningkatan akumulasi ASA pada tumbuhan C3 dan C4 yang mendapat cekaman kekeringan. Manfaat Penelitian Penelitian ini dapat diperoleh data dan informasi tentang respon anatomi daun dan fisiologi tumbuhan C3 dan C4 yang dibutuhkan dalam pengembangan tumbuhan yang toleran terhadap cekaman kekeringan.