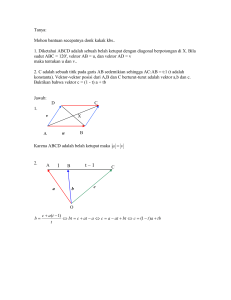
KLONING GEN HUMAN INTERFERON ALPHA 2a PADA VEKTOR
advertisement

UNIVERSITAS INDONESIA KLONING GEN HUMAN INTERFERON ALPHA 2a PADA VEKTOR pET-32b(+) DAN EKSPRESI PADA Escherichia coli TESIS ARIZAH KUSUMAWATI 1006787092 FAKULTAS FARMASI PROGRAM S2 ILMU KEFARMASIAN DEPOK JANUARI 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 UNIVERSITAS INDONESIA KLONING GEN HUMAN INTERFERON ALPHA 2a PADA VEKTOR pET-32b(+) DAN EKSPRESI PADA Escherichia coli TESIS Diajukan sebagai salah satu syarat untuk memperoleh gelar Magister Farmasi ARIZAH KUSUMAWATI 1006787092 FAKULTAS FARMASI PROGRAM STUDI S2 ILMU KEFARMASIAN DEPOK JANUARI 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 KATA PENGANTAR Puji syukur saya panjatkan kepada Allah SWT, karena atas berkat dan rahmatNya, saya dapat menyelesaikan tesis ini. Penulisan tesis ini dilakukan dalam rangka memenuhi salah satu syarat untuk mencapai gelar Magister Farmasi pada Fakultas Farmasi Universitas Indonesia. Saya menyadari banyaknya bantuan dan bimbingan dari berbagai pihak dari masa perkuliahan sampai pada penyusunan tesis ini, sehingga tesis ini dapat terselesaikan. Oleh karena itu, perkenankanlah saya dengan setulus hati mengucapkan rasa terima kasih yang sebesar-besarnya kepada: (1) Prof. Dr. Maksum Radji, M.Biomed., Apt. dan Dr. Adi Santoso, M.Sc., selaku dosen pembimbing yang telah menyediakan waktu, tenaga, dan pikiran untuk mengarahkan saya dalam penelitian dan penyusunan tesis ini; (2) Prof. Dr. Yahdiana Harahap, MS., Apt., selaku Dekan Fakultas Farmasi Universitas Indonesia; (3) Prof. Dr. Effionora Anwar, MS., Apt., selaku Ketua Program Studi S2 Ilmu Kefarmasian Fakultas Farmasi Universitas Indonesia yang telah memberikan dorongan untuk menyelesaikan program S2 Ilmu Kefarmasian; (4) Dr. Amarila Malik, M.Si., Apt., Dr. Herman Suryadi, MS., Apt., dan Dr. Mahdi Jufri, M.Si., Apt. selaku Dewan penguji yang telah banyak memberikan penilaian maupun saran-saran untuk perbaikan dan penyempurnaan tesis ini; (5) Pusat Penelitian Bioteknologi Lembaga Ilmu Pengetahuan Indonesia yang telah memberikan kesempatan kepada saya untuk menjadi peserta tugas belajar program pasca sarjana dan memfasilitasi saya demi kelancaran penelitian tesis ini; (6) Kementerian Riset dan Teknologi yang telah memberikan bantuan dana pendidikan kepada saya untuk dapat mengikuti program tugas belajar S2 Ilmu Kefarmasian di Universitas Indonesia; (7) Ayah, Ibu, Kakak, Mertua dan Suami yang senantiasa memberikan doa, semangat dan kasih sayang kepada saya; Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 (8) Teman-teman di Laboratorium Terapetik Protein dan Vaksin atas bantuan yang telah diberikan selama melaksanakan penelitian di Pusat Penelitian Bioteknologi LIPI; (9) Semua pihak yang telah memberikan bantuan hingga tesis ini dapat diselesaikan. Akhir kata, saya berharap Allah SWT berkenan membalas segala kebaikan semua pihak yang telah membantu. Semoga tesis ini membawa manfaat bagi pengembangan ilmu. Depok, 2 Januari 2013 Penulis Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 ABSTRAK Nama Program Studi Judul : Arizah Kusumawati : S2 Ilmu Kefarmasian : Kloning Gen Human Interferon Alpha 2a pada Vektor pET-32b(+) dan Ekspresi pada Escherichia coli Interferon (IFN) merupakan sitokin yang diproduksi oleh berbagai tipe sel sebagai respon rangsangan terhadap stimulasi virus, bakteri, parasit, sel tumor, atau antigen lain. Interferon α termasuk kelompok IFN tipe I yang mempunyai berbagai efek biologis yang meliputi antiviral, antitumor dan juga sebagai immunoterapetik. Penelitian ini bertujuan untuk mensintesis protein rekombinan human IFN α2a melalui sistem ekspresi pada bakteria E. coli BL21(DE3). Pada gen human ifn α2a dilakukan penambahan situs pemotongan enzim restriksi Nco I dan Xho I menggunakan metode PCR, kemudian dilanjutkan dengan proses ligasi ke vektor pET-32b(+) dan selanjutnya ditransformasikan pada E. coli DH5α. Hasil sekuensing menunjukkan bahwa vektor rekombinan (pET-32b(+)-IFN α2a) memiliki urutan nukleotida yang benar. Vektor rekombinan ini selanjutnya ditransformasikan ke dalam E.coli BL21(DE3). Klon transforman yang diperoleh dikultur dan diinduksi dengan penambahan IPTG 1 mM sehingga mengekspresikan protein rekombinan human IFN α2a. Dari hasil isolasi, diperoleh protein rekombinan human IFN α2a dalam bentuk protein terfusi sehingga mempermudah proses deteksi dan purifikasi. Protein dikarakterisasi melalui metode SDS PAGE dilanjutkan dengan Western blot dan pewarnaan CBB. Pita protein rekombinan human IFNα2a yang diperoleh berukuran 36 kDa. Hasil maksimal ditunjukkan ekspresi pada suhu 37⁰C dengan waktu inkubasi 5 jam setelah induksi. Kata Kunci xiv+74 halaman Daftar Pustaka : Interferon, vektor pET-32b(+), E. coli DH5α, E. coli BL21(DE3). : 34 gambar; 1 tabel; 5 lampiran : 48 (1990-2011) Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 ABSTRACT Name : Arizah Kusumawati Study Program : Magister of Pharmacy Title : Cloning Human Interferon Alpha 2a Gene at pET-32b(+) Vector and Expression in Escherichia coli Interferon (IFN) is a cytokine produced by various cell types as a response of stimulation to viruses, bacteria, parasites, tumor cells, or other antigens. Interferon α type I IFN groups have various biological effects, including antiviral, antitumor and immunotherapeutic. The aim of this research is to synthesize recombinant human IFN α2a proteins through bacterial expression systems in E. coli BL21 (DE3). Addition genes of human IFN α2a, which are restriction enzyme cutting sites for Nco I and Xho I, are added through PCR method. This step is followed by ligation process to the pET-32b(+) vector and then transformed into E. coli DH5α. The recombinant vector (pET-32b(+)-IFN α2a) has a nucleotide right sequence after it was being sequenced, after was transformed into E. coli BL21 (DE3). Obtained transformant clones were cultured and induced by addition of IPTG 1 mM to produce the expression of recombinant human IFN α2a proteins. As result of isolation process, recombinant protein of human IFN α2a are collected in fused protein thus can simplify the detection and purification method. The proteins are characterized by the SDS PAGE method followed by Western blot and CBB staining. The results show that the recombinant human IFN α2a protein bands are exactly 36 kDa. The maximum expression results were obtained at 37⁰C with 5 hours incubation after induction process. Key Words xiv+74 pages Bibliography : Interferon, pET-32b(+) vector, E. coli DH5α, E. coli BL21(DE3) : 34 pictures; 1 tables; 5 appendices : 48 (1990-2011) Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 DAFTAR ISI HALAMAN JUDUL ……….......................................................................... i HALAMAN PERNYATAAN BEBAS PLAGIARISME ............................... ii HALAMAN PERNYATAAN ORISINALITAS ........................................... iii HALAMAN PENGESAHAN ........................................................................ iv KATA PENGANTAR ..................................................................................... v LEMBAR PERSETUJUAN PUBLIKASI ...................................................... vii ABSTRAK ....................................................................................................... viii ABSTRACT ..................................................................................................... ix DAFTAR ISI ………………………………………………….............…...... x DAFTAR GAMBAR ...………………………………………........................ xii DAFTAR TABEL ………………………………………............................... xiv DAFTAR LAMPIRAN …………………………………………................... xv BAB 1 PENDAHULUAN …………………………………........................ 1.1 Latar Belakang ......……………………………..….................... 1.2 Rumusan Masalah ………………………………..................… 1.3 Hipotesis ..................................................................................... 1.4 Tujuan Penelitian ........................................................................ 1.5 Manfaat Penelitian ...................................................................... 1 1 2 2 3 3 BAB 2 TINJAUAN PUSTAKA …………………….................….....……. 2.1 Interferon .............……....................................................…….... 2.2 Kloning dan Isolasi DNA ............................................................ 2.3 Vektor pET-32b(+) ...................................................................... 2.4 Manipulasi Enzimatis DNA ......................................................... 2.4.1 Enzim Endonuklease Restriksi .......................................... 2.4.2 Enzim DNA Ligase ........................................................... 2.4.3 Enzim DNA Polimerase ................................................... 2.5 Amplifikasi DNA ....................................................................... 2.6 Analisis DNA ............................................................................. 2.6.1 Pemisahan Menggunakan Gel ........................................... 2.6.2 Sekuensing DNA ............................................................... 2.7 Ekspresi Protein pada Escherichia coli ...................................... 2.8 Analisis Protein .......................................................................... 2.8.1 Western Blot ...................................................................... 2.8.2 Analisis Densitometri ........................................................ 4 4 8 11 13 13 14 15 15 17 17 18 20 23 23 23 BAB 3 METODE PENELITIAN ............................................................... 3.1 Tempat dan Waktu ..................................................................... 3.2 Bahan dan Alat .......................................................................... 3.2.1 Bahan ................................................................................ 3.2.2 Alat .................................................................................... 3.3 Tahap Kerja ................................................................................. 3.3.1 Penyiapan DNA Insert ...................................................... 3.3.2 Penyiapan Vektor pET-32b(+) .......................................... 25 25 25 25 26 26 26 29 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 3.3.3 Pembuatan Sel Kompeten E. coli ...................................... 3.3.4 Kloning Gen human ifn α2a ........................................... 3.3.5 Skrining dan Verifikasi Klon Transforman di E. coli DH5α ................................................................................. 3.3.6 Kultivasi dan Isolasi Vektor Rekombinan ........................ 3.3.7 Transformasi Vektor Rekombinan ke E. coli BL21(DE3) ........................................................................ 3.3.8 Skrining dan Verifikasi Klon Transforman di E. coli BL21(DE3) ........................................................................ 3.3.9 Ekspresi, Isolasi dan Karakterisasi Protein Rekombinan Human IFN α2a ................................................................. 29 30 31 31 34 34 35 BAB 4 HASIL DAN PEMBAHASAN ........................................................ 4.1 Penyiapan DNA Insert ................................................................ 4.2 Penyiapan Vektor pET-32b(+) .................................................. 4.3 Kloning Gen human ifn α2a .................................................... 4.4 Skrining dan Verifikasi Klon Transforman di E. coli DH5α .......................................................................................... 4.5 Kultivasi dan Isolasi Vektor Rekombinan .................................. 4.6 Transformasi Vektor Rekombinan ke E. coli BL21 BL21(DE3) ................................................................................. 4.7 Skrining dan Verifikasi Klon Transforman di E. coli BL21(DE3) ................................................................................. 4.8 Ekspresi, Isolasi dan Karakterisasi Protein Rekombinan Human IFN α2a .......................................................................... 39 39 42 44 BAB 5 KESIMPULAN DAN SARAN ......................................................... 5.1 Kesimpulan ................................................................................. 5.2 Saran ........................................................................................... 68 68 68 DAFTAR PUSTAKA .................................................................................... 69 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 46 48 53 54 56 DAFTAR GAMBAR Gambar 2.1 Gambar 2.2 Gambar 2.3 Gambar 2.4 Gambar 4.1 Gambar 4.2 Gambar 4.3 Gambar 4.4 Gambar 4.5 Gambar 4.6 Gambar 4.7 Gambar 4.8 Gambar 4.9 Gambar 4.10 Gambar 4.11 Gambar 4.12 Gambar 4.13 Gambar 4.14 Gambar 4.15 Gambar 4.16 Gambar 4.17 Gambar 4.18 Gambar 4.19 Gambar 4.20 Gambar 4.21 Gambar 4.22 Mekanisme Aksi Interferon ............................................................. Jalur Jak-Stat ………………………………………………….... Gen human interferon α2a …………………………………… ... Vektor pET-32b(+) ………...................................................... Elektroforesis Hasil PCR untuk Penyiapan DNA Insert ........... Elektroforesis Hasil Purifikasi Produk PCR dan Hasil Pemotongan Ganda dengan Enzim Nco I dan Xho I ................. Elektroforesis Hasil Purifikasi Metode Fenol Kloroform Setelah Pemotongan Ganda ....................................................... Elektroforesis Hasil Pemotongan Vektor pET-32b(+) ……… Elektroforesis Hasil Purifikasi Vektor pET-32b(+) Setelah Pemotongan Ganda dengan Enzim Xho I dan Nco I .................. Hasil Transformasi pada E. coli DH5α ...................................... Elektroforesis Hasil PCR dengan Primer IFN_NcoI_F dan IFN_XhoI_R pada Klon Transforman 1-11 E. coli DH5α ....... Elektroforesis Hasil PCR dengan Primer pET_Screen_F dan pET_Screen_R pada Klon Transforman 1-11 E. coli DH5α . ... Elektroforesis Hasil Miniprep Klon Transforman di E. coli DH5α ………………………………………………………. ... Elektroforesis Skrining Metode PCR Menggunakan Primer (IFN_NcoI_F dengan pET_Screen_R) dan (Primer pET_Screen_F dengan IFN_XhoI_R) pada Hasil Miniprep Klon 1, 2, 3, 4, dan Klon 11 E. coli DH5α ……………......... Elektroforesi Hasil Pemotongan Vektor Rekombinan Klon 2 .. Elektroforesis Hasil Qiagen Plasmid Maxi Kit ……………... Hasil Alignment Sekuen Klon 2 dengan Stag-18mer-primer .... Hasil Alignment Sekuen Klon 2 dengan T7terminatorprimer ……………………………………….….................... Hasil Transformasi Vektor Rekombinan pada E. coli BL21(DE3) ………………………………………………….. .. Elektroforesis Hasil PCR Skrining dengan Primer pET_Screen_F dan pET_Screen_R pada Klon Transforman E. coli BL21(DE3) ..................................................................... Elektroforesis Hasil PCR Skrining dengan Primer IFN_NcoI_F dan IFN_XhoI_R pada Klon Transforman E. coli BL21(DE3) …………………………………………….. Western Blot Klon 1 Inkubasi 37⁰C IPTG 1 mM …………... Pewarnaan CBB Klon 2, 3, 6, dan 7 Inkubasi 37⁰C IPTG 1 mM ....................................................................................... Pewarnaan CBB Klon 4 dan Klon 5 Inkubasi 37⁰C IPTG 1 mM ……………………………………………………....... Pewarnaan CBB Pelet Klon 3 Inkubasi 37⁰C IPTG 1 mM .... Pewarnaan CBB Supernatan Klon 3 Inkubasi 37⁰C IPTG 1 mM ……………………………………………………….. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 5 6 8 12 39 40 41 42 43 45 47 47 48 49 50 51 52 52 54 55 55 57 58 58 60 60 Gambar 4.23 Gambar 4.24 Gambar 4.25 Gambar 4.26 Gambar 4.27 Gambar 4.28 Gambar 4.29 Gambar 4.30 Western Blot Klon 3 Inkubasi 30⁰C IPTG 1 mM..................... Pewarnaan CBB Klon 3 Inkubasi 30⁰C IPTG 1 mM................ Pewarnaan CBB Pelet Klon 3 Inkubasi 28⁰C IPTG 1 mM....... Pewarnaan CBB Supernatan Klon 3 Inkubasi 28⁰C IPTG 1 mM …...……………………………………….................... Pewarnaan CBB Pelet Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM ...…………………......................................................... Western Blot Pelet Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM ……………………......................................................... Western Blot Supernatan Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM …….…….............................................................. Pewarnaan CBB Hasil Isolasi Protein dari Pelet …………….. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 61 61 62 62 63 63 64 66 DAFTAR TABEL Tabel 4.1 Hasil Kuantifikasi Tingkat Ekspresi pada Pelet dan Supernatan …………………….............................................. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 65 DAFTAR LAMPIRAN Lampiran 1 Lampiran 2 Lampiran 3 Lampiran 4 Lampiran 5 Skema Kerja Penelitian ........................................................... Vektor pcDNA3.1+:IFN-Gene ............................................... Hasil Sekuen Klon 2 E. coli DH5α dengan Stag-18merprimer ...................................................................................... Hasil Sekuen Klon 2 E. coli DH5α dengan T7terminatorprimer ...................................................................................... Nukleotida Human Interferon Alpha 2a (GenBank: DI084466.1) ………………………………………………… Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 73 74 75 77 79 BAB 1 PENDAHULUAN 1.1 Latar Belakang Interferon (IFN) adalah obat yang pada awalnya dikembangkan untuk terapi penyakit kanker, tetapi pada saat ini interferon juga digunakan untuk pengobatan berbagai penyakit. Terapi interferon telah digunakan secara luas untuk pengobatan hepatitis kronis (hepatitis B dan C), kaposi’s sarkoma, dan karsinoma lainnya (Tae-Ok Bae et al., 1995). Interferon α termasuk interferon tipe I dan merupakan obat yang potensial untuk dikembangkan karena tingginya kebutuhan interferon α dalam pengobatan. Interferon α memiliki berbagai efek biologis meliputi antiviral, antitumor dan juga sebagai immunoterapetik (Brassard, 2002). Interferon α2a merupakan bagian dari fokus produksi protein teraputik interferon α dengan teknologi DNA rekombinan. Beberapa penelitian sebelumnya tentang interferon α2a diantaranya adalah purifikasi dan karakterisasi rekombinan human interferon α2a yang diproduksi dari Saccharomyces cerevisiae (Tae-Ok Bae et al., 1995), dan efek heat shock produksi rekombinan human interferon α2a pada Escherichia coli galur MSD462 dengan vektor ekspresi pZe0148 (Roy et al., 2005). Pada penelitian ini akan dilakukan kloning gen human interferon α2a pada vektor pET-32b(+) dan ekspresinya menggunakan inang Escherichia coli galur BL21(DE3). Sistem pET merupakan sistem yang didesain untuk kloning dan ekspresi tingkat tinggi pada E. coli. Vektor pET-32b(+) dilengkapi dengan multipel situs restriksi, fusi sekuen thioredoxin (Trx.Tag), histidin (His•Tag), S•Tag, serta situs pemotongan thrombin dan enterokinase (pET Sistem Manual, 2006). Protein yang dihasilkan dalam bentuk protein terfusi sehingga dapat memudahkan proses deteksi dan purifikasi protein rekombinan. Tag thioredoxin pada vektor pET32b(+) berfungsi untuk mendukung pembentukan ikatan disulfida, meningkatkan kelarutan protein, serta melindungi protein rekombinan terhadap agregasi yang tidak diinginkan selama ekspresi. Tag histidin kedepannya akan digunakan untuk purifikasi protein rekombinan. Situs pemotongan enterokinase yang terdapat di 1 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 vektor pET-32b(+) kedepannya akan digunakan untuk pemisahan antara fusi tag dengan protein target setelah proses purifikasi. Sebagian besar produk farmasi yang telah dipasarkan pada kelompok rekombinan interferon α diproduksi dan dipurifikasi dari E. coli (Rabhi-Essafi, 2007). Inang E. coli galur BL21(DE3) dipilih sebagai sistem ekspresi karena mempunyai berbagai keuntungan. Escherichia coli BL21(DE3) merupakan bakteri yang umum untuk ekspresi protein yang menggunakan promotor T7. DE3 menunjukkan bahwa inang merupakan lisogen dari α prophage (DE3), oleh karena itu membawa salinan kromosom gen T7 RNA polimerase di bawah kontrol promotor lacUV5 yang diinduksi dengan penambahan isopropyl-1-thio-β-Dgalactopyranoside (IPTG). T7 RNA polimerase diekspresikan pada saat penambahan IPTG yang mana akan menginduksi level ekspresi tinggi protein pada vektor ekspresi dengan promotor T7 (Studier, 1991). Escherichia coli BL21(DE3) merupakan inang umum yang digunakan untuk ekspresi protein karena defisiensi protease Lon dan OmpT, yang mana kedua protease tersebut dapat menginduksi proteolisis dari protein yang dioverekspresi (Sørensen dan Mortensen, 2005). 1.2 Rumusan Masalah Interferon α2a adalah salah satu agen terapetik berbasis bioteknologi yang dibutuhkan dan mempunyai nilai ekonomi yang tinggi. Rekombinan interferon α2a yang beredar dan digunakan di indonesia merupakan produk impor. Oleh karena itu, sangat perlu adanya pengembangan sistem produksi rekombinan interferon α2a di dalam negeri untuk meningkatkan kemampuan produksi bahan obat berbasis bioteknologi serta mengurangi ketergantungan dari luar negeri. Pada penelitian ini dilakukan kloning gen human interferon α2a pada vektor pET32b(+) dan ekspresi pada inang sel E. coli BL21(DE3) untuk menghasilkan protein rekombinan human interferon α2a. 1.3 Hipotesis Sintesis protein rekombinan human interferon α2a dapat dilakukan dengan menggunakan gen sintetik human interferon α2a yang dikloning pada vektor pET- Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 32b(+) dan diekspresikan menggunakan sistem ekspresi prokariotik pada sel bakteria E. coli BL21(DE3). 1.4 Tujuan penelitian Tujuan umum pada penelitian ini adalah untuk mensintesis protein rekombinan human interferon α2a melalui sistem ekspresi pada bakteria E. coli BL21(DE3). Adapun tujuan khusus pada penelitian ini yaitu untuk mendesain vektor rekombinan pET-32b(+)-IFN α2a dan melakukan analisis ekspresi protein rekombinan human interferon α2a pada inang ekspresi E. coli BL21(DE3). 1.5 Manfaat Penelitian Manfaat dari penelitian ini yaitu dihasilkannya protein rekombinan human interferon α2a sehingga dapat diuji lebih lanjut untuk dapat digunakan dalam pengobatan. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 BAB 2 TINJAUAN PUSTAKA 2.1 Interferon Interferon (IFN) termasuk kelompok glikoprotein yang diproduksi oleh berbagai tipe sel sebagai respon terhadap rangsangan yang diterima oleh sel. Rangsangan tersebut bisa disebabkan oleh virus, bakteri, parasit, sel tumor, atau antigen lain. Interferon mempunyai berat molekul sekitar 20-30 kDa. Interferon juga termasuk dalam golongan sitokin seperti interleukin (ILS), colonystimulating factors dan growth factors. Sitokin bekerja pada reseptor spesifik di permukaan sel dan berfungsi sebagai pengatur kelangsungan hidup sel, proliferasi sel, diferensiasi dan aktivasi fungsional sel (Obeid dan Bouvois, 2006). Interferon berdasarkan tipe reseptornya dikelompokkan menjadi 2 tipe. Interferon tipe I berikatan pada reseptor tipe 1, yaitu IFN alpha (α), IFN beta (β), IFN omega (ω), dan IFN tao (τ). Interferon tipe 2 berikatan pada reseptor tipe 2, yaitu IFN gamma (γ). Hampir semua tipe sel memproduksi IFN tipe I. Interferon tipe II hanya diproduksi oleh sel limfosit T dan NK-cells (Natural Killer Cells) (Jonasch dan Haluska, 2001). Interferon memiliki aktivitas spektrum luas dan mekanisme kerjanya melalui interaksi yang komplek. Interferon mempunyai aktivitas antivirus, antitumor, berpengaruh pada metabolisme dan diferensiasi sel serta memodulasi sistem imun (Jonasch dan Haluska, 2001). Interferon dapat mencegah replikasi virus pada sel, serta dapat mengaktifkan fungsi khusus dari sel meliputi deferensiasi, pertumbuhan, pengekspresian antigen permukaan dan immunoregulasi sel (Meager, 2006). Aktivitas antivirus IFN melalui mekanisme pencegahan replikasi pada sel-sel sekitar yang terinfeksi. Pencegahan replikasi dilakukan melalui pengikatan IFN pada reseptor permukaan membran sel yang mengaktifkan gen-gen pengkode protein yang menghalangi replikasi virus. Ekspresi gen pengkode IFN terjadi melalui jalur transduksi sinyal dan aktivasi transkripsi yang dikenal dengan jalur Jak-Stat. Interferon α dan β berikatan pada tipe reseptor yang sama, sedangkan IFN γ berikatan pada tipe reseptor yang berbeda (Samuel, 2001). 4 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Mekanisme aksi dari IFN dimulai dengan zat penginduksi IFN akan memicu sel untuk mengaktifkan gen ifn sehingga dihasilkan mRNA yang kemudian ditranslasikan menjadi protein IFN. Protein IFN selanjutnya disekresikan keluar sel. Interferon ekstraseluler akan terikat ke reseptor pada membran sel sekitarnya. Proses pengikatan IFN pada reseptor ini akan menginisiasi sinyal kaskade JAK/STAT yang selanjutnya akan menstimulasi ekspresi gen penghasil protein efektor. Protein efektor akan memediasi efek IFN sebagai antivirus, anti tumor, dan imunomodulator (Pang et al., 2005). Sel yang teraktivasi juga dapat menghasilkan protein aktivator bagi sel lain sehingga menghasilkan protein efektor (Samuel, 2001). Gambar 2.1 Mekanisme Aksi Interferon (Pang et al., 2005). Jalur transduksi sinyal untuk aktivasi transkripsi dan pengekspresian gen pengkode ifn yaitu melalui jalur Jak-Stat. Protein Signal Transducer and Activator of Transcription (STAT) merupakan faktor transkripsi yang dapat difosforilasi pada residu asam amino tirosin oleh enzim tirosin kinase Janus Family of Tyrosine Kinase (JAK). Protein STAT terdiri atas tujuh macam yaitu Stat-1, Stat2, Stat-3, Stat-4, Stat-5a, Stat-5b dan Stat-6. Protein JAK terdiri 4 macam yaitu Jak-1, Jak-2, Jak-3 dan Tyk-2 (Samuel, 2001). Proses transduksi sinyal diinisiasi oleh pengikatan IFN pada subunit reseptor tirosin kinase. Pengikatan IFN akan mengaktivasi faktor transkripsi Jak dan Stat melalui fosforilasi tirosin. Kinase Jak-1 dan Tyk-2 yang teraktivasi oleh Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 IFN α dan IFN β akan menghasilkan fosforilasi dan dimerisasi protein Stat-1 (p91) dan Stat-2 (p113) yang selanjutnya ditranslokasi bersama IRF-9 (p48) ke inti sel. Komplek ketiga protein ini disebut dengan IFN-Stimulated Gene Factor 3 (ISGF 3) yang dapat mengaktifkan gen-gen pengkode ifn α dan β melalui IFNStimulted Response Element (ISRE). Pada IFN γ kinase Jak-1 dan Jak-2 yang teraktivasi akan memfosforilasi dan menyebabkan homodimerisasi protein Stat-1 yang kemudian ditranslokasi ke inti sel. Komplek dimer ini disebut dengan Jak-1 dan Gamma Activation Factor (GAF) yang mengaktifkan gen-gen pengkode ifn γ. Komplek GAF akan mengaktivasi gen-gen IFN γ melalui elemen GammaActivated Sequence (GAS) (Samuel, 2001). Gambar 2.2 Jalur Jak-Stat (Samuel, 2001). Interferon α dan β dibentuk oleh berbagai sel sebagai reaksi terhadap infeksi viral, bakteri, mikoplasma dan protozoa. Interferon α diproduksi oleh leukosit dan IFN β dipoduksi oleh fibroblas. Kedua IFN tersebut sama-sama dikodekan oleh gen yang terletak pada kromosom 9 manusia. Interferon α dan β berfungsi sebagai virustatis yaitu mencegah infeksi lebih lanjut dengan cara menduduki reseptor-reseptor khas pada membran sel normal sehingga tidak dapat Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 dipenetrasi oleh virus. Kedua IFN tersebut juga bersifat sitostatis yaitu menghambat pertumbuhan sel-sel tumor dan menstimulasi makrofag serta NKcells (Natural Killer Cells) yang mana dapat mendeteksi sel-sel tumor dan sel-sel yang diinvasi virus dan kemudian menghancurkannya. Interferon α digunakan untuk pengobatan pada hepatitis dan jenis leukimia tertentu. Interferon β digunakan pengobatan pada penyakit multiple sclerosis (Tan Hoan Tjay, 2007). Interferon gamma (IFN γ) diproduksi oleh limfosit T dan NK-cells dengan gen pengkode ifn γ terletak pada kromosom 12 manusia. Gen ifn γ memiliki 3 intron dan terdiri dari 146 asam amino dengan kandungan karbohidrat 30% (Arbabi, 2003). Interferon γ berfungsi sebagai immunostimulator (Tan Hoan Tjay, 2007). Interferon γ memiliki daya antiviral lebih lemah dibandingkan IFN α dan IFN β. Aktivitas biologis dari IFN γ juga dapat mencegah pertumbuhan sel neoplastik (Arbabi, 2003). Interferon α merupakan glikoprotein yang terdiri atas 165 asam amino dalam bentuk 2a dan 2b dengan masing-masing asam amino lisin dan arginin pada posisi 23. Interferon α memiliki dua ikatan disulfida yaitu antara Cys1 ke Cys98 dan Cys29 ke Cys138 (Tae-ok Bae et al., 1995). Interferon β adalah glikoprotein yang terdiri atas 165 asam amino dalam bentuk 1a dan 1b dengan masing-masing asam amino sistein dan serin pada posisi 17. Interferon γ adalah glikoprotein yang terdiri atas 140 asam amino dengan bentuk 1a dan 1b yang memiliki masingmasing asam amino glutamin atau arginin diposisi 137 (Tan Hoan Tjay, 2007). Interferon α2a merupakan subtipe dari IFN α yang terdiri atas 165 asam amino sistein dengan 4 sistein dan 2 ikatan disulfida. Interferon α2a adalah glikoprotein dengan berat molekul sekitar 19.000 dalton (Tae-Ok Bae et al., 1995). Struktur 3 dimensi protein IFN α2a yang ditentukan menggunakan spektroskopi nuclear magnetic resonance (NMR) pada pH 3,5 menunjukkan bahwa IFN α2a sebagian besar mengandung alpha heliks (6 alpha heliks, 65% alpha heliks) dalam struktur sekunder dan sisanya adalah kumparan koil dan loop (Klaus et al., 1997). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Gambar 2.3 Gen human interferon α2a. (http://www.roche-australia.com/fmfiles/re7229005/downloads/anti-virals/roferon-pi.pdf) Penggunaan IFN α2a telah disetujui oleh FDA (Food and Drug Administration) untuk pengobatan non-hodgkin’s lymphoma (NHL), hairy cell leukemia, chronic myelogenous leukemia (CML), AIDS-related kaposi’s sarcoma, dan hepatitis C kronis (Jonasch dan Haluska, 2001). Banyak upaya yang telah dilakukan untuk kloning dan ekspresi rekombinan human IFN α2a pada berbagai mikroorganisme. Beberapa studi mengenai IFNα2a diantaranya dilakukan oleh Tae-Ok Bae et al. (1995) purifikasi dan karakterisasi rekombinan human IFN α2a yang diproduksi dari Saccharomyces cerevisiae, dan Roy et al. (2005) efek heat shock produksi rekombinan IFN α2a pada Escherichia coli galur MSD462 dengan vektor ekspresi pZe0148. Pada penelitian ini akan dilakukan kloning gen human ifn α2a pada vektor pET-32b(+) dan ekspresinya menggunakan inang E. coli galur BL21(DE3). 2.2 Kloning dan Isolasi DNA Kloning DNA (deoxyribonucleic acid) merupakan proses penggandaan jumlah DNA rekombinan yang dapat melalui proses perkembangbiakan sel bakteri, sel ragi, sel mamalia (Muladno, 2010). Kloning DNA merupakan teknik Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 DNA rekombinan yang memungkinkan untuk dibuatnya satu DNA tertentu dalam jumlah besar dan murni. Metode kloning DNA melibatkan ligasi sekuen tertentu misalnya gen manusia ke dalam vektor. Vektor merupakan urutan DNA yang terdapat di dalam satu sel, tetapi bukan merupakan bagian dari genom sel. Pembawa kloning dengan DNA yang diklon di dalamnya akan diperbanyak dalam sel inang untuk menghasilkan sejumlah besar DNA dengan urutan spesifik yang kemudian dilakukan isolasi untuk studi selanjutnya (Grompe et al., 1998). Vektor adalah DNA untai ganda sirkuler yang ukurannya kecil (2000-5000 pb) yang dapat bereplikasi di dalam sel inang. Vektor berdasarkan jumlah salinannya dibedakan menjadi dua yaitu vektor dengan jumlah salinan tinggi dan salinan rendah. DNA vektor murni dapat dipisahkan dari DNA genom sel inang, karena DNA vektor memiliki ukuran jauh lebih kecil. Ukuran DNA genom sel inang memiliki ukuran yang besar misalnya pada genom E. coli 1x106 pb. Vektor memiliki origin of replication (ORI) sendiri sehingga mampu memperbanyak diri dengan bantuan mesin-mesin seluler endogen. Vektor kloning juga memiliki marka untuk seleksi yang umumnya berupa gen yang membawa resistensi terhadap antibiotika tertentu. Hanya klon rekombinan yang mengandung vektor maka yang akan mampu tumbuh dalam media yang mengandung antimikroba tertentu (Grompe et al., 1998). Vektor modern saat ini telah direkayasa sehingga memiliki beberapa situs restriksi (polylinker sites). Vektor pada saat sekarang ini telah dibuat mempunyai jumlah salinan tinggi dan memiliki ukuran 3 kb sehingga dapat menerima DNA sisipan sampai 15 kb. Kebanyakan DNA yang diklon ke dalam vektor berukuran relatif kecil kurang dari 10 kb, karena ukuran DNA yang jauh lebih besar biasanya tidak stabil. Oleh karena itu, juga dikembangkan vektor kloning yang dapat membawa DNA dengan ukuran besar diantaranya vektor kloning faga (phage), kosmid, Bacterial Artificial Chromosomes (BAC) dan Yeast Artificial Chromosomes (YAC) (Grompe et al., 1998). Prosedur kloning suatu gen tertentu didahului dengan pemotongan DNA vektor dengan menggunakan enzim restriksi, sehingga DNA vektor sirkuler terbuka dan menjadi linier. DNA untai ganda linier lain yang diinginkan dibuat dengan ujung-ujungnya sesuai untuk diligasikan ke dalam celah sehingga Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 terbentuk kembali vektor sirkuler yang mengandung insert. Vektor rekombinan yang diperoleh kemudian akan dimasukkan ke dalam sel inang melalui proses transformasi (Grompe et al., 1998). Proses pemasukan molekul DNA ke dalam sel disebut transformasi karena masuknya molekul DNA ke dalam sel dapat mengubah fenotip sel tersebut (Muladno, 2010). Proses transformasi pada bakteri dapat dilakukan dengan cara memanaskan campuran DNA dan sel bakteri yang kompeten pada suhu 42⁰C selama 1 menit (heat shock) atau dengan memberikan aliran listrik (electroporation), sehingga DNA vektor rekombinan dapat melalui dinding bakteri kemudian masuk ke dalam sel bakteri. Setelah proses transformasi selesai, bakteri ditumbuhkan pada media agar yang mengandung marka antibiotik. Pada kondisi ini, hanya sel yang mengandung vektor rekombinan yang dapat tumbuh dan membentuk koloni. Tiap koloni bakteri ini disebut klon, tiap klon diperbanyak dalam media cair untuk dilakukan isolasi DNA dalam jumlah besar. Klon yang mengandung vektor rekombinan merupakan sumber DNA yang permanen, karena sel bakteri tidak mati dan dapat terus menerus membelah tanpa batas (Grompe et al., 1998). Isolasi DNA terdiri dari beberapa tahap yaitu (1) kultivasi sel dalam media yang sesuai; (2) pemecahan dinding sel; (3) ekstraksi DNA; (4) purifikasi DNA. Proses pemecahan dinding sel bakteri dapat dilakukan secara fisik dengan cara sonikasi maupun secara kimia dengan menggunakan enzim lisozim, etilendiamin tetra asetat (EDTA), dan sodium diodesil sulfat (SDS). Seteleh dinding sel lisis, dilanjutkan pemisahan debris sel menggunakan metode disentrifus. Metode disentrifus digunakan untuk memisahkan antara komponen sel yang tidak larut akan mengendap dan ekstrak sel dalam supernatan yang jernih. Ekstrak sel dalam supernatan selain mengandung DNA juga masih mengandung protein dan RNA, sehingga perlu untuk dimurnikan (Radji, 2011). Proses pemurnian DNA yang umumnya dilakukan yaitu dengan penambahan larutan fenol ataupun dengan campuran fenol kloroform dengan perbandingan 1:1, untuk mengendapkan protein dengan cara disentrifus dan dihancurkan secara enzimatis dengan proteinase. Pemurnian DNA selanjutnya dengan penambahan RNAse untuk membersihkan DNA dari RNA. Molekul DNA Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 yang diperoleh kemudian dilakukan presipitasi menggunakan etanol absolut disimpan pada suhu -20⁰C, kemudian disentrifus untuk mengendapkan DNA dan mempermudah pemisahan (Radji, 2011). 2.3 Vektor pET-32b(+) Sistem pET adalah sistem unggul yang dikembangkan untuk kloning dan ekspresi protein rekombinan pada E. coli. Studier et al. (1990) pertama kali menjelaskan tentang sistem ekspresi pET, yang mana telah dikembangkan untuk berbagai aplikasi ekspresi. Lebih dari 40 jenis vektor pET yang berbeda tersedia secara komersial (pET Sistem Manual, 2006). Sistem pET mencakup promotor hibrid, multipel situs kloning yang tergabung dengan fusi yang berbeda, situs pemotongan protease, dan modifikasi latar belakang genetik dari vektor pBR322 untuk berbagai tujuan ekspresi (Sørensen dan Mortensen, 2005). Vektor pET-32b(+) merupakan sistem yang dilengkapi dengan multipel situs restriksi dan terdapat fusi sekuen thioredoxin (Trx.Tag), histidin (His•Tag), S•Tag, serta situs pemotongan thrombin dan enterokinase (pET Sistem Manual, 2006). Protein yang dihasilkan dalam bentuk protein terfusi sehingga memudahkan untuk proses deteksi dan purifikasi protein rekombinan. Tag thioredoxin berfungsi untuk mendukung pembentukan ikatan disulfida dan meningkatkan kelarutan protein. Tag histidin 6x pada terminal N dan terminal C berfungsi untuk deteksi protein target menggunakan metode Western blot. Tag histidin juga dapat digunakan untuk purifikasi protein target menggunakan Immobilized metal-affinity chromatography (IMAC) dengan matriks Ni2+-NTA, Co2+-CMA (Talon) dan dielusi menggunakan imidazol (Terpe, 2002). Fusi peptida S•Tag dapat digunakan untuk uji kuantitatif, deteksi menggunakan metode Western blot serta purifikasi protein rekombinan berdasarkan metode pemisahan afinitas. Situs pemotongan enterokinase digunakan untuk pemisahan antara fusi tag dan protein target setelah proses purifikasi. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 pET-32b(+) (5899 pb) Gambar 2.4 Vektor pET-32b(+). (https://wasatch.biochem.utah.edu/chris/links/pET32.pdf) Sistem pET dapat mengekspresikan protein tingkat tinggi karena mekanisme ekspresinya dikontrol dengan menggunakan promotor T7lac, inang Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 dengan pylsS, serta penambahan glukosa ke media berdasarkan karakteristik protein target. PlysS pada inang ekspresi berfungsi untuk meningkatkan toleransi λDE3 lisogen untuk vektor dengan insert yang toksik, serta vektor menjadi lebih stabil. Vektor pET-32b(+) memiliki promotor T7lac dengan sekuen lac operator downstream dari promotor T7. Vektor juga membawa sekuen pengkode untuk represor lac (lacI) dengan orientasi dari promotor T7lac dan lacI berbeda. Vektor apabila digunakan dalam DE3 lisogen, maka represor lac beraksi di promotor lacUV5 dalam kromosom inang untuk menekan transkripsi gen T7 RNA polimerase dan juga pada promotor T7lac dalam vektor untuk memblokir transkripsi gen target (pET Sistem Manual, 2006). Vektor rekombinan ditransformasikan ke dalam galur sel E. coli yang kromosomnya mempunyai salinan gen untuk T7 RNA polimerase (T7 gen 1) untuk memproduksi protein. Inang dengan galur DE3 lisogen dipilih karena defisiensi protease, auksotrof asam amino, dapat meningkatan kelarutan, dan suplementasi kodon yang jarang (rare codon) (pET Sistem Manual, 2006). Escherichia coli galur BL21(DE3) adalah inang yang umum untuk ekspresi protein yang menggunakan promotor T7. DE3 menunjukkan bahwa inang merupakan lisogen dari α prophage (DE3), oleh karena itu membawa salinan kromosom dari gen T7 RNA polimerase di bawah kontrol promotor lacUV5 yang diinduksi dengan penambahan isopropyl-1-thio-β-D-galactopyranoside (IPTG). Escherichia coli BL21(DE3) adalah inang yang paling banyak digunakan untuk ekspresi protein karena defisiensi protease Lon dan OmpT (Sørensen dan Mortensen, 2005). 2.4 Manipulasi Enzimatis DNA Manipulasi DNA dapat dilakukan secara enzimatis dengan menggunakan berbagai macan enzim diantaranya yaitu : 2.4.1 Enzim Endonuklease Restriksi Enzim endonuklease restriksi umumnya diperoleh dari bakteri, yang mana dapat mengenal DNA untai ganda dengan urutan basa spesifik serta memotong pada tempat yang tertentu. Enzim endonuklease restriksi berfungsi untuk membatasi masuknya DNA asing dengan cara memotong DNA pada situs Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 restriksi yang tidak terdapat pada bakteri. Setiap enzim endonuklease restriksi dapat mengenal, mengikat dan memotong urutan basa tertentu. Urutan basa yang dipotong biasanya terdiri dari 4-8 pasang basa (pb), dan enzim endonuklease restriksi hanya memotong DNA yang mempunyai urutan basa yang benar-benar persis dengan kode tersebut. Sebagai contohnya yaitu urutan basa yang dikenal oleh enzim EcoRI adalah GAATTC. Semua situs yang memiliki urutan basa GAATTC akan dipotong oleh enzim EcoR1, namun jika ada satu basa yang berbeda misalnya CAATTC maka mampu mencegah terjadinya pemotongan. Proses pemotongan DNA dengan enzim endonuklease restriksi dapat menghasilkan potongan asimetris 5' atau 3' terbuka sehingga ujungnya kohesif (sticky end), ataupun potongan simetris sehingga ujungnya tumpul (blunt end) (Grompe et al., 1998). Enzim endonuklease restriksi diberi nama sesuai dengan bakteri penghasilnya, misalnya EcoRI berasal dari Escherichia coli. Penggunaan enzim restriksi adalah dengan menginkubasi enzim bersama-sama dengan DNA dalam larutan bufer yang sesuai, sehingga akan dihasilkan potongan-potongan DNA dengan ukuran tertentu. Pemotongan enzim restriksi bersifat spesifik untuk urutan basa tertentu, sehingga DNA yang berbeda akan menghasilkan pola pemotongan yang khas. Pola yang khas ini dapat diketahui setelah dilakukan elektroforesis gel dan pewarnaan sehingga fragmen DNA yang berbeda-beda ukurannya dapat terlihat (Grompe et al., 1998). Enzim endonuklease restriksi sangat bermanfaat diaplikasian pada teknologi DNA rekombinan karena sangat selektif dalam memotong untai DNA (Radji, 2011). 2.4.2 Enzim DNA Ligase Enzim DNA ligase adalah enzim yang berfungsi mengikatkan dua potong DNA secara kovalen (Grompe et al., 1998). Enzim DNA ligase merupakan enzim yang terdapat pada sistem biologis yang dapat mengkatalis pembentukan ikatan kovalen yang merekatkan kembali potongan fragmen-fragmen DNA. Enzim DNA ligase adalah salah satu enzim yang sangat penting dalam perkembangan teknologi DNA rekombinan (Radji, 2011). Enzim DNA ligase ini terutama digunakan untuk menyisipkan sepotong DNA ke dalam vektor kloning. Enzim DNA ligase yang paling banyak digunakan adalah T4 DNA ligase. Reaksi ligasi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 terjadi antara gugus 3'OH dari salah satu untai DNA dengan gugus fosfat dari untai DNA pasangannya. Reaksi ligasi menggunakan ATP sebagai sumber energi (Grompe et al., 1998). Proses ligasi menggunakan T4 DNA ligase memerlukan kofaktor ATP yang kemudian membentuk kompleks enzim-AMP. Kompleks ini menempel dan menyambung gugus 5'-fosfat dan 3'-hidroksi dengan ikatan kovalen sehingga terbentuk rantai fosfodiester (Sudjadi, 2008). Ligasi DNA hanya dapat terjadi antara dua ujung fragmen DNA yang kompatibel. Kompatibilitas inilah yang digunakan untuk mengatur ligasi beberapa fragmen DNA yang berbeda secara sekaligus dalam satu campuran reaksi (Grompe et al., 1998). Proses penyambungan fragmen DNA dengan ujung kohesif dengan enzim DNA ligase cukup efisien dan telah banyak digunakan dalam pembuatan vektor rekombinan. Pada fragmen DNA dengan ujung tumpul memerlukan kondisi ligasi yang spesial, yaitu kadar fragmen DNA harus tinggi dan larutan reaksi agak kental sehingga gerakan molekul lebih lambat (Sudjadi, 2008). 2.4.3 Enzim DNA Polimerase Enzim DNA polimerase adalah enzim yang menggunakan satu DNA untai tunggal sebagai cetakan (template) untuk mensintesis untai kedua yang komplemen dengan cetakannya. Enzim DNA polimerase membutuhkan primer dalam mensintesis DNA baru dari arah 5' ke 3'. Primer adalah sepotong kecil DNA yang komplemen terhadap untai DNA yang akan disalin, polimerase melekat pada primer dan kemudian akan memperpanjang primer dengan cara menambahkan blok-blok deoksinukleotida dari arah 5' ke 3' (Grompe et al., 1998). Primer merupakan oligonukleotida rantai pendek yang dibuat secara sintetik dan umumnya memiliki panjang 15-32 pasang basa. Enzim DNA polimerase memiliki aktivitas optimal pada suhu 92-95⁰C dan digunakan dalam polymerase chain reaction (PCR) (Radji, 2011). 2.5 Amplifikasi DNA Amplifikasi atau perbanyakan DNA untai ganda selain dengan menggunakan vektor kloning dalam sel inang hidup juga dapat dilakukan dengan metode polymerase chain reaction (PCR). Dengan menggunakan metode PCR Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 mampu diperoleh DNA dalam jumlah besar dan murni secara kimia. Metode PCR memungkinkan untuk dibuat sejumlah besar DNA dengan urutan basa tertentu dalam waktu yang singkat dan tanpa kloning. Sejumlah kecil DNA untai tunggal dapat disintesis dari nukleotida tunggal dengan bantuan oligonukleotida (Grompe et al., 1998). Metode PCR merupakan suatu proses sintesis enzimatik untuk mengamplifikasi fragmen DNA secara in vitro. Teknik PCR dapat meningkatkan jumlah fragmen DNA hingga mencapai 106-107 kali dalam waktu singkat (Radji, 2011). Prosedur PCR menggunakan DNA polimerase yang tahan terhadap panas (thermostable) dan dua primer oligonukleotida sintetik. Urutan DNA gen yang akan diamplifikasi harus diketahui sebagian sehingga dapat dibuat primer yang membatasi ujung DNA target. Prosedur PCR terdiri dari 3 tahap yaitu: (1) denaturasi DNA untai ganda, (2) penempelan primer ke DNA (annealing), dan (3) memperpanjang primer (extension) (Grompe et al., 1998). Ketiga tahapan siklus tersebut diatas diulang sesuai dengan jumlah siklus amplifikasi yang akan dilakukan (Radji, 2011). Pada tahap denaturasi terjadi pemanasan pada suhu 94-98⁰C, yang mana bertujuan untuk memisahkan untai ganda DNA target menjadi untai tunggal. Untai tunggal lalu siap dihibridisasikan dengan oligonukleotida yang sesuai pada proses annealing. Pada proses annealing temperatur diturunkan sampai di bawah titik leleh hibrid DNA-oligonukleotida yaitu pada suhu sekitar 50-68⁰C. Tahapan selanjutnya yaitu setelah primer menempel maka enzim DNA polimerase dapat menyalin urutan DNA untai tunggal pada temperatur 72⁰C (extension), dengan deoksinukleotida trifosfat (dNTP) yang tersedia. Setiap untai DNA target yang ada sejak awal akan disalin dan jumlah DNA akan menjadi dua kali lipat pada akhir tiap siklus. Jadi, setelah 25-30 siklus maka urutan DNA yang diinginkan diperbanyak sampai jutaan kali lipat. DNA hasil PCR dapat divisualisasikan sebagai pita (band) pada gel agarosa dengan pengecatan menggunakan etidium bromida (EtBr), dan selanjutnya dapat digunakan untuk kloning ataupun sekuensing DNA. Perubahan temperatur berturut-turut pada PCR dilakukan pada mesin thermocycler. Satu kali proses PCR biasanya selesai dalam jangka waktu 3 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 jam. Oleh karena itu, metode PCR merupakan metode tercepat untuk membuat DNA tertentu untuk dilakukan studi lebih lanjut (Grompe et al., 1998). Keterbatasan dari metode PCR diantaranya yaitu adanya salah menambahkan nukleotida oleh enzim Taq polimerase (Taq error) dan ketidakmampuan untuk mengamplifikasi fragmen DNA yang besar. Modifikasi Taq generasi baru yang mampu mengamplifikasi sampai 20 kb dengan tingkat fidelitas tinggi dapat mengurangi kesalahan PCR dan meningkatkan kemampuan amplifikasi. Kegunaan PCR diantaranya yaitu: (1) untuk mengamplifikasi DNA pada daerah tertentu sebagai strategi rekayasa genetik; (2) memperbanyak DNA genom untuk keperluan diagnostik; (3) memperbanyak DNA untuk sekuensing; (4) untuk memnyisipkan atau membuat mutasi (site directed mutagenesis); (5) dikombinasikan dengan reverse transcription untuk mengkuantifikasi jumlah mRNA awal (RT-PCR) (Grompe et al., 1998). 2.6 Analisis DNA Analisis DNA dapat dilakukan dengan menggunakan berbagai metode diantaranya yaitu : 2.6.1 Pemisahan Menggunakan Gel Teknik dasar pemisahan DNA dan RNA menggunakan elektroforesis pada sistem gel berdasarkan prinsip perbedaaan ukuran. DNA dan RNA pada pH netral bermuatan negatif dan akan bermigrasi ke arah anoda. Pada saat migrasi pada matriks polimer gel, maka fragmen kecil akan bergerak lebih cepat daripada fragmen yang besar. Migrasi elektroforetik melalui gel akan dapat memisahkan campuran fragmen DNA sehingga tampak sebagai pita-pita yang berbeda ukurannya. Matriks gel yang dapat dipakai untuk pemisahan asam nukleat diantaranya yaitu gel agarosa dan gel poliakrilamid. Gel agarosa dapat digunakan untuk pemisahan fragmen DNA berukuran 0.1-20 kb, sedangkan poliakrilamid untuk fragmen DNA berukuran 0.025-2 kb (Grompe et al., 1998). Gel agarosa mempunyai daya pemisahan lebih rendah dibandingkan dengan gel poliakrilamid, tetapi mempunyai rentang pemisahan yang lebih besar. Gel agarosa biasanya dilakukan dalam konfigurasi horisontal dalam kekuatan medan listrik dan arah tetap (Sudjadi, 2008). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Gel agarosa dibuat dengan melarutkan serbuk agarosa dalam bufer kemudian dipanaskan dan lalu dituang pada cetakan dan didiamkan sampai dingin. Setelah dingin dan mengeras, agarosa membentuk matriks dengan kerapatan yang ditentukan oleh konsentrasi agarosa. Jika medan magnet diberikan antara kedua ujung gel, maka DNA yang bermuatan negatif pada pH netral dan bergerak ke anoda. Kecepatan migrasi dari DNA tergantung dari ukuran (panjang) DNA, konformasi DNA, konsentrasi agarosa, dan besaran tegangan yang digunakan (Sudjadi, 2008). Fragmen DNA yang telah dipisahkan dengan elektroforesis gel dapat divisualisasikan dengan cara pewarnaan gel setelah elektroforesis selesai sehingga fragmen DNA dapat terlihat. Pewarna yang biasanya digunakan adalah etidium bromida yang mana dapat mengikat DNA dan akan berfluoresensi merah dibawah sinar UV (Grompe et al., 1998). Akrilamid merupakan suatu monomer yang digunakan dalam pembuatan gel akrilamid, yang ditambahkan amonium persulfat dan distabilkan oleh TEMED, maka terjadi reaksi berantai sehingga monomer terpolimerisasi menjadi rantai panjang. Jika dalam reaksi terdapat bisakrilamid, maka reaksi menjadi bersambung melintang menjadi gel yang pori-porinya ditentukan oleh panjang rantai dan jumlah sambungan silang. Gel poliakrilamid dibuat dengan cara menuangkan antara dua lempeng kaca yang dipisahkan dengan pembatas dengan ketebalan tertentu. Elektroforesis gel poliakrilamid yang digunakan untuk memisahkan protein dengan sistem vertikal (Sudjadi, 2008). 2.6.2 Sekuensing DNA Sekuensing adalah analisis DNA yang paling detil karena menggunakan penentuan urutan basa-basa nukleotida. Teknik sekuensing DNA merupakan salah satu teknik yang penting dalam teknologi DNA rekombinan karena sangat bermanfaat untuk menentukan urutan basa nukleotida suatu gen ataupun sekuen genom total suatu sel atau organisme. Teknik sekuensing DNA untuk pengurutan basa DNA terdiri 2 macam cara yaitu (1) cara degradasi kimiawi oleh A. Maxam dan W. Gilbert di Amerika, dan (2) cara determinasi rantai oleh F. Sanger dan A.R. Coulson di Inggris (Radji, 2011). Metode penghentian rantai dengan dideoksi (dideoxy chain termination) yang dikembangkan oleh Sanger merupakan metode yang paling banyak Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 digunakan. Pada mulanya DNA didenaturasi dan dipisahkan menjadi untai tunggal dengan cara pemanasan. Satu primer oligonukleotida yang telah dilabel radioaktif ditambahkan ke dalam reaksi dan akan menempel pada sekuen pasangannya pada DNA target. DNA polimerase digunakan untuk menyalin DNA untai tunggal. Penambahan dNTP dalam jumlah banyak dan sampai jenuh akan menghasilkan produk ekstensi dengan ujung terlabel radioaktif, tetapi tidak menghasilkan informasi urutan basa. Penambahan sedikit ddNTP ke dalam campuran dNTP akan dapat memberikan informasi urutan basa DNA. Dideoksinukleotida akan terinkorporasi pada ujung 3' untai DNA yang baru disintesis. DNA polimerase tidak dapat menambahkan basa baru pada ddNTP, sehingga inkorporasi ddNTP akan mengakibatkan penghentian sintesis rantai DNA. Penambahan dNTP dan ddNTP dengan rasio yang tepat memungkinkan untuk menghentikan sintesis rantai DNA pada tiap posisi nukleotida (Grompe et al., 1998). Sebagai contohnya, jika ektensi primer dilakukan menggunakan dATP, dTTP, dGTP dan ddCTP, maka polimerase akan mensintesis untai DNA baru sampai harus menggunakan ddCTP. Inkorporasi ddCTP pada titik ini menyebabkan DNA polimerase tidak dapat melanjutkan ekstensi. Dengan demikian, panjang produk hasil ekstensi yang terlabel radioaktif menentukan posisi G pertama yang disalin. Untuk menentukan posisi G yang lainnya, maka reaksi sekuensing yang sebenarnya dilakukan dengan menggunakan campuran dCTP dan ddCTP dengan perbandingan ~200:1. Pada kondisi ini kemungkinan terjadi penghentian rantai DNA adalah ~1:200 yang terjadi ketika terdapat G pada DNA yang disekuensing. Hasilnya akan diperoleh produk ekstensi dengan berbagai panjang, yang kemudian divisualisasi dengan elektroforesis pada gel poliakrilamid. Berdasarkan pada panjang produk, maka tiap fragmen akan menentukan posisi satu G. Untuk menentukan posisi keempat basa, empat reaksi sekuensing dilakukan untuk tiap sampel. Pada tiap reaksi dicampurkan dNTP dan ddNTP yang sesuai dikombinasi dengan 3 dNTP lainnya dalam konsentrasi jenuh. Keempat reaksi sekuensing kemudian dielektroforesis bersebelahan pada gel (poliakrilamid denaturasi), sehingga hasil sekuens DNA dapat langsung dibaca (Grompe et al., 1998). Berdasarkan perbedaan migrasi masing-masing fragmen Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 DNA, maka akan dapat ditentukan urutan basa-basa DNAnya. Hasil sekuensing DNA merupakan urutan basa yang komplementer terhadap urutan basa-basa yang terbaca pada hasil elektroforesis (Radji, 2011). Perkembangan teknologi modern dewasa ini telah memungkinkan dilakukannya automatisasi sekuensing DNA. Perkembangan peralatan yang ada pada saat ini telah memungkinkan dapat membaca hasil sekuensing secara langsung dan sekaligus menyimpan data ke dalam database komputer. Adanya automasisasi dapat mengurangi faktor kesalahan yang sering terjadi dalam pembacaan dan pemulisan urutan DNA secara manual. Mesin sekuensing sekarang telah menggunakan fluoresen sebagai pengganti radioaktif. Senyawa fluoresen dapat diinkorporasikan ke dalam primer sekuensing atau ke dalam nukleotida. Pada sekuensing manual menggunakan elektroforesis gel atau elektroforesis kapiler untuk memisahkan fragmen DNA berdasarkan ukurannya. Pada sekuensing otomatis deteksi fragmen DNA yang berfluoresensi dilakukan dengan bantuan sinar laser dan sinyal diproses oleh komputer (Grompe et al., 1998). 2.7 Ekspresi Protein pada Escherichia coli Pemilihan sistem ekspresi untuk produksi protein rekombinan tergantung pada banyak faktor. Faktor-faktor tersebut diantaranya karakteristik pertumbuhan sel, tingkat ekspresi, modifikasi pascatranslasi, aktivitas biologis protein, dan regulasi produksi protein terapetik (Hodgson, 1993). Dalam pemilihan sistem ekspresi juga perlu mempertimbangkan rincian biaya dalam proses, desain, dan pertimbangan ekonomi lainnya (Makrides, 1996). Inang E. coli. merupakan inang pertama yang digunakan untuk kloning dan ekspresi protein rekombinan. Inang E. coli baik digunakan untuk ekspresi protein intraseluler yang relatif kecil dan tidak memerlukan modifikasi pascatranslasi yang terlalu banyak untuk membuat protein tersebut fungsional. Keuntungan sistem ekspresi pada E. coli diantaranya yaitu mudah untuk dilakukan manipulasi DNA rekombinan serta proses seleksi dan ekspresi yang cepat (Grompe et al., 1998). Tidak setiap gen dapat diekspresikan secara efisien pada organisme E. coli. Hal ini bisa disebabkan oleh sekuen gen yang unik, stabilitas dan efisiensi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 translasi mRNA, kemudahan dalam folding protein, degradasi protein oleh protease sel inang, perbedaan besar dalam penggunaan kodon antara gen asing dan di E. coli, serta potensi toksisitas protein ke inang. Kelemahan E. coli sebagai sistem ekspresi mencakup ketidakmampuannya untuk melakukan banyak modifikasi pascatranslasi pada protein eukariotik, kurangnya mekanisme sekresi untuk efisiensi pelepasan protein ke dalam medium kultur, serta keterbatasan kemampuan untuk memfasilitasi pembentukan ikatan disulfida yang luas. Di sisi lain, banyak protein eukariot yang masih memiliki aktivitas biologis dalam bentuk nonglikosilasi dan oleh karena itu dapat diproduksi dalam E. coli (Fuh et al., 1990). Sistem ekspresi protein rekombinan E. coli merupakan sistem pilihan untuk produksi IFN α mengingat gen ifn α tidak memiliki intron, dan IFN α non glikosilasi telah diketahui bioaktif. Escherichia coli dapat ditumbuhkan mencapai kepadatan sel yang tinggi, dan galur yang digunakan untuk produksi protein rekombinan umumnya adalah aman. Escherichia coli juga dapat menjadi pilihan untuk fermentasi skala besar. Sebagian besar produk farmasi yang telah dipasarkan pada kelompok rekombinan IFN α diproduksi dan dipurifikasi dari E. coli (Rabhi-Essafi, 2007). Sistem ekspresi untuk produksi protein rekombinan dalam E. coli melalui kombinasi antara vektor dan galur E. coli sebagai inang ekspresi (Sørensen dan Mortensen, 2005). Protein dapat diekspresikan dengan bantuan vektor melalui induksi ekspresi menggunakan pemberian reagen seperti isopropyl-1-thio-β-Dgalactopyranoside (IPTG). Reagen IPTG berfungsi melepas represi promotor sehingga mengasilkan ekspresi protein rekombinan dalam jumlah yang tinggi. Dilekatkannya protein tertentu pada molekul protein misalnya β galaktosidase dapat lebih menstabilkan protein, yang mana jika tidak dilekatkan mungkin akan terdegradasi dalam E. coli. Penambahan tag pada protein yang diekspresikan berfungsi untuk memudahkan menemukan dan mengisolasi protein setelah diekspresikan. Tag yang banyak digunakan diantaranya Glutation-S-transferase (GST) yang dapat diisolasi menggunakan kolom glutation atau tag histidin yang diisolasi menggunakan kolom nikel (Grompe et al., 1998). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Ekspresi protein rekombinan pada dasarnya dapat diarahkan ke tiga lokasi yaitu sitoplasma, periplasma atau media kultivasi (Sørensen dan Mortensen, 2005). Protein IFN α yang diekspresikan pada E.coli seringnya dalam bentuk presipitasi agregat tidak larut dalam sitoplasma yang disebut dengan badan inklusi (Swaminathan dan Khanna, 1999; Bedarrain et al., 2001; Srivasta et al., 2005), yang umumnya merupakan protein misfolding dan secara biologis inaktif (Villaverde dan Carrio, 2003). Proses refolding dari badan inklusi kurang diinginkan karena hasil perolehan kembalinya rendah dan memerlukan optimasi kondisi refolding yang berbeda untuk setiap protein target (Middelberg, 2002). Selain itu, proses resolubilisasi tidak dapat secara penuh mengembalikan pelipatan protein dan fungsi yang optimal protein (Fathallah et al., 2009). Beberapa bagian dari protein yang diekspresikan seperti ikatan disulfida (Lilie et al., 1998; Makrides, 1996), modifikasi pascatranslasi (Zhang et al., 1998), hidrofobisitas, muatan (net charge) dan struktur sekunder merupakan faktor-faktor yang menentukan dalam pembentukan badan inklusi (Chiti et al., 2003). Faktor-faktor lain yang juga menentukan pembentukan badan inklusi yaitu terkait dengan lingkungan tempat protein diekspresikan diantaranya pH dan temperatur (Yon, 2002). Reaksi agregasi terjadi pada suhu yang tinggi karena temperatur yang kuat dapat meningkatkan kekuatan interaksi hidrofobik sehingga membuat pelipatan protein salah (Kiefhaber et al., 1991). Tujuan utama ekspresi protein rekombinan pada sistem bakteria adalah mendapatkan akumulasi tingkat tinggi produk terlarut protein rekombinan (Villaverde dan Carrio, 2003). Kelarutan protein target dapat dipengaruhi oleh temperatur induksi dan waktu induksi. Temperatur yang lebih rendah menjadikan level ekspresi lebih lambat sehingga terjadi peningkatkan pelipatan protein target, meskipun dapat menurunkan hasil protein target. Waktu induksi yang lama juga dapat meningkatkan ekspresi, akan tetapi dapat menginduksi proteolisis protein target, terjadinya heterogenitas sampel dan menurunkan hasil yang diperoleh (Fathallah et al., 2009). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 2.8 Analisis Protein Analisis protein dapat dilakukan dengan menggunakan berbagai metode diantaranya yaitu : 2.8.1 Western Blot Protein spesifik dalam campuran protein dapat diidentifikasi dengan teknik Western blotting. Pada metode Western blotting, protein dipisahkan berdasarkan ukuran massanya dengan cara elektroforesis gel poliakrilamida SDS-PAGE (sodium dodecyl sulfate polyacrilamide gel electrophoresis) dengan sistem vertikal (Kuby, 1994). Protein yang akan dielektroforesis sebelumnya ditambah dengan SDS dan kemudian dipanaskan. Sodium dedosil sulfat merupakan suatu detergen anionik yang berguna untuk menyelubungi molekul protein. Penyelubungan protein menyebabkan terganggunya interaksi non-kovalen sehingga molekul protein dalam struktur primer. Merkaptoetanol juga ditambahkan untuk mereduksi ikatan disulfida. Kompleks SDS dengan protein yang terdenaturasi memiliki jumlah muatan negatif yang sebanding dengan ukuran protein (Sudjadi, 2008) Kompleks protein-SDS kemudian dielektroforesis sehingga molekul protein bergerak menuju kutub positif dan terpisah berdasarkan ukuran massanya. Protein yang telah terpisahkan menjadi pita-pita pada gel poliakriamid kemudian ditransfer ke membran nitroselulosa. Proses transfer makromolekul dari gel ke membaran nitriselulosa disebut dengan proses blotting (Radji, 2011). Identifikasi protein dilakukan dengan merendam membran nitroselulosa dalam larutan yang mengandung antibodi primer, kemudian dicuci untuk menghilangkan antibodi primer yang tidak berikatan dengan protein target. Langkah selanjutnya adalah penambahan antibodi sekunder yang hanya melekat pada antibodi sekunder dan tidak melekat pada protein target. Antibodi sekunder telah terkonjugasi dengan suatu enzim yang dapat divisualisasikan dengan penambahan substrat karena dapat mengoksidasi substrat yang tidak berwarna menjadi berwarna (Sudjadi, 2008). 2.8.2 Analisis Densitometri Densitas merupakan kemampuan sebuah material untuk menyerap atau memantulkan sinar. Densitas dapat dibedakan menjadi dua bagian yaitu densitas Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 transmisi yaitu kemampuan lapisan material untuk menyerap sinar yang datang dan densitas refleksi yaitu kemampuan lapisan material untuk memantulkan sinar yang datang. Pengukuran densitas dapat menggunakan sebuat alat ukur yang disebut densitometer. Densitometer merupakan alat yang berfungsi untuk mengukur besarnya densitas atau dengan kata lain merupakan alat yang digunakan untuk mengukur derajat kehitaman atau kepekatan (densitas optis) suatu model atau bahan semi-transparan (Siregar, 2009). Analisis densitometri merupakan salah satu cara untuk konfirmasi hasil elektroforesis. Hasil analisis densitometri terhadap pita-pita pada gel hasil elektroforesis dapat digunakan untuk konfirmasi keberadaan pita, dan untuk kuantifikasi proporsi penyusun protein yang dielektroforesis (Aulanni’am, 2004). Suatu pita menandakan adanya akumulasi protein pada gel hasil elektroforesis diidentifikasi sebagai oleh suatu puncak (peak). Masing-masing puncak memiliki karakteristik ketinggian (height) sebagai intensitas densitograf dan luas daerah di bawah kurva (area) sebagai gambaran kuantitas protein pada pita tersebut. Ketebalan pita pada gel hasil SDS-PAGE dikuantifikasi dalam bentuk luas daerah di bawah kurva pada kurva densitograf (Mustofa et al., 2006). Kuantifikasi intensitas pita Western blot dapat dilakukan dengan sistem digitalisasi automatik menggunakan UN-SCAN-IT. Program UN-SCAN-IT dapat mengkuantifikasi data Western blot dengan menggunakan dua metode. Metode analisis segmental memberikan informasi piksel total dan analisis pita pada setiap segmen, sedangkan analisis lane yang memberikan informasi profil densitas ekstraksi dan analisis pita untuk seluruh jalur (lane). Pada pengukuran pita dilakukan pemindaian sebagai JPEG dalam format 8 bit grayscale pada 600 dpi dan intensitas piksel. Pada pengukuran intensitas piksel dilakukan pengkoreksian terhadap latar (background). Derajat kehitaman atau kepekatan digunakan program UN-SCAN-IT untuk menganalisa pita dan diukur dalam bentuk piksel. Piksel adalah unsur gambar atau representasi sebuah titik terkecil dalam sebuah gambar grafis yang dihitung per inci. Data digital piksel total yang diperoleh dapat digunakan untuk menghitung kepadatan optik relatif pada setiap pita untuk analisis yang lebih lanjut (Rezvani et al., 2009). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 BAB 3 METODE PENELITIAN 3.1 Tempat dan Waktu Penelitian ini dilakukan di laboratorium Terapetik Protein dan Vaksin, Bidang Biologi Molekuler, Pusat Penelitian Bioteknologi, Lembaga Ilmu Pengetahuan Indonesia. Waktu pelaksaan penelitian adalah dari bulan Juli 2011 sampai Juli 2012. 3.2 Bahan dan Alat 3.2.1 Bahan Aquades, aquabides (ddH2O), nuclease free water, GoTaq Green Master Mix (Promega), enzim restriksi Nco I, Xho I, dan Mlu I (Roche), primer IFN_NcoI_F 5' CATGCCATGGCCTGTGATCTGCCTCAAACC 3' (1st BASE), primer IFN_XhoI_R 5' CCGCTCGAGTCATTCCTTACTTCTTAAAC 3' (1st BASE), primer pET_Screen_F 5' TAATACGACTCACTATAGGGGAAT 3' (1st BASE), primer pET_Screen_R 5' TTATTGCTCAGCGGTGGCAGCAGC 3' (1st BASE), STag-18mer-primer 5' GAACGCCAGCACATGGAC 3' (Macrogen), T7terminator-primer 5' GCTAGTTATTGCTCAGCGG 3' (Macrogen), agarosa (Fermentas), marker DNA 1 kilo basa (Fermentas), loading buffer, etidium bromida (EtBr), enzim T4 DNA ligase (NEB), DNA ligation buffer (NEB), Agarose Gel DNA Extrction Kit (Roche), metanol, etanol, kalsium klorida (CaCl2), ampisilin (Sigma), ekstrak ragi (Difco), tripton (Difco), agar bakto (Difco), isopropyl-1-thio-β-D-galactopyranoside /IPTG (Roche), natriun klorida (NaCl), sodium deodesil sulfat (SDS), akrilamid, ammonium persulfat (APS), separating gel buffer, stacking gel buffer, tetrametil metilen diamin (TEMED), bufer elektroforesis, nitro blue tetrazolium/NBT (Promega), 5-bromo-4chloro-3indolyl-phosphate/BCIP (Promega), Anti-α-Interferon Mouse mAb (MMHA-2) (Calbiochem), Anti-mouse IgG (H+L) AP conjugate (Promega), etanol, metanol, gen sintetik human ifn α2a dalam vektor pcDNA3.1+:IFN-Gene (DNA 2.0 Inc., Menlo Park), vektor pET-32b(+) (Novagen), E. coli galur DH5α (Invitrogen), E. coli galur BL21(DE3) (Novagen). 25 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 3.2.2 Alat Alat PCR (Biometra), alat elektroforeis DNA (Advance), alat elektroforeis protein (Bio-rad), Ultraviolet (UV) Transiluminator, autoklaf, inkubator shaker, penangas air, laminar, lemari pendingin -20⁰C, lemari pendingin 4⁰C, oven, mikropipet, vortex, sentrifus, cawan petri, labu erlenmeyer, tabung reaksi, tip, tabung PCR, tabung sentrifus. 3.3 Tahap Kerja Metode yang digunakan dalam penelitian ini secara garis besar berdasarkan metode pada buku panduan pET Sistem Manual (2006). 3.3.1 Penyiapan DNA insert Penyiapan DNA insert dilakukan dengan metode PCR menggunakan cetakan gen human ifn α2a dalam vektor pcDNA3.1+IFN-Gene. Proses PCR dilakukan dengan menggunakan sepasang primer yaitu IFN_Nco_I F 5' CATGCCATGGCCTGTGATCTGCCTCAAACC 3' dan IFN_Xho_I R 5' CCGCTCGAGTCATTCCTTACTTCTTAAAC 3' yang telah ditambahkan situs pemotongan Nco I dan Xho I yang dicetak miring dan digaris bawahi. Proses PCR dimulai dengan dicampurkannya semua komponen yang dibutuhkan dalam reaksi ke dalam tabung PCR, yaitu ddH2O (6,5 µl), DNA (2 µl), primer IFN_NcoI_F (2 µl), primer IFN_XhoI_R (2 µl), dan GoTaq Green Master Mix (12,5 µl) sehingga volume totalnya 25µl. Reaksi PCR dilakukan dengan siklus sebagai berikut: denaturasi awal pada suhu 94⁰C selama 5 menit, denaturasi 94⁰C selama 1 menit, penempelan 55⁰C selama 1 menit, dan polimerisasi 72⁰C selama 2 menit (tiga tahap ini dilakukan sebanyak 30 siklus) serta polimerisasi akhir pada suhu 72⁰C selama 6 menit (pET Sistem Manual, 2006). Produk hasil PCR selanjutnya dilakukan pengecekan dengan metode elektroforesis pada medium agarosa 1% dengan sampel hasil PCR (5 µl), ddH2O (5 µl), loading buffer (2 µl) dengan tegangan 100 volt selama kurang lebih 30 menit. Elektroforesis dilakukan sampai loading buffer mencapai dua pertiga dari panjang gel. Gel agarosa kemudian diwarnai dengan larutan EtBr dan diamati diatas sinar UV. Hasil PCR yang benar kemudian dilakukan purifikasi dengan menggunakan 2 metode yang berbeda yaitu gel (Agarose Gel DNA Extraction Kit) dan metode fenol kloroform. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Tahapan purifikasi hasil PCR menggunakan Agarose Gel DNA Extraction Kit dilakukan berdasarkan manual (Roche) adalah: 1. Hasil PCR 60 µl dielektroforesis pada gel agarosa 1%, kemudian divisualisasikan dengan etidium bromida (EtBr). 2. Gel agarosa dilihat pada UV transiluminator, selanjutnya pita fragmen hasil PCR diambil dengan dipotong menggunakan scalpel kemudian dimasukkan ke tabung 2 ml. 3. Berat agarosa lalu ditimbang dan ditambahkan 300 µl agarose solubilisation buffer (vial 2) per 100 mg gel agarosa. 4. Sebanyak 10 µl suspensi silika (vial 1) ditambahkan ke campuran untuk mengikat DNA. 5. Campuran diinkubasi selama 10 menit pada suhu 57⁰C dan divortex setiap 2 menit. 6. Campuran selanjutnya disentrifus 12.000 rpm selama 1 menit dan supernatannya dibuang dengan pemipetan. 7. Pelet ditambahkan 500 µl nucleic acid binding buffer (vial 3) dan divortex. 8. Campuran lalu disentrifus 12.000 rpm selama 1 menit dan supernatannya dibuang dengan pemipetan. 9. Pelet dicuci dengan 500 µl washing buffer (vial 4). 10. Campuran disentrifus 12.000 rpm selama 1 menit dan supernatannya dibuang dengan pemipetan. 11. Pelet dicuci lagi dengan 500 µl washing buffer (vial 4). 12. Campuran disentrifus 12.000 rpm selama 1 menit dan supernatannya dibuang dengan pemipetan. 13. Proses pengeringan dilakukan pada suhu kamar selama 10-15 menit. 14. Pelet dilarutkan dengan 20 µl nuclease free water dan diinkubasi 10 menit pada suhu 15-25⁰C untuk DNA insert, sedangkan untuk vektor diinkubasi 10 menit pada suhu 56-60⁰C dengan divortex setiap 2-3 menit. 15. Campuran disentrifus 12.000 rpm selama 1 menit dan supernatan diambil dan dipindahkan ke tabung baru. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Tahapan proses purifikasi hasil PCR dengan metode fenol kloroform yaitu: 1. Hasil PCR 60 µl ditambahkan ddH2O 90 µl kemudian ditambahkan 150 µl fenol kloroform dan divortex. 2. Campuran disentrifus 12.000 rpm selama 5 menit pada suhu 4⁰C. 3. Bagian atasnya diambil dan dipindahkan ke tabung baru kemudian ditambahkan etanol absolut (2,5 kali volume) dan disimpan semalam. 4. Campuran disentrifus 12.000 rpm selama 30 menit pada suhu 4⁰C dan supernatan dibuang dengan pemipetan. 5. Pelet dicuci dengan 1 ml etanol 70%, disentrifus 12.000 rpm selama 5 menit pada suhu 40C kemudian supernatan dibuang dengan pemipetan. 7. Pelet dikeringkan pada suhu kamar kurang lebih 30 menit selanjutnya dilarutkan dengan 20 µl nuclease free water. Hasil PCR yang telah dipurifikasi kemudian dipotong menggunakan enzim restriksi Nco I dan Xho I dan dilanjutkan purifikasi dengan fenol kloroform. Proses pemotongan dimulai dengan dicampurkannya semua komponen ke dalam tabung, yaitu ddH2O (10,5 µl), bufer H (2,5 µl), hasil PCR yang telah dipurifikasi (10 µl), enzim Nco I (1 µl), enzim Xho I (1 µl) dengan volume total 25 µl dan diinkubasi pada suhu 37⁰C dalam penangas air (waterbath) selama 1 jam. Hasil dari proses pemotongan kemudian dilakukan pengecekan dengan metode elektroforesis pada medium agarosa 1% dengan sampel (2 µl), ddH2O (8 µl), loading buffer (2 µl) dan diamati diatas sinar UV. Tahapan proses purifikasi DNA insert yang telah dipotong dengan metode fenol kloroform yaitu: 1. DNA insert hasil pemotongan 23 µl ditambahkan ddH2O 100 µl kemudian ditambahkan 130 µl fenol kloroform dan divortex. 2. Campuran disentrifus 12.000 rpm selama 5 menit pada suhu 4⁰C. 3. Bagian atasnya diambil dan dipindahkan ke tabung baru kemudian ditambahkan etanol absolut (2,5 kali volume) dan disimpan semalam. 4. Campuran disentrifus 12.000 rpm selama 30 menit pada suhu 4⁰C, kemudian supernatan dibuang dengan pemipetan. 5. Pelet dicuci dengan 1 ml etanol 70% dan disentrifus 12.000 rpm selama 5 menit pada suhu 4⁰C kemudian supernatan dibuang dengan pemipetan. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 7. Pelet dikeringkan pada suhu kamar kurang lebih 30 menit selanjutnya dilarutkan dengan 20 µl nuclease free water. 3.3.2 Penyiapan Vektor pET-32b(+) Vektor pET-32b(+) dengan konsentrasi 300 ng/µl dilakukan pemotongan dengan enzim restriksi Nco I dan Xho I pada suhu 37⁰C selama 1 jam. Proses pemotongan vektor pET-32b(+) dimulai dengan dicampurkannya semua komponen ke dalam tabung, yaitu ddH2O (16,5 µl), bufer H (2,5 µl), vektor pET32b(+) (4 µl), enzim Nco I (1 µl), enzim Xho I (1 µl) dengan volume total 25 µl dan selanjutnya diinkubasi pada suhu 37⁰C dalam penangas air selama 1 jam. Hasil proses pemotongan kemudian dilakukan pengecekan dengan metode elektroforesis pada medium agarosa 1% dengan sampel (3 µl), ddH2O (7 µl), loading buffer (2 µl) dan dilakukan pengamatan diatas sinar UV. Vektor pET32b(+) yang telah dipotong kemudian dipurifikasi menggunakan fenol kloroform yang mana metodenya serupa dengan purifikasi fenol kloroform pada penyiapan DNA insert. Vektor pET-32b(+) yang telah linear selanjutnya dilakukan tahapan ligasi dengan DNA insert (gen human ifn α2a) yang telah dipersiapkan. Reaksi ligasi terdiri dari nuclease free water (12 µl), bufer T4 DNA ligase (2 µl), vektor pET32b(+) (2 µl), DNA insert (3 µl), enzim T4 DNA ligase (1 µl) dengan volume total 20 µl dan diinkubasi pada suhu 4⁰C semalam. Hasil dari proses ligasi diharapakan dapat diperoleh vektor rekombinan pET-32b(+)-IFN α2a yang sirkuler. 3.3.3 Pembuatan Sel Kompeten E. coli Proses pembuatan sel kompeten E. coli adalah sebagai berikut : 1. Stok beku 20 µl dari koloni tunggal E. coli diinokulasikan pada 4 ml LB cair dan ditumbuhkan pada suhu 37⁰C dikocok 200 rpm semalam. 2. Sebanyak 800 µl kultur semalam diinokulasikan pada 100 ml LB cair dan ditumbuhkan pada suhu 37⁰C dikocok 250 rpm selama 3 jam. 3. Kultur didinginkan di es selama 5-10 menit kemudian dituang ke 3 tabung sentrifus masing-masing 35 ml. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 4. Kultur disentrifus 6000 rpm pada suhu 4⁰C selama 5 menit dan supernatan dibuang. 5. Pelet diresuspen dalam 4 ml CaCl2 60 mM dingin untuk masing-masing tabung, dikocok-kocok supaya homogen. 6. Campuran didiamkan di es 30 menit lalu disentrifus 6000 rpm pada suhu 4⁰C selama 5 menit dan supernatannya dibuang. 7. Pelet diresuspen dalam 4 ml CaCl2 60 mM dingin untuk masing-masing tabung, dikocok-kocok supaya homogenkan. 8. Campuran didiamkan di es selama 30 menit lalu disentrifus 6000 rpm pada suhu 4⁰C selama 5 menit dan supernatannya dibuang. 9. Pelet diresuspen dengan 1 ml CaCl2 60 mM dingin untuk masing-masing tabung, kemudian digabung menjadi satu dan ditambah 1 ml CaCl2 60 mM sehingga volume akhir 4 ml. Sel kompeten dapat langsung digunakan atau disimpan semalam dalam lemari es 4⁰C untuk meningkatkan kompetensi. 3.3.4 Kloning Gen human ifn α2a Hasil ligasi kemudian ditransformasikan pada sel kompeten E. coli galur DH5α sebagai inang kloning. Proses transformasi dilakukan dengan menggunakan metode heat shock pada suhu 42⁰C selama 90 detik (Radji, 2011). Adapun proses dari transformasi adalah sebagai berikut: 1. Sel kompeten 100 µl ditambahkan hasil ligasi 5-10 µl. 2. Campuran diinkubasi di es selama 30 menit. 3. Campuran dilakukan heat shock pada penangas air suhu 42⁰C selama 90 detik. 4. Campuran didinginkan di es selama 1 menit. 5. Campuran ditambahkan 400 µl LB cair steril. 6. Campuran diinkubasi pada suhu 37⁰C dengan dikocok selama 45 menit. 7. Campuran diinokulasi ke media LB agar yang telah ditambah ampisilin dengan metode sebar 20 µl, 50 µl, 80 µl, dan 350 µl (suspensi sel 350 µl disentrifus dulu dan buang supernantan sisakan 80 µl). 8. Tahap inkubasi dilakukan pada inkubator 37⁰C semalan dan selanjutnya diisolasi koloni tunggal yang tumbuh. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Hasil transformasi ditumbuhkan pada media LB padat yang ditambah ampisilin (pET Sistem Manual, 2006). Sebagai kontrol negatif digunakan suspensi sel kompeten yang ditumbuhkan pada media LB padat yang ditambah ampisilin. Klon transforman E. coli diseleksi dengan menggunakan marker antibiotik ampisilin. Hanya klon E. coli yang mengandung vektor rekombinan pET-32b(+)IFN α2a yang dapat tumbuh pada media LB padat yang ditambah ampisilin. 3.3.5 Skrining dan Verifikasi Klon Transforman di E. coli DH5α Verifikasi masuknya gen DNA insert pada vektor rekombinan dilakukan dengan beberapa metode yaitu PCR colony screening, isolasi vektor rekombinan pET-32b(+)-IFN α2a, analisis restriksi dan juga sekuensing (pET Sistem Manual, 2006). Proses PCR colony screening dilakukan dengan menggunakan primer pET_Screen_F 5' TAATACGACTCACTATAGGGGAAT 3' dan pET_Screen_R 5' TTATTGCTCAGCGGTGGCAGCAGC 3'. Metode PCR colony screening dilakukan dengan tahapan sebagai berikut: 1. Koloni yang tumbuh berdiameter lebih dari 1 mm diambil dengan pipet tip. 2. Pipet tip dicelupkan ke tabung 0,5 ml yang telah diisi 50 µl aquades. 3. Tabung dipanaskan di heat block pada suhu 99⁰C selama 5 menit untuk melisiskan sel dan merusak DNAse. 4. Campuran disentrifus pada 12.000 rpm selama 1 menit untuk mengendapkan debris sel. 5. Campuran diambil 10 µl supernatan dan dilakukan prosesPCR. 6. Reaksi PCR terdiri atas supernatan (10µl), primer pET_Screen_F (2 µl), primer pET_Screen_R (2 µl), GoTag Green Master Mix (12,5 µl) dengan volume total 26,5 µl. 7. Setelah proses PCR selesai dilakukan pengecekan dengan elektrofosesis gel agarosa dan divisualisasikan menggunakan EtBr. 3.3.6 Kultivasi dan Isolasi Vektor Rekombinan Metode isolasi vektor rekombinan pET-32b(+)-IFN α2a dilakukan dengan tahapan sebagai berikut: 1. Media LB cair dibuat dan disterilisasi dengan autoklaf. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Contoh pembuatan media LB cair 10 ml yaitu: bakto tripton 1% = 0,2 gram, ekstrak ragi 0,5% = 0,1 gram, NaCl 1% = 0,2 gram (pET Sistem Manual, 2006). Setelah steril dan sebelum digunakan perlu ditambahakan 10 µl ampisilin 5%. 2. Koloni tunggal diinokulasikan ke dalam tabung yang berisi media 3 ml yang telah ditambah ampisilin dan diinkubasi semalam pada suhu 37⁰C dengan dikocok 200 rpm. 3. Kultur diambil dan dimasukkan ke tabung steril 1,5 ml kemudian disentrifus 12.000 rpm selama 1 menit. Selain itu juga dilakukan pembuatan kultur stok yang disimpan pada suhu -20⁰C dengan ditambah gliserol. 4. Supernatan dibuang dengan pemipetan kemudian pelet divortex. 5. Pelet ditambahkan 150 µl larutan 1 diaduk sampai larut dan divortex kemudian diinkubasi di es selama 10 menit. Larutan 1 terdiri dari 50 mM glukosa, 25 mM tris pH 8, 10 mM EDTA dan ddH2O. 6. Campuran ditambahkan 180 µl larutan 2 diaduk perlahan-lahan sampai ada lendir kemudian ditaruh dies selama 10 menit. Larutan 2 terdiri dari 0,2 M NaOH, SDS 1% dan ddH2O. 7. Campuran ditambahkan 135 µl larutan 3 dingin (sodium asetat 3 M pH 5,7) diaduk perlahan-lahan dan dibolak-balik. 8. Campuran disentrifus 12.000 rpm pada suhu 4⁰C selama 5 menit. Pada pelet berisi DNA genom, sedangkan pada supernatan berisi DNA vektor. 9. Supernatan diambil dan dipindahkan ke tabung baru, ditambahkan 400 ml fenol kloroform (1:1) kemudian divortex. 10. Campuran disentrifus 12.000 rpm selama 5 menit pada suhu 4⁰C. 11. Bagian atas diambil dan pindahkan ke tabung baru kemudian ditambahkan etanol absolut (2,5 kali volume) dan disimpan semalam. 12. Campuran disentrifus 12.000 rpm selama 30 menit pada suhu 4⁰C kemudian supernatan dibuang dengan dipipet. 13. Pelet dicuci dengan ditambahkan 1 ml etanol 70%. 14. Campuran disentrifus 12.000 rpm selama 5 menit pada suhu 4⁰C kemudian supernatan dibuang dengan aspirasi. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 15. Pelet dikeringkan pada suhu kamar kurang lebih 30 menit dan dilarutkan dengan 20 ml ddH2O yang ditambah 1 µl RNAse 1:5. 16. Campuran diinkubasi selama 20-60 menit pada 37⁰C, lalu disimpan di -20⁰C. Vektor rekombinan pET-32b(+)-IFN α2a yang telah diisolasi selanjutnya dilakukan analisis restriksi dan disekuen untuk konfirmasi adanya DNA insert dan peletakan kodon yang tepat. Vektor rekombinan pET-32b(+)-IFN α2a dilakukan pemotongan kembali menggunakan enzim restriksi Nco I dan Xho I. Komponen analisis restriksi terdiri dari ddH2O (12,5 µl), bufer H (2,5 µl), vektor rekombinan (8 µl), enzim Nco I (1 µl), enzim Xho I (1 µl) dengan total volume 25 µl dan diinkubasi 37⁰C pada penangas air selama 1 jam. Hasil pemotongan kemudian dielektroforesis pada gel agarosa 1%. Vektor rekombinan pET-32b(+)-IFNα2a yang telah benar selanjutnya dilakukan perbanyakan dengan dikultivasi. Isolasi dan purifikasi purifikasi vektor rekombinan dilakukan dengan Qiagen Plasmid Maxi Kit serta diukur konsentrasinya menggunakan alat Nanophotometer. Prosedur Qiagen Plasmid Maxi Kit adalah sebagai berikut: 1. Kultur stok gliserol 35 µl ditumbuhkan ke dalam LB cair 5 ml yang telah ditambah ampisilin dan diinkubasi pada 37⁰C dikocok 200 rpm semalam. 2. Kultur diambil 4 ml dan diinokulasikan ke LB cair 200 ml yang telah ditambah ampisilin. 3. Kultur diinkubasi pada 37⁰C dikocok 200 rpm selama 6 jam. 4. Kloramfenikol 30 mg ditambahkan dan diinkubasi pada 37⁰C dikocok 200 rpm semalam. 5. Pemanenan sel dilakukan dengan disentrifus pada 4000 rpm 4⁰C selama 15 menit dan supernatannya dibuang. 6. Pelet diresuspensi dengan 10 ml bufer P1 yang mana sudah mengandung RNase. 7. Bufer P2 10 ml ditambahkan dengan dibolak-balik 5 kali dan campuran selanjutnya diinkubasi selama 5 menit pada suhu kamar. 8. Bufer P3 10 ml dingin ditambahkan dan dihomogenkan dengan dibolak-balik 5 kali dan campuran selanjutnya diinkubasi selama 20 menit pada suhu kamar. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 9. Campuran disentrifus pada 13.000 rpm 4⁰C selama 30 menit dan supernatannya diambil. 10. Qiagen tip 500 diekuilibrasi dengan ditambahkan 10 ml bufer QBT dan bufer dibiarkan keluar menetes karena gaya gravitasi sampai kolom menjadi kosong. 11. Supernatan dimasukkan ke Qiagen tip 500. 12. Qiagen tip 500 dilakukan pencucian 2 kali dengan 30 ml bufer QC. 13. DNA plasmid dielusi dengan ditambahkan 15 ml buffer QF. 14. Hasil elusi dipresipitasi dengan ditambahkan 10,5 ml isopropanol. 15. Campuran disentrifus pada 13.000 rpm 4⁰C selama 30 menit dan supernatannya dibuang. 16. Pelet dicuci dengan ditambahkan 5 ml etanol 70% dan disentrifus pada 13.000 rpm 4⁰C selama 10 menit dan supernatannya dibuang. 17. Pelet dikeringkan pada suhu ruang selama 10 menit kemudian pelet dilarutkan 2 ml nuclease free water. Karakterisasi selanjutnya yaitu dengan analisis urutan nukleotida menggunakan metode sekuensing. Pembacaan dilakukan dua arah yaitu menggunakan STag-18mer-primer dan T7terminator-primer. Penentuan urutan nukleotida dilakukan di Korea (Macrogen). 3.3.7 Transformasi Vektor Rekombinan ke E.coli BL21(DE3) Vektor rekombinan pET-32b(+)-IFNα2a yang benar selanjutnya ditransformasikan ke sel kompeten E. coli BL21(DE3) untuk mengekspresikan protein rekombinan human IFN α2a. Metode transformasi yang digunakan yaitu heat shock pada suhu 42⁰C selama 90 detik. Tahapan transformasi vektor rekombinan pET-32b(+)-IFNα2a ke sel kompeten E. coli BL21(DE3) dilakukan seperti metode transformasi hasil ligasi ke sel kompeten E. coli DH5α. Hasil transformasi kemudian ditumbuhkan pada media LB padat yang ditambah ampisilin (pET Sistem Manual, 2006). 3.3.8 Skrining dan Verifikasi Klon Transforman di E. coli BL21(DE3) Klon transforman E. coli BL21(DE3) yang diperoleh diseleksi dengan media selektif LB padat yang mengandung antibiotik ampisilin. Verifikasi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 masuknya vektor rekombinan pET-32b(+)-IFN α2a pada inang ekspresi E. coli BL21(DE3) dilakukan menggunakan PCR colony screening dan indentifikasi protein terfusi. Proses PCR colony screening klon transforman E. coli BL21(DE3) dilakukan seperti PCR colony screening pada klon transforman E. coli DH5α. Hasil proses PCR colony screening kemudian dilakukan pengecekan dengan metode elektroforesis pada medium agarosa 1% dengan sampel (5 µl), ddH2O (5 µl), loading buffer (2 µl) dilakukan pewarnaan dengan EtBr dan diamati diatas sinar UV. 3.3.9 Ekspresi, Isolasi dan Karakterisasi Protein Rekombinan Human IFN α2a Klon transforman E. coli BL21(DE3) yang terpilih selanjutnya dilakukan uji ekspresi protein rekombinan pada skala kecil menggunakan medium produksi yaitu media LB cair 10 ml yang ditambah 50 µg/ml ampisilin. Inkubasi dilakukan dengan variasi suhu pada 28⁰C, 30⁰C dan 37⁰C. Kultur sel bakteri pada OD600 sekitar 0,5 diinduksi menggunakan IPTG konsentrasi akhir 1 mM. Adapun protokol induksi yang digunakan adalah sebagai berikut: 1. Kultur stok gliserol 20 µl ditumbuhkan pada media LB cair 3 ml yang telah ditambah ampisilin dan diinkubasi pada 37⁰C dikocok 200 rpm semalam. 2. Kultur semalam diambil 0,5 ml dan dimasukkan ke LB cair 10 ml yang telah ditambah ampisilin. 3. Kultur ditumbuhkan dengan dikocok 250 rpm pada suhu 37⁰C mencapai OD600: 0,5 sekitar 2 jam. 4. Kultur diambil 0,5 ml dimasukkan ke tabung 1,5 ml dan disentrifus pada 8000 rpm 4⁰C selama 10 menit, selanjutnya dipisahkan antara supernatan dan pelet sebagai sampel 0 jam sebelum induksi lalu disimpan pada -20⁰C. 5. Kultur diinduksi dengan penambahan IPTG konsentrasi akhir 1 mM dan dilanjutkan inkubasi pada variasi suhu 28⁰C, 30⁰C, dan 37⁰C dengan dikocok 250 rpm. 6. Pengambilan sampel 0,5 ml dilakukan pada waktu 1, 2, 3, 4, 5, 6, 7 jam dan semalam, selanjutnya disentrifus dan dipisahkan antara supernatan dan pelet Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 lalu disimpan pada -20⁰C. Pelet selanjutnya diresuspensi dengan menggunakan 100 µl buffer PBS 1x. Isolasi protein rekombinan human IFN α2a dari pelet dilakukan dengan metode sebagai berikut: 1. Kultur stok gliserol 20 µl ditumbuhkan pada media LB cair 3 ml yang telah ditambah ampisilin dan diinkubasi pada 37⁰C dikocok 200 rpm semalam. 2. Kultur semalam diambil 1,25 ml dan dimasukkan ke LB cair 25 ml yang telah ditambah ampisilin. 3. Kultur ditumbuhkan dengan dikocok 250 rpm pada suhu 37⁰C mencapai OD600: 0,5 sekitar 2 jam. 4. Kultur diinduksi dengan penambahan IPTG konsentrasi akhir 1 mM dan dilanjutkan inkubasi pada suhu 37⁰C dengan dikocok 250 rpm. 5. Pemanenan sel dilakukan setelah 5 jam inkubasi dengan disentrifugasi 12.000 rpm pada suhu 4⁰C selama 10 menit untuk memisahkan pelet dan supernatan. 6. Pelet diresuspensi dengan 2,5 ml bufer 50 mM tris HCl pH 8 yang ditambah 1 mM EDTA dan 1 mM PMSF. 7. Campuran disonikasi sebanyak 10 kali selama 30 detik di es dengan selang waktu 30 detik di es. 8. Campuran disentrifugasi pada 12.000 rpm suhu 4⁰C selama 10 menit untuk memisahkan antara supernatan (protein terlarut di sitoplasma) dan pelet. 9. Pelet dicuci dengan bufer 50 mM tris HCl yang ditambah 1% triton X-100 (750 µl bufer untuk 0,1 gram pelet). 10. Campuran diikubasi 5 menit pada suhu kamar dan selanjutnya disentrifus 12.000 rpm pada suhu 4⁰C selama 5 menit. 11. Tahap pencucian dilakukan sebanyak 2 kali. 12. Pelet disolubilisasi dengan bufer 50 mM tris HCl yang ditambah 6 M guanidin HCl dan 800 mM merkaptoetanol (750 µl bufer untuk 0,1 gram pelet). 13. Campurn diinkubasi selama 30 menit pada suhu kamar dan selanjutnya disentrifus pada 12.000 rpm pada suhu 4⁰C selama 15 menit. 14. Supernatan dipisahkan dari pelet selanjutnya di cek menggunakan SDS PAGE dengan pewarnaan CBB atau Western blot dan penyimpanan dilakukan pada Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 suhu -20⁰C. Supernatan merupakan protein yang tidak terlarut yaitu sebagai badan inklusi di dalam sitoplasma. Karakterisasi protein rekombinan human IFN α2a yang diperoleh dilakukan dengan metode elektroforesis SDS-PAGE yang dilanjutkan dengan pewarnaan CBB atau Western blot. Tahapan metode elektroforesis SDS-PAGE adalah sebagai berikut: 1. Pembuatan gel poliakrilamid yang terdiri atas separating gel dan stacking gel. Larutan separating gel 12%: 1,5 M tris HCl pH 8,8 (2,5 ml); 10% SDS (100 µl); 40% akrilamid (3 ml); aquades (4,4 ml); APS 10% (100 µl); TEMED (6 µl). Larutan separating gel dimasukkan dalam cetakan dan dibiarkan selama 30 menit supaya berpolimerisasi sehingga terbentuk gel. Larutan stacking gel: 0,5 M tris HCl pH 6,8 (1,25 ml); 10% SDS (100 µl); 40% akrilamid (900 µl); aquades (7 ml); APS 10% (100 µl); TEMED (15 µl). Larutan stacking gel dimasukkan ke dalam cetakan pada posisi diatas separating gel kemudian sisir dimasukkan pada cetakan dan gel akan terbentuk setelah 30 menit. 2. Penyiapan sampel untuk dielektroforesis. Pada supernatan dan pelet masing-masing diambil 10 µl dimasukkan ke dalam tabung dan ditambahkan 10 µl loading buffer RSB 2x (1:1) lalu dipanaskan pada 95⁰C selama 4 menit. Marker 5 µl ditambah 5 µl loading bufer RSB 2x. 3. Sampel dimasukkan kedalam sumuran dan dielektroforesis dalam bufer elektroforesis pada tegangan 90 volt selama 90 menit. Bufer elektroforesis untuk 1 liter pH 8,3: tris 15,5 g, glisin 72 g, SDS 5 g, lalu ditambah aquades sampai 1 liter. Gel poliakrilamid yang telah selesai dielektroforesis SDS PAGE kemudian dikarakterisasi dengan pewarnaan commasie brilliant blue (CBB) atau menggunakan Western blot. Proses pewarnaan CBB adalah sebagai berikut: 1. Gel SDS-PAGE diinkubasi dengan stain (40% metanol: 20 ml; 7% asam asetat glasial: 3,5 ml; ddH2O: 26,5 ml; CBB stain: 6 ml) dan dikocok 3 jam. 2. Gel dicuci dengan destain I (40% metanol: 20 ml; 7% asam asetat glasial: 3,5 ml; ddH2O: 26,5 ml) dan dikocok semalam. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 3. Gel dicuci dengan destain II (5% methanol: 2,5 ml; 7% asam asetat glasial: 3,5 ml; ddH2O 44 ml) dikocok 1 jam. Tahapan metode Western blot adalah sebagai berikut: 1. Protein sebelumnya telah dipisahkan dengan metode SDS PAGE. 2. Protein selanjutnya ditransfer ke membran nitriselulosa dengan cara dielektroforesis dalam bufer transfer pada tegangan 90 volt selama 90 menit. Bufer transfer untuk 1 liter: tris 25 mM 3,03 g, glisin 192 mM 14,4 g, 20% metanol 200 ml, ditambah aquades sampai 1 liter. 3. Membran diambil kemudian diinkubasi dengan 10% susu bebas lemak dalam bufer TBS 1x dengan dikocok selama 1 jam. Bufer TBS (Tris Buffer Saline) untuk 1 liter dengan pH 7,6: tris 2,42 g, NaCl 8g, HCl 1N 3,8 g, ditambah aquades sampai 1 liter. 4. Pencucian dilakukan 3 kali menggunakan 0,1% Tween 20 dalam TBS 1x selama 15 menit, 5 menit dan 5 menit dengan dikocok. 5. Larutan antibodi primer ditambahkan dan diinkubasi 1 jam dengan dikocok. Larutan antibodi primer: 10 µl anti-α-Interferon Mouse mAb dilarutkan pada 10 ml 10% susu bebas lemak dalam bufer TBS 1x. 6. Pencucian dilakukan 3 kali menggunakan 0,1% Tween 20 dalam TBS 1x selama 15 menit, 5 menit dan 5 menit dengan dikocok. 7. Larutan antibodi sekunder ditambahkan dan diinkubasi 1 jam dengan dikocok. Larutan antibodi sekunder: 3 µl anti-mouse IgG (H+L) AP conjugate dilarutkan pada 10 ml 10% susu bebas lemak dalam bufer TBS 1x. 8. Pencucian dilakukan 3 kali menggunakan 0,1% Tween 20 dalam TBS 1x selama 15 menit, 5 menit dan 5 menit dengan dikocok. 9. Hasil analisis diketahui dengan munculnya pita pada membran nitriselulosa setelah pemberian substrat NBT-BCIP (bufer alkaline phosphatase 5 ml; NBT 33 µl; dan BCIP 16,5 µl). Hasil SDS PAGE yang dilanjutkan Western blot difoto yang selanjutnya dilakukan analisis densitometri dengan sistem digitalisasi menggunakan UN-SCAN-IT Gel versi 6.1. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 automatik BAB 4 HASIL DAN PEMBAHASAN 4.1 Penyiapan DNA insert Penyiapan DNA insert dilakukan dengan metode PCR menggunakan cetakan gen sintetik human ifn α2a dalam vektor pcDNA3.1:IFN-Gene. Metode PCR dipilih karena mampu diperoleh sejumlah besar DNA dengan urutan basa tertentu dalam waktu yang singkat (Grompe et al., 1998). Primer spesifik dibuat berdasarkan sekuen gen human ifn α2a pada GenBank DI084466.1. Sepasang primer digunakan untuk tahap amplifikasi gen human ifn α2a yaitu IFN_NcoI_F dan IFN_XhoI_R yang telah ditambahkan situs restriksi Nco I dan Xho I. Penambahan situs restriksi bertujuan untuk menempelkan situs restriksi pada gen human ifn α2a yang selanjutnya digunakan untuk proses ligasi ke vektor pET32b(+). Dua jenis enzim endonuklease restriksi digunakan untuk menghindari keterbalikan arah transkripsi atau translasi gen yang disisipkan ke dalam plasmid (Radji, 2011). Hasil amplifikasi gen human ifn α2a selanjutnya dilakukan pengecekan dengan dielektroforesis pada gel agarosa 1%. 1 2 3 4 750 pb 500 pb 5 6 7 510 pb Gambar 4.1 Elektroforesis Hasil PCR untuk Penyiapan DNA Insert. Keterangan: (1) Marker; (2-7) Produk PCR. Metode PCR yang digunakan pada penelitian ini dapat mengamplifikasi gen human ifn α2a dengan baik. Amplifikasi dengan primer IFN_NcoI_F dan IFN_XhoI_R menghasilkan produk yang berukuran 510 pb (Gambar 4.1). Berdasarkan pita DNA pada hasil elektroforesis dapat diketahui bahwa primer IFN_NcoI_F dan IFN_XhoI_R secara spesifik mengamplifikasi gen target yang 39 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 diindikasikan dengan terdapatnya pita tunggal. Adanya perbedaan ketebalan pita yang tampak pada hasil elektroforesis kemungkinan dikarenakan oleh proses pemipetan dalam pengambilan sampel pada tabung kurang diaduk homogen, meskipun volume yang diambil sama akan tetapi konsentrasinya dapat berbeda. Hasil amplifikasi dengan metode PCR selanjutanya dilakukan purifikasi sebelum digunakan untuk tahapan kloning. Purifikasi bertujuan untuk menghilangkan sisa-sisa primer, garam, enzim, dan pengotor-pengotor lain yang dapat mengganggu proses kloning. Purifikasi produk PCR untuk penyiapan DNA insert pada penelitian ini dilakukan dengan menggunakan dua metode yang berbeda yaitu metode Agarose Gel DNA Extraction Kit (Roche) dan metode purifikasi fenol kloroform. Hasil purifikasi produk PCR selanjutnya dilakukan pengecekan dengan dielektroforesis pada gel agarosa 1%. 1 510 pb 2 3 4 5 6 7 750 pb 500 pb 510 pb Gambar 4.2 Elektroforesis Hasil Purifikasi Produk PCR dan Hasil Pemotongan Ganda dengan Enzim Nco I dan Xho I. Keterangan: (1) Produk PCR; (2) Produk PCR dipurifikasi Agarose gel DNA extraction kit; (3) Produk PCR dipurifikasi Fenol kloroform; (4 dan 6) Kosong; (5) Marker (7) Hasil purifikasi Agarose gel DNA extraction kit dipotong enzim Nco I dan Xho I. Pita hasil purifikasi menggunakan metode Agarose Gel DNA Extraction Kit dan metode fenol kloroform sama-sama mempunyai ukuran 510 pb (Gambar 4.2 nomor 2 dan 3). Akan tetapi hasil elektroforesis purifikasi dengan metode Agarose Gel DNA Extraction Kit menunjukkan pita yang lebih bersih daripada purifikasi dengan metode fenol kloroform. Pