Transformasi Nitogren dalam Tanah Tergenang
advertisement

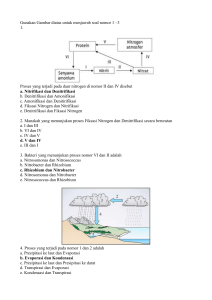
6 II. TINJAUAN PUSTAKA 2.1. Pengaruh Penggenangan Tanah terhadap Sifat-sifat Tanah Sifat-sifat fisik, kimia, dan biologi tanah tergenang sangat berbeda dengan sifat-sifat tanah lahan kering. Dalam tanah sawah, oksigen yang semakin berkurang, pembuatan teras, dan pembentukan gundukan/pematang akan mengubah sifat kimia tanahnya dan adanya genangan air di atas permukaan tanah dapat melindungi tanah dari sebagian kerusakan akibat proses -proses yang memengaruhi produktivitas jangka panjangnya, seperti erosi tanah. 2.1.1. Pengaruh Penggenangan terhadap Sifat Fisikokimia Tanah Dengan penggenangan, air memenuhi ruang pori tanah, dan udara dalam tanah dikeluarkan. Tiadanya oksigen dalam tanah maka organisme tanah akan menggunakan sumber penerima elektron lain dari komponen tanah lainnya sebagai pengganti oksigen, dan akibatnya tanah menjadi bersifat reduktif. Oksigen terlarut dalam air genangan yang berasal dari atmosfer atau dari aktivitas fotosintetik berbagai hidrofit, akan berdifusi ke lapisan tanah permukaan di bawah air genangan yang bersifat oksidatif. Walaupun lapisan yang lebih dalam dari tanah tergenang tetap tereduksi, lapisan teroksidasi tersebut sering berperan penting dalam transformasi kimia dan siklus hara yang terjadi dalam tanah tergenang. Pada lapisan tanah teroksidasi yang tipis tersebut, jasad renik melakukan proses biologi yang bersifat aerobik dan berbagai senyawa mineral berada dalam bentuk teroksidasi seperti SO 42- , NO3- , Fe3+ dan Mn4+, sedangkan pada zona anaerob yang lebih bawah didominasi oleh bentuk-bentuk tereduksi seperti senyawa fero (Fe 2+) dan mangano (Mn2+ ), ammonia, dan sulfida (S2-) (Mohanty dan Dash, 1982). Karena adanya besi feri (Fe 3+) dalam lapisan teroksidasi, tanah sering berwarna coklat atau merah kecoklatan. Sebaliknya warna sedimen tereduksi yang didominasi oleh besi fero (Fe2+) sering memberikan warna abu kebiruan sampai abu kehijauan karena adanya proses gleisasi. Sebelum perubahan ini terjadi, organisme telah mereduksi ion nitrat menjadi gas N2O dan N2, dan mangani (Mn4+) menjadi mangano (Mn2+) (Greenland, 1997). 7 2.1.2. Pengaruh Penggenangan terhadap Sifat Biokimia Tanah Konsentrasi O2 di bawah lapisan tanah teroksidasi yang tipis menurun tajam dan mendekati nol. Laju penipisan O2 ini tergantung pada suhu, ketersediaan bahan organik untuk respirasi jasad renik, dan kadang-kadang pada kebutuhan O2 dari reduktan seperti besi fero (Gambrell dan Patric k, 1978). Pada kondisi tanpa O2 jasad renik aerob mati atau menjadi tidak aktif dan jasad renik anaerob fakultatif atau anaerob obligat menjadi aktif dalam zona anaerob. Organisme-organisme ini menciptakan zona tereduksi dengan sistem potensial redoks campuran yang memengaruhi sifat kimia dan elektrokimia tanah (Mitsch dan Gosselink, 1993). Skema dari profil tanah tergenang ditunjukkan dalam Gambar 1. Akibat dari penggenangan, potensial redoks dari lapisan tanah yang tereduksi menurun tajam. Potensial redoks (Eh), suatu ukuran tekanan elektron (atau ketersediaan elektron) dalam larutan, sering digunakan untuk mengkuantifikasi derajat reduksi elektrokimia dari tanah-tanah tergenang. Oksidasi terjadi tidak hanya selama pengambilan oksigen tetapi juga bila ion hidrogen dilepaskan (misalnya : H 2S S2- + 2H +) atau, yang lebih umum bila secara kimia memberikan elektron (misal: Fe 2+ Fe 3+ + e -). Reduksi adalah proses yang berlawanan yaitu memberikan oksigen, menerima hidrogen (hidrogenasi), atau menerima elektron. Potensial redoks dapat diukur dalam tanah tergenang dan merupakan ukuran kuantitatif dari kecenderungan tanah untuk mengoksidasi atau mereduksi bahan atau komponen tanah (Faulkner dan Richardson, 1989). Tanah yang teraerasi dicirikan dengan potensial redoks +400 milivolt (mV) atau lebih besar. Bila proses reduksi cukup intens, tanah dapat mempunyai potensial redoks –300 mV. Tingkat oksidasi dan reduksi dari sistem redoks, seperti oksigen, nitrat, nitrit, mangan, besi, dan sulfur juga berbagai senyawa organik yang mudah terdekomposisi menentukan potensial redoks tanah (Qixiao dan Tianren, 1997). Dalam lapisan tanah tereduksi, untuk menggantikan oksigen yang diperlukan dalam metabolisme aerob, organisme anaerob menggunakan penerima elektron lain yang lebih lemah. Penerima elektron yang terkuat setelah O 2 adalah nitrat (NO3-). Nitrat direduksi menjadi gas N2 atau N2O pada potensial redoks 8 NH3 Kedalaman Tanah (cm) Permukaan air 0 1 NH 4, NH 4OH, NH3 (aq) 2 Permukaan tanah 3 NH4+ 4 HNO 2 HNO 3 La pisa n Tan ah Tero ks i dasi Nitrifikasi 5 pencucian 6 7 HNO 3 HNO2 HNO N 2O dan N2 Denitrifikasi 8 gas volatil 9 Lap i sa n Tan ah Tere du ks i 10 Gambar 1. Skema dari Lapisan Oksidasi – Reduksi (Sumber: Mikkelsen, 1987) +220 mV dan proses ini disebut denitrifikasi. Nitrat bersifat stabil hanya pada lapisan tanah bagian atas, yaitu di bawah genangan air dan tapak-tapak mikro yang bersifat aerob dalam lapisan anaerob seperti di sekitar perakaran tanaman padi. Daerah di sekitar perakaran padi, kira-kira tiga milimeter tebalnya, merupakan zona teroksidasi (Zhiyu et al., 1990). Kondisi teroksidasi ini terjadi karena adanya transpor oksigen melalui aerenchyma dari tanaman padi ke daerah dekat permukaan akar. Nitrat tersebut biasanya bergerak ke bawah ke dalam lapisan tereduksi melalui difusi dan aliran massa yang selanjutnya secara biologi didenitrifikasi menjadi N2 dan N2O (Mikkelsen et al., 1995). Apabila O2 dan NO3- habis terpakai, maka potensial redoks turun dan hidroksida Mn4+ dan Fe3+ akan direduks i masing-masing menjadi Mn2+ pada +200 mVdan Fe 2+ pada +120 mV. Bentuk-bentuk tereduksi dari Fe dan Mn ini mempunyai kelarutan yang lebih besar daripada bentuk teroksidasinya. Akibatnya, ketersediaan Fe dan Mn meningkat di bawah kondisi tergenang. Bila suplai penerima elektron lebih kecil daripada laju suplai elektron maka kondisi reduksi yang lebih kuat akan terjadi dan potensial redoks turun menjadi sekitar – 150 mV dan selanjutnya sulfat (SO 42-) 9 direduksi menjadi sulfida (S2-). Bila SO42- habis maka jasad renik akan menggunakan energi yang tersimpan dalam senyawa organik dengan mereduksi H+ dan H2 dan bahan organik ddekomposisi secara anaerob menjadi CO 2, asamasam organik dan alkohol. Pada kondisi sangat tereduksi dekomposisi bahan organik menghasilkan CH 4, biasanya pada nilai Eh di bawah –250 sampai –300 mV. Tanah cenderung mempertahankan nilai Eh pada selang tertentu sampai komponen tanah yang teroksidasi habis, misalnya tanah yang direduksi akan cenderung mempertahankan Eh pada sekitar +220 mV selama ada NO 3-. Bila NO3- habis, maka Eh turun dan selanjutnya terjadi reduksi penerima elektron yang lebih lemah daripada NO3-. Dengan penggenangan terjadi akumulasi N-NH4+ dan hilangnya N-NO3yang sebelumnya sudah ada dalam tanah. Dalam tanah-tanah tergenang, amonia, amin, merkaptan dan sulfida dihasilkan dari dekomposisi protein. Mineralisasi N berkorelasi positif dengan persentase C- dan N-organik dalam tanah tetapi berkorelasi negatif dengan nisbah C/N (Mikkelsen, 1987), nisbah lignin/nitrogen (L/N) (Becker et al., 1994), dan nisbah tannin/nitrogen (T/N) (Clement et al., 1995). Salah satu akibat penting dari penurunan potensial redoks setelah penggenangan adalah perubahan pH (Qixiao dan Tianren, 1997). Reaksi tanah (pH) dari sebagian besar tanah-tanah setelah penggenangan cenderung mendekati netral. Dalam sebagian besar proses reduksi yang terjadi dalam tanah, seperti reduksi oksida -oksida besi, mangan dan sulfat, terjadi konsumsi proton. Tetapi asam-asam organik dan karbon dioksida (CO2) ya ng dihasilkan selama dekomposisi bahan organik dapat memberikan proton ke dalam tanah, yang menyebabkan penurunan pH. Oleh karena itu, arah dan besarnya perubahan pH tanah selama penggenangan ditentukan oleh jumlah relatif proton yang dikonsumsi dan dilepaskan. Dalam tanah sawah yang masam, pada awal dekomposisi bahan organik tanah, dengan reduksi oksida-oksida besi dan mangan, nilai pH meningkat tajam sebagai hasil dari konsumsi proton yang jauh lebih besar daripada pelepasan proton. Tetapi pada tahap dekomposisi bahan organik yang aktif, pH dapat menurun karena produksi proton yang cepat. Selanjutnya pH tanah meningkat secara perlahan dan sampai pada tingkat tertentu stabil (Qixiao 10 dan Tianren, 1997). Reaksi tanah (pH) tanah sawah alkalin dikendalikan oleh kesetimbangan kimia dari sistem CaCO 3-CO 2 dalam tanah-tanah berkapur (Ponnamperuma, 1977; Qixiao dan Tianren, 1997), sistem Na2CO 3-CO2 untuk tanah sodik (Ponnamperuma, 1977). Biasanya nilai pH menurun karena adanya akumulasi CO 2 selama penggenangan. Penurunan pH tanah-tanah alkalin dan peningkatan pH tanah-tanah masam tersebut menguntungkan pertumbuhan tanaman. Akibat penggenangan, kekuatan ion (ionic strength) dalam larutan tanah meningkat, kemudian menurun. Dalam tanah-tanah masam atau agak masam, reduksi feri dan mungkin mangani yang tidak larut menjadi bentuk yang lebih larut menyebabkan peningkatan kekuatan ion. Dalam tanah netral sampai alkalin, Ca2+ dan Mg2+ juga menyebabkan peningkatan kekuatan ion. meningkatkan kelarutan Fe, Ca dan Mg. mengandung N-NO3- , Bahan organik Jika tanah yang awalnya banyak kekuatan ion dalam tanah dapat berkurang dengan penggenangan karena hilangnya NO3- akibat denitrifikasi (Mikkelsen, 1987). Proses reduksi tanah yang terjadi dalam tanah tergenang merupakan proses biokimia, dan jasad renik bertanggung jawab pada perubahan-perubahan (transformasi) yang terjadi di dalam tanah. Proses reduksi tidak terjadi pada tanah yang steril. Tanaman padi juga memengaruhi tingkat reduksi tanah karena adanya sekresi O2 dari akar-akar tanaman padi. 2.2. Pengaruh Penggenangan terhadap Tanaman Padi Padi merupakan tanaman yang unik karena dapat bertahan hidup dan bereproduksi di bawah kondisi lahan kering, tergenang dan air dalam. Walaupun medium berair (aquatik) cocok untuk pertumbuhan dan hasil tanaman padi, tetapi pertumbuhan akar memerlukan suplai O2 dan melepaskan CO2 selama respirasi. Hal ini dapat dilakukan oleh tanaman padi karena adanya sistem saluran pembawa udara (aerenchyma ) yang mampu mengalirkan O2 dari daun ke korteks akar, sehingga akar -akar tanaman padi dapat mengaerasi tanah tanpa mengambil O 2 dari tanah. 11 2.3. Transformasi Nitrogen dalam Tanah Tergenang Nitrogen (N) merupakan unsur hara pembatas pertumbuhan tanaman dalam tanah-tanah tergenang, baik tanah-tanah tergenang tersebut merupakan lahan basah alami ataupun pada lahan basah pertanian seperti tanah sawah (Gambrell dan Patrick, 1978). Transformasi N dalam tanah tergenang melibatkan berbagai proses mikrobiologi, dan beberapa proses tersebut menyebabkan hara N menjadi kurang tersedia bagi tanaman. Ion ammonium merupakan bentuk utama dari N yang dimineralisasi dalam sebagian besar tanah tergenang. Gambar 2 menggambarkan interaksi kompleks yang ada dalam tanah-tanah tergenang yang menyebabkan unsur hara N hilang dari tanah. Kehilangan N dapat terjadi dalam lapisan tanah yang teroksidasi dan tereduksi, dari air genangan, terbawa oleh aliran permukaan, pencucian, serapan N oleh tanaman dan karena mekanisme lainnya. Nitrogen mengalami beberapa transformasi fisikokimia dan biologi dalam tanah. Transformasi fisikokimia meliputi terperangkapnya (fiksasi) NH4+ dalam kisi-kisi minerali liat dan volatilisasi NH3. Transformasi N secara biologi meliputi mineralisasi-imobilisasi, fiksasi N2 atmosfer secara biologi, nitrifikasi- denitrifikasi, dan serapan tanaman. Sistem budidaya padi yang melibatkan penggenangan secara terus menerus dan berkala memengaruhi perilaku N tanah dan N yang diberikan ke dalam tanah. Kondisi khusus yang terjadi di bawah lingkungan tanah tergenang mempercepat proses amonifikasi dan menekan nitrifikasi bila tidak ada O2. Dengan penggenangan, N-NH4+ terakumulasi dalam tanah dan N -NO 3- hilang. Sumber-sumber pupuk N untuk tanaman padi dapat dibagi menjadi dua kelompok, yaitu sisa-sisa tanaman (pupuk organik) dan pupuk N mineral. Sebelum N yang terkandung dalam sisa-sisa tanaman menjadi tersedia bagi tanaman, sisa tanaman yang diberikan ke dalam tanah harus mengalami dekomposisi atau degradasi secara biologi lebih dulu. Urea mengalami hidrolisis enzimatik dan diubah menjadi N -NH 4+ (Kirk dan Olk, 2000): NH2CONH2 + 3H2O 2NH4+ + HCO3- + OH- 12 Kehilangan NH3 lewat daun Air hujan Pupuk N [CO(NH 2) 2, (NH4) 2SO4] N2 + O2 atm NH3 daun busuk alga AIR Lapisan N2 tanah oksida tif bakteri NH 4+ N-org NO 3- fungi aliran keluar TANAH residu tanam an Lapisan tanah reduktif NH4+ NO 3- N2 + N 2O bakteri N2 N-org Tapak bajak NH4+ NO 3- Lapisan tanah teroksidasi Fraksi Pencucian Gambar 2. Skema Transformasi Nitrogen dalam Ekosistem Tanah Sawah Tergenang (Sumber: Mikkelsen, 1987). 2.3.1. Pergerakan Nitrogen dalam Tanah Tergenang Pergerakan N dalam tanah berperan penting dalam menentukan bentuk kimia dan kete rsediaannya bagi pertumbuhan tanaman. Menurut Mikkelsen (1987) dua proses penting yang terlibat dalam pergerakan atau pengangkutan N adalah (1) pergerakan bahan-bahan terlarut dalam larutan tanah karena aliran 13 massa, dan (2) difusi molekul atau ion karena adanya gradien konsentrasi. Prosesproses ini memengaruhi difusi molekul dari bahan terlarut, seperti NH4+, NO2- , NO3-, urea dan gas-gas termasuk O 2, NH3 , N 2 dan N 2O (Rolston et al., 1990). Dalam tanah-tanah tergenang, pergerakan N terlarut dari lapis an tanah tereduksi (anaerob) ke lapisan permukaan yang aerob terjadi terutama melalui difusi dan dipengaruhi oleh gradien konsentrasi, sumber N, dan konsentrasi dalam lapisan anaerob (Reddy dan Patrick, 1984). Difusi ammonium dapat menyebabkan terjadinya pergerakan ammonium terlarut dari tanah ke air genangan, bahkan bila bahan pupuk dimasukkan ke dalam tanah (deep-placed). Nitrat yang ada dalam air genangan, dalam lapisan aerob, atau yang ada di lapisan oksidatif di sekitar akar padi segera berdifusi ke dalam lapisan anaerob yang terletak di bawahnya. Nitrogen-nitrat yang berdifusi ke dalam lapisan anaerob tampaknya hilang melalui denitrifikasi; sedangkan N-NH4+ yang berdifusi dari lapisan tanah aerob ke dalam air genangan rentan terhadap nitrifikasi dan volatilisasi ammonia (Savant dan De Datta, 1982). Pergerakan N-NH4+ dari tapak-tapak pertukaran ke dalam larutan dapat terjadi sebagai akibat dari adanya serapan tanaman, imobilisasi N yang membentuk jaringan tubuh jasad renik, nitrifikasi dan volatilisasi. 2.3.2. Mineralisasi dan Imobilisasi Nitrogen Ketersediaan N bagi tanaman sebagian besar dikendalikan oleh besarnya pengaruh dua proses di dalam tanah yang saling berlawanan, yaitu mineralisasi dan imobilisasi N. Mineralisasi N merupakan salah satu dari berbagai proses dalam siklus N di alam yang paling penting. Mineralisasi N adalah transformasi biologi dari N yang terikat secara organik menjadi N-mineral (N-NH4+ dan NNO3-) selama proses dekomposisi (Gambrell dan Patrick, 1978), dan dimulai dengan aminisasi dan amonifikasi, berturut-turut adalah konversi mikrobiologi dari N-organik menjadi R-NH2 dan menjadi N-NH4+ (Mikkelsen et al., 1995), dan selanjutnya menjadi N-NO3- melalui proses nitrifikasi. Tahap aminisasi dan amonifikasi berlangsung dengan bantuan jasad renik heterotrof, sedangkan nitrifikasi terjadi karena peranan bakteri ototrof. Pada kondisi tergenang, mineralisasi berhenti pada pembentukan N-NH4+ karena kondisi oksidatif yang diperlukan untuk nitrifikasi terhalang dengan adanya air yang tergenang. 14 Imobilisasi N merupakan proses kebalikan dari mineralisasi N, dan didefinisikan sebagai konversi N-mineral menjadi bentuk N-organik dalam jaringan tubuh jasad renik (Soil Science Society of America, 1987). Imobilisasi (sintesis) dan mineralisasi atau pelepasan N dari senyawa organik dalam tanah terjadi karena aktivitas jasad renik yang mengarah pada pertukaran secara terus -menerus antara bentuk-bentuk N-organik dan mineral (Mikkelsen, 1987). Mineralisasi dan imobilisasi merupakan proses yang berlawanan yang terjadi secara serentak dan terus-menerus dan sangat memengaruhi ketersediaan N bagi tanaman dan konversi N dalam tanah dalam bentuk organik atau terfiksasi. Konsentrasi NNH4+ dapat meningkat atau menurun tergantung pada dominasi relatif kedua proses tersebut. Proses amonifikasi, yang melibatkan hidrolisis enzimatik dan deaminasi N organik tanah menjadi ammonium, terjadi secara bersamaan dengan proses assimilatory dari imobilisasi N oleh mikrorganisme tanah. Pembebasan N -NH4+ ke dalam tanah tergenang tergantung pada kebutuhan N populasi jasad renik tanah, nisbah C/N dari sisa-sisa tanaman yang terdekomposisi (Mikkelsen, 1987), komposisi kimia bahan organik dan beberapa faktor lingkungan. Komponen organik dalam sisa-sisa tanaman umumnya dibagi menjadi enam kategori, yaitu selulosa, hemiselulosa, lignin, fraksi larut air (meliputi gula sederhana, asam amino, dan asam-asam alifatik), komponen larut dalam eter dan alkohol, dan protein (Nagarajah, 1997). Lignin merupakan komponen organik dari sisa-sisa tanaman yang paling tahan terhadap pelapukan. Secara agronomi, N yang dimineralisasi sangat penting dan merupakan 50-80% dari N-total yang diasimilasi oleh tanaman padi (Mikkelsen et al, 1995). Untuk mencapai penggunaan N tanah yang efisien da lam bentuk N terfiksasi secara biologi, sisa-sisa tanaman, dan pupuk, maka perlu dipertimbangkan aspek laju-waktu proses mineralisasi relatif terhadap kebutuhan N tanaman padi. Dalam jangka pendek, suplai N untuk tanaman padi diatur oleh laju mineralisasi N-organik menjadi N-NH4+. Senyawa N-organik, terutama protein dan turunannya mengalami pelapukan secara anaerob menghasilkan bentuk-bentuk yang lebih sederhana, seperti asam amino (Nagarajah, 1997). Nitrogen-asam amino lebih rentan terhadap mineralisasi daripada fraksi N tanah lainnya (Mikkelsen, 1987). Deaminasi senyawa N yang lebih sederhana 15 menghasilkan pelepasan N-NH 4+ yang merupakan bentuk akhir dari senyawa N dan bersifat stabil dalam lapisan tereduksi. Dalam tanah tergenang, laju mineralisasi N sisa-sisa tanaman yang diberikan ke dalam tanah ditentukan terutama oleh nisbah L/N-nya (Becker et al., 1994). Suplai N melalui mineralisasi bahan organik tanah telah diteliti oleh Stanford dan Smith (1972). Mereka melakukan percobaan di laboratorium untuk menentukan besarnya N yang dimineralisasi atau dilepaskan dari bahan organik tanah. Smith et al. (1977) juga telah melakukan percobaan di lapang dengan mengukur jumlah N mineral dalam pot-pot yang berisi tanah tanpa tanaman yang dibenamkan ke dalam tanah. Hasil yang diperoleh menunjukkan bahwa potensi mineralisasi N tanah di lapang sesuai dengan hasil pengukuran dari percobaan laboratorium. Percobaan ini menunjukkan bahwa sebagian dari N dalam tanah berasal dari proses mineralisasi senyawa N yang mudah dimineralisasi, dan sisanya diasumsikan sebagai bentuk senyawa N yang tidak tersedia dan tetap berada dalam bahan organik tanah yang relatif stabil. 2.3.3. Nitrogen terlarut dan dapat dipertukarkan Nitrogen terlarut dan dapat dipertukarkan (N-tersedia) merupakan fraksi N yang sangat penting sebagai nutrisi tanaman. Sumber utama dari N-tersedia berasal dari pupuk dan N hasil mineralisasi. Nitrogen-NH4+ yang dibebaskan selama pelapukan sisa-sisa tanaman, dengan cepat dijerap pada kompleks pertukaran kation dan berada dalam keseimbangan dengan N-NH4+ dalam larutan tanah. Proporsi relatif dari kedua bentuk N tersebut sebagian besar diatur oleh kapasitas pertukaran kation (KTK) tanah (Mikkelsen, 1987; Ando et al., 1996) dan sifat dari kompleks pertukaran kation (Nagarajah, 1997). Beberapa dari NNH4+ juga berada dalam bentuk tidak dapat dipertukarkan (terperangkap dalam kisi-kisi mineral liat). Proses ini terjadi dalam tanah yang banyak mengandung mineral liat 2:1 seperti vermikulit dan illit. Setelah beberapa hari penggenangan, konsentrasi N-NH4+ dapat dipertukarkan dalam tanah dapat meningkat karena mineralisasi bahan organik tanah dan pelepasan N-NH4+ yang terfiksasi dalam kisi mineral liat. Menurut Mikkelsen (1987) pemberian jerami padi akan menurunkan tingkat N tersedia karena imobilisasi, segera setelah diberikan ke dalam tanah. Tetapi setelah 16 periode waktu tertentu N yang terimobilisasi tersebut akan dilepaskan kembali melalui mineralisasi sehingga menjadi tersedia bagi tanaman padi. 2.3.4. Penambatan (Fiksas i) N2 Secara Biologi Dalam sistem pertanian subsisten (di banyak bagian dari Asia tropis), usaha tani padi sawah telah dilakukan secara terus menerus selama berabad-abad tanpa pemberian pupuk N tanpa memperlihatkan penurunan konsentrasi N tanah yang nyata. Di antara proses-proses yang turut menyumbang suplai N di lahan sawah, penambatan N2 secara biologi dianggap sebagai faktor penting dalam mempertahankan kesuburan N tanah. Data neraca N dari beberapa percobaan jangka panjang di lapang menunjukkan bahwa sumbangan penambatan N secara biologi ke dalam tanah selama musim pertanaman padi sawah berkisar dari 19 sampai 38 kg N ha -1 di Jepang, dan 30 sampai 52 kg N ha -1 di Philipina. Sumbangan N ini terutama berasal dari jasad renik asli (indigenous) yang bersifat asosiatif dan penambat N2 yang hidup bebas, yang meliputi bakteri heterotrof dan fototrof serta cyanobakteri (alga hijau-biru) yang ada dalam sistem tanahtanaman-air genangan lahan sawah (Kundu dan Ladha, 1995). Sumber unsur hara N terbesar adalah N2 udara yang merupakan 80% dari atmosfer bumi. Akan tetapi sebagian besar organisme hanya dapat menggunakan N yang bersenyawa dengan atom-atom lainnya untuk membentuk suatu ion seperti NH4+ atau NO3-. Bentuk N sebagai N2 tidak dapat digunakan secara langsung oleh sebagian besar tanaman karena adanya ikatan rangkap tiga yang membuatnya menjadi molekul yang bersifat inert (Deacon, 2003). Gas N2 ini sangat stabil dan tersedia melimpah bagi organisme yang mampu memanfaatkannya. Penambatan atau fiksasi N2 secara biologi dapat mengkonversikan gas N2 menjadi N organik melalui aktivitas organisme tertentu, baik aerob maupun anaerob, yang memiliki enzim nitrogenase. Dengan demikian penambatan N2 secara biologi menjadi sumber N utama bagi lahan sawah dalam system pertanian padi sawah yang bersifat tradisional dan subsisten. Nitrogenase merupakan enzim yang sangat sensitif terhadap O2 (Bergesen, 1980). Ekosistem sawah sangat cocok untuk proses fiksasi N2 karena tegangan O2 dalam ekosistem sawah rendah. Fiksasi N2 dapat terjadi dalam air genangan, lapisan tanah aerob, lapisan tanah anaerob, rhizosfer tanaman yang oksidatif, pada 17 permukaan daun dan batang tanaman (Reddy dan Graetz, 1988). Fiksasi N2 pada kondisi sawah dapat dilakukan oleh bakteri non-simbiotik (alga hijau-biru), dan pada kondisi lahan kering dilakukan oleh bakteri simbiotik dari genus Rhizobium, atau oleh aktinomisetes. Roger dan Watanabe (1986) mengklasifikasikan organisme penambat N2 secara ekologi menjadi : (1) tiga kelompok jasad renik autotrof yang terdiri dari bakteri fotosintetik, alga hijau biru yang hidup bebas (non-simbiotik), dan Anabaena azollae sp. yang berasosiasi dengan tanaman paku air Azolla, dan (2) tiga kelompok jasad renik heterotrof yang terdiri dari bakteri penambat N2 dalam tanah (aerob, anaerob fakultatif dan anaerob obligat) yang hidup bebas, bakteri penambat N2 yang berasosiasi dengan akar-akar tanaman padi, dan organisme yang bersimbiose dengan legum (Rhizobium). Sampai saat ini hanya tanaman legum yang mampu menambat N 2 dan pupuk hijau Azolla yang digunakan sebagai sumber N bagi tanaman padi melalui penambatan N2 secara biologi (Mikkelsen et al., 1995). Menurut Kundu dan Ladha (1995) tanah yang sangat reduktif (yang tercipta karena penggenangan secara terus -menerus selama masa pertumbuhan tanaman padi) dan pelumpuran yang intens memberikan pengaruh yang kurang baik bagi bakteri penambat N asli (indigenous) dalam tanah sawah. Oleh karena itu, sistem yang dapat mendorong atau meningkatkan penambatan N2 akan sangat membantu mempertahankan kesuburan N pada tanah-tanah sawah (Ladha dan Kundu, 1997). Selain pemberian pupuk hijau dari tanaman yang mampu menambat N2 seperti Sesbania dan Azolla, pemberian sisa tanaman dengan nisbah C/N yang besar (seperti halnya jerami padi) juga dapat meningkatkan penambatan N2. Jerami padi merupakan sumber energi yang baik bagi bakteri heterotrof, dan pengembalian jerami padi ke dalam lahan sawah secara nyata dapat meningkatkan fiksasi N2 oleh bakteri heterotrof maupun fototrof (Matsuguchi, 1979; Ventura et al., 1986; Adachi et al., 1997). Roger dan Ladha (1990) juga menyatakan bahwa pemberian jerami ke dalam tanah dapat memberikan N sebesar 2-4 kg N untuk setiap ton jerami. Hal ini menurut Ponnamperuma (1984) karena pemberian jerami padi dan N mineral meningkatkan populasi bakteri aerob penambat N2. Selain itu Greenland (1997) menyatakan bahwa aktivitas bakteri penambat N2 dan alga hijau-biru ditentukan oleh kondisi lingkungan tempat tumbuhnya. Aktivitas 18 mereka sebagian besar tergantung pada ketersediaan fosfor (P) dalam tanah sawah tetapi konsentrasi N yang tinggi dalam tanah sawah cenderung menghambat fiksasi N2. 2.3.5. Volatilisasi Amonia Volatilisasi ammonia (NH 3) merupakan mekanisme kehilangan N yang penting dalam sistem pertanaman yang dipupuk. Menurut Mikkelsen (1987) faktor-faktor dominan yang memengaruhi volatilisasi NH3 adalah pH tanah dan pe (- log konsentrasi elektron), tekanan parsial CO2 (pCO2) dan kimia karbonat, sifat pertukaran kation dan aktivitas jasad renik. Selain itu, kecepatan angin, konsentrasi NH3 terlarut dan tekanan parsial NH3 dalam air dan udara, suhu udara dan radiasi langsung juga memengaruhi volatilisasi NH3. Menurut Zhenghu dan Honglang (2000) laju volatilisasi ammonia berkorelasi positif dengan pH tanah, kandungan CaCO3 , dan garam total, tetapi berorelasi negatif dengan kandungan bahan organik, KTK, dan kandungan liat. Dari ketiga faktor yang berkorelasi negatif, KTK merupakan faktor yang korelasinya sangat tinggi dengan volatilisasi ammonia, sedangkan faktor pH tanah merupakan faktor yang dominan di antara ketiga faktor yang berkorelasi positif. Menurut Zhenghu dan Honglang (2000) bahan organik berpengaruh secara tidak langsung pada penurunan volatilisasi ammonia melalui pengaruhnya terhadap penurunan pH tanah dan meningkatnya KTK tanah karena adanya pembentukan berbagai asam-asam organik dan humus selama proses dekomposisi bahan organik. Amonia yang dihasilkan dalam sistem karbonat aquatik melibatkan reaksi berikut (Mikkelsen et al., 1978): NH4+ + OH- (NH3)aq + H 2O NH4+ + HCO 3- (NH3) aq + H2O + CO2 2NH 4+ + CO32- 2(NH3) aq + H2O + CO2 Volatilisasi ammonia terjadi bila pH air genangan meningkat di atas pH 7,5 (Greenland, 1997). Hilangnya gas CO2 yang meningkat karena meningkatnya suhu air genangan pada siang hari dapat menyebabkan pH meningkat. Akan tetapi penyebab utama peningkatan pH dalam air genangan tanah sawah adalah pertumbuhan alga atau adanya proses biologi yang berlawanan yaitu fotosintesis dan respirasi (Greenland, 1997). Respirasi dan fotosintesis menyebabkan 19 perubahan tekanan parsial CO2 dalam air genangan, dan sistem karbonat ini sangat menentukan pH air. Nilai pH air genangan ditentukan oleh konsentrasi CO2 dalam air (Manahan, 1994). Sistem karbonat atau sistem CO2 - HCO3- - CO32- dalam air digambarkan oleh reaksi berikut dan konstanta kesetimbangannya (Manahan, 1994): CO2 + H2O HCO3- + H+ K1 = 4,45 x 10-7 (pK 1 = 6,35) HCO3- CO32- + H+ K2 = 4,69 x 10-11 (pK 2 = 10,33) Dari persamaan di atas, bila fotosintesis aktif maka akan terjadi penurunan konsentrasi CO2 dalam sistem dan hal ini menyebabkan persentase fraksi mol asam karbonat meningkat, akibatnya pH sistem meningkat. Tingkat perubahan pH yang disebabkan oleh jasad renik ditentukan oleh jumlah, jenis dan aktivitas organisme yang ada. Korelasi antara pH air dan sistem asam karbonat bersifat kompleks dan tidak dapat digambarkan secara lengkap tanpa mempertimbangkan sejumlah variabel. Dengan memperhatikan reaksi biokimia dari jasad renik aquatik, reaksi yang paling sederhana adalah : fotosintesis n CO2 + n H 2O (CH2O)n + n O2 respirasi Setiap hari terjadi fluktuasi pH dalam air genangan dari 7,5 – 9,5 dan nilai pH maksimum terjadi kira-kira pada pukul 14.00 dan menurun sepanjang sore hari. Pola perubahan pH ini sesuai dengan siklus fotosintesis dan respirasi dari jasad renik aquatik (Mikkelsen, 1987). Amonia dan bentuk ionnya (NH4+) merupakan hasil dekomposisi bahan organik tanah dan sisa-sisa tanaman yang terjadi dalam perairan alami. Penggunaan pupuk N pada lahan sawah juga menyebabkan konsentrasi garam NNH4+ terlarut meningkat. langsung, atau seperti Pupuk ammonium dalam air dapat berdisosiasi urea terdekomposisi melalui hidrolisis katalitik menghasilkan ion-ion NH4+. Ion-ion NH4+ , yang memiliki ikatan sangat lemah dengan molekul air, dominan dalam air dengan pH di atas 7,2. Dengan meningkatnya konsentrasi ion hidroksil (OH -) dalam air, maka terjadi peningkatan perubahan ion NH4+ menjadi NH3 yang dapat menghilang dari air dalam bentuk gas (Mikkelsen et al., 1978). Ventura dan Yoshida (1977) mengukur volatilisasi 20 NH3 dari sumber N yang berbeda pada tanah liat tergenang, dan menyatakan bahwa kehilangan NH3 pada dasarnya terjadi selama sembilan hari pertama setelah pemberian pupuk N. Kehilangan tersebut kecil bila pH tanah di bawah 7,4. Percobaan lapang yang mereka lakukan menunjukkan bahwa kehilangan N setelah pemberian 100 kg N/ha dengan cara disebar adalah sebesar 3,8% untuk ammonium sulfat (ZA) dan 8,2% dengan pemberian urea. Menurut Vlek dan Stumpe (1978) volatilisasi NH3 dari tanah yang dipupuk urea lebih besar daripada tanah yang dipupuk ammonium sulfat. Hal ini terjadi karena hidrolisis urea dalam tanah mendorong terciptanya lingkungan yang ideal untuk volatilisasi, yaitu alkalinitas dan pH yang tinggi. Volatilisasi ammonia berkurang 50% bila pupuk dimasukkan ke dalam tanah. 2.3.6. Nitrifikasi dan Denitrifikasi Oksidasi biologi dari N-NH4+ menjadi N-NO3- (nitrifikasi) menghasilkan konversi atau perubahan kation NH4+ yang relatif tidak mobil menjadi bentuk anion (NO3-) yang lebih mobil, yang pada gilirannya anion ini rentan terhadap denitrifikasi. Menurut Kakuda et al. (1999), denitrifikasi merupakan proses utama kehilangan N dalam tanah sawah. Tanah tergenang merupakan lingkungan yang ideal untuk denitrifikasi karena lingkungan tanah tergenang memiliki suatu lapisan permukaan teroksidasi yang tipis yang di bawahnya adalah lapisan tereduksi yang tebal. Lapisan tanah teroksidasi mendukung proses nitrifikasi dan lapisan tanah tereduksi merupakan lapisan tanah yang kekurangan oksigen dan menyediakan bahan organik yang mudah didekomposisi untuk mendukung proses reduksi bentuk-bentuk N teroksidasi (denitrifikasi). Adanya lapisan zona aerob dan anaerob dalam tanah tergenang, begitu juga dalam rhizosfer (karena adanya bagian tanaman padi yang mengangkut O 2 ke rhizosfer), memudahkan terjadinya reaksi nitrifikasi-denitrifikasi. Sebagaimana diketahui bahwa akar-akar tanaman tersebar dalam tanah permukaan dan tanah di bawah perm ukaan (subsurface). Reaksi ini tampaknya terjadi secara serentak. Denitrifikasi dalam rhizosfer dipengaruhi oleh metabolisme tanaman dan lingkungan tanah (Kakuda et al., 1999). Hal ini didukung oleh adanya suplai C dari akar (Mahmood et al., 1997). Eksudat senyawa organik oleh akar-akar yang hidup merangsang respirasi bakteri (Klemedtsson et al., 1987). Denitrifikasi tergantung pada senyawa organik yang 21 berasal dari akar bila suplai C dari tanah terbatas (Prade dan Trolldenier, 1990). Namun demikian, pertumbuhan tanaman padi juga berpengaruh terhadap kehilangan N melalui denitrifikasi, yaitu bahwa serapan N oleh tanaman dapat mengurangi jumlah N yang hilang melalui denitrifikasi. Ion nitrat (NO 3- ) merupakan ion bermuatan negatif sehingga tidak dapat dijerap oleh partikel tanah yang bermuatan negatif dan selanjutnya menjadi sangat mobil dalam larutan. Bila ion nitrat tidak segera diasimilasi oleh tanaman atau jasad renik (assimilatory nitrate reduction), atau hilang melalui pencucian, maka ion nitrat berpotensi mengalami dissimilatory nitrogenous oxide reduction, suatu istilah yang mengacu pada beberapa jalur mekanisme reduksi nitrat (Wiebe et al., 1981), yang paling umum adalah reduksi nitrat menjadi ammonia dan denitrifikasi. Jasad renik yang bertanggung jawab pada proses denitrifikasi dalam tanah adalah bakteri heterotrof. Jasad renik ini membutuhkan oksida-oksida N sebagai penerima elektron terakhir dan C-organik sebagai donor elektron, serta kondisi anaerob. Denitrifikasi adalah proses reduksi bentuk-bentuk N mineral teroksidasi menjadi gas nitrogen, terutama N 2O dan N2, yang dilakukan oleh jasad renik dalam kondisi anaerob, dimana nitrat (NO3- ) bertindak sebagai penerima elektron terakhir atau NO 3- direduksi. Proses ini menyebabkan N-NO3- hilang karena dikonversi menjadi gas N2O dan N2. Jalur pembentukan gas-gas tersebut digambarkan sebagai berikut (Firestone, 1982; Ostrom et al., 2000) : Denitrifikasi : 2 NO3- 2 NO2- 2 NO N2O N2 Nitrifikasi : O2 2 NH4+ 2 NH2OH 2 (NOH) jalur 1 N2O 2 NO 2 NO2 jalur 2 NO2 Nitrifikasi – Denitrifikasi : O2 2 NH4+ 2 NH 2OH 2 (NOH) 2 NO N2O jalur 3 2 NO 2 22 Sifat anaerob tanah tergenang menyebabkan ketidakstabilan NO 3- , NO2, dan N2O yang berperan sebagai penerima elektron terakhir dalam respirasi anaerob berbagai jasad renik heterotrof. Hal ini menyebabkan hilangnya N dari ekosistem sawah dalam bentuk N2 dan N2O. Di-nitrogen (N2) merupakan produk denitrifikasi yang paling akhir. Peningkatan konsentrasi N2O sebagai gas rumah kaca dalam atmosfer dapat menyebabkan terjadinya kerusakan lapisan ozon stratosfer sehingga berpengaruh terhadap perubahan iklim global (Intergovermental Panel on Climate Change/IPCC, 1994). Terdapat dua mekanisme pembentukan gas N2O selama nitrifikasi, yaitu sebagai hasil reduksi NO 2- pada kondisi anaerob oleh bakteri nitrifikasi tertentu dan hasil dari berbagai reaksi antara dalam oksidasi NH4+ (Ritchie dan Nicholas, 1972). Proses nitrifikasi dan denitrifikasi terjadi melalui suatu “pool” nitrat (Gambar 3), dimana nitrat yang terbentuk dalam zona aerob berdifusi ke zona anaerob dan direduksi menjadi N2O dan N2. Hasil penelitian Russow et al. (2000) dengan menggunakan pelacak 15 N-NO3- menunjukkan bahwa NO3- merupakan pool utama pembentukan N2O pada kondisi jenuh air melalui proses denitrifikasi. Penelitian Russow et al. (1996) dengan metode aliran gas 15 N juga menunjukkan bahwa sekitar 60% dari nitrat tanah direduksi menjadi N2. Sebaliknya nitrit berada dalam dua pool yang terpisah karena kecepatan transformasinya lebih tinggi daripada kecepatan difusinya (Russow et al., 2000). Kehilangan N dari pupuk N yang diberikan ke dalam tanah melalui denitrifikasi bervariasi dari 0 sampai 70% (Mikkelsen, 1987). Dengan demikian upaya mengendalikan kehilangan N karena denitrifikasi dapat memperbaiki efisiensi penggunaan pupuk N oleh tanaman padi sawah. Beberapa praktek pengelolaan tanah dan tanaman untuk mengendalikan kehilangan N karena denitrifikasi telah banyak dilakukan misalnya dengan penempatan pupuk N pada lapisan tanah tereduksi, penggunaan bahan penghambat (inhibitor) nitrifikasi dan urease. 23 NO N 2O N2 atmosfer tanah Zona Aerob NH4+ BOT Zona Anaerob NH 4+ N2 N 2O JALUR NITRIFIKASI JALUR DENITRIFIKASI NO 2- NO2pool nitrit BOT = Bahan organik tanah NO 3pool nitrat Gambar 3. Model Pasangan Proses Nitrifikasi – Denitrifikasi yang Menggambarkan Mekanisme Kedua Proses Tersebut dengan Penekanan Khusus pada Pembentukan N2O dan N 2 (Sumber : Russow et al., 2000). 2.3.7. Kehilangan Nitrogen dari Tanaman Kehilangan N juga dapat terjadi melalui bagian atas tanaman setahun dan tahunan, termas uk tanaman padi. Sejumlah penelitian menunjukkan bahwa banyak N yang hilang dari permukaan daun berkaitan dengan penguapan air transpirasi (Stutte dan Weiland, 1978; Weiland dan Stutte, 1979; da Silva dan Stutte, 1981). da Silva dan Stutte (1981) menyata kan bahwa laju kehilangan N berkurang selama periode antara pembentukan anakan (tillering) dan 1-3 minggu setelah awal pembentukan malai (panicle initiation), selanjutnya meningkat sampai pada stadia pengisian bulir (heading ). Umumnya daun yang berkembang penuh (matured leaf) menunjukkan laju kehilangan N yang lebih tinggi daripada daun yang lebih tua. Namun konsentrasi N dalam larutan hara tidak memengaruhi laju kehilangan N per satuan luas daun dari daun yang berkembang penuh. Sebaliknya pada daun-daun yang lebih tua dari tanaman yang disuplai dengan takaran N tinggi (80 ppm), laju kehilangan N lebih tinggi daripada yang 24 ditumbuhkan pada medium dengan takaran N rendah (20 ppm). Glutamin sintetase dianggap sebagai jalur mekanisme utama dari asimilasi ammonia dan peningkatan penambatan kembali NH3 yang dilepaskan selama fotorespirasi. Nitrogen yang dilepaskan melalui daun-daun tanaman selama fotorespirasi berupa NH3 dan amin. Dengan mengkuantifikasikan pengaruh fisiologi terhadap kehilangan N dapat membantu menentukan waktu aplikasi dan jumlah pupuk N yang diperlukan untuk produksi padi yang optimum. 2.4. Siklus Pertumbuhan Tanaman Padi Sawah Tanaman padi biasanya memerlukan waktu 3-6 bulan dari perkecambahan sampai stadia pematangan, tergantung pada varietas dan lingkungan dimana tanaman padi tersebut ditumbuhkan. Menurut Yoshida (1981) secara agronomi pertumbuhan tanaman padi dibagi menjadi tiga stadia, yaitu vegetatif, reproduktif dan proses pematangan (ripening ). Stadia vegetatif menunjuk pada suatu masa dari perkecambahan sampai inisiasi pembentukan malai (initiation of panicle primordial), yang sebagian besar dicirikan oleh pembentukan anakan (tiller formation ) (Mikkelsen et al., 1995); stadia reproduktif adalah suatu masa dari inisiasi pembentukan malai sampai pembungaan, pada stadia ini jumlah bakal bulir padi (spikelet) dan potensi ukurannya sebagian besar ditentukan oleh nutrisi tanaman dan faktor -faktor lingkungan (Mikkelsen et al., 1995); dan proses pematangan (ripening ) adalah dari pembungaan sampai matang (grain maturity) (Gambar 4). Persentase bulir yang matang dan ukuran bulir ditentukan selama stadia perkembangan ini. Varietas padi yang berumur 120 hari bila ditanam di lingkungan tropika, menghabiskan waktu 60 hari untuk masa vegetatif, 30 hari untuk masa reproduktif, dan 30 hari untuk masa pemasakan. Stadia vegetatif dicirikan dengan pembentukan anakan yang aktif, peningkatan tinggi tanaman secara perlahan, dan munculnya daun pada interval yang teratur. Semua hal tersebut berkaitan dengan peningkatan luas daun yang menerima cahaya. Stadia reproduktif dicirikan dengan pemanjangan tangkai (culm elongation), yang meningkatkan tinggi tanaman; penurunan jumlah anakan, munculnya daun bendera (daun terakhir), pembungaan, pembentukan bulir padi (booting), dan pengisian bulir padi. 25 Jumlah Pertumbuhan Jumlah Anakan Tinggi Tanaman Anakan yang tidak Efektif Jumlah malai ( panicle) Bobot bulir 0 20 60 90 120 Vegetatif Gambar 4. Reproduktif Masak (panen) Pengisian bulir padi (heading) Pembentukan bulir padi (booting) Awal munculnya malai Pembentukan anakan Tanam Perkecambahan Hari setelah Perkecambahan Proses pematangan Siklus Pertumbuhan Varietas Tanaman Padi Berumur 120 Hari (Sumber: Yoshida, 1981) 2.5. Peranan dan Fungsi Nitrogen dalam Tanaman Padi Nitrogen merupakan komponen ya ng penting dari asam amino, asam nukleat, nukleotida, dan khlorofil. Nitrogen yang diserap oleh tanaman akan diasimilasi menjadi asam amino, yang berikutnya akan membentuk protein dan asam nukleat. Selain itu, N juga menjadi bagian integral dari khlorofil yang merupakan komponen utama tanaman yang menyerap cahaya yang dibutuhkan dalam proses fotosintesis. Struktur dasar dari khlorofil adalah cincin porfirin, yang terdiri dari empat cincin pyrole, masing-masing mengandung satu N dan empat atom C. Atom Mg diikat pada pusat cincin porfirin (Havlin et al., 1999). 26 Suplai N yang cukup berkaitan dengan aktivitas fotosintetik yang tinggi, pertumbuhan vegetatif yang pesat, dan warna hijau gelap. Kelebihan N berkaitan dengan unsur hara lainnya seperti P, K, dan S, dapat menunda kematangan tanaman. Suplai N memengaruhi penggunaan karbohidrat dalam tanaman. Bila suplai N rendah, maka karbohidrat akan dideposisikan dalam sel-sel vegetatif yang menyebabkan sel-sel vegetatif tersebut menjadi menebal. Tetapi bila karbohidrat yang dideposisikan dalam bagian vegetatif lebih sedikit, akan lebih banyak protoplasma yang terbentuk dan karena protoplasma ini sangat terhidrasi maka akan dihasilkan tanaman yang lebih sukulen. Bila suplai N cukup dan kondisi lingkungan sesuai untuk pertumbuhan, maka lebih banyak protein yang terbentuk (Havlin et al., 1999). Menurut Dobermann dan Fairhust (2000) diperlukan suplai N yang cukup selama masa pemasakan untuk menunda penuaan daun-daun, mempertahankan fotosintesis selama pengisian bulir, dan meningkatkan kandungan protein dalam gabah. Selanjutnya mereka menyatakan bahwa N -NO3- dan N -NH4+ adalah sumber utama dari serapan N mineral. Lebih lanjut menurut mereka, sebagian besar N-NH4+ yang diserap dikandung dalam senyawa organik dalam akar, sedangkan N-NO3bersifat lebih mobil dalam xylem dan juga disimpan dalam vakuola dari bagian tanaman yang berbeda. Untuk memenuhi fungsi esensial sebagai hara tanaman, N-NO3- harus direduksi menjadi ammonia melalui aksi reduktase nitrat dan nitrit. Nitrogen diperlukan sepanjang masa pertumbuhan, tetapi kebutuhan N yang terbesar adalah antara awal sampai pertengahan pembentukan anakan dan stadia pembentukan malai. Nitrogen berfungsi dalam mendorong pertumbuhan yang cepat (meningkatkan tinggi tanaman dan jumlah anakan) dan meningkatkan ukuran daun, jumlah bulir per malai, persentase bulir yang berisi pada masingmasing malai, dan kandungan protein gabah. Dengan demikian N memengaruhi semua parameter yang berhubungan dengan hasil. Konsentrasi N daun sa ngat erat kaitannya dengan laju fotosintesis daun dan produksi biomassa tanaman. Nitrogen juga mendorong kebutuhan akan unsur hara makro lainnya seperti P dan K (Dobermann dan Fairhust, 2000).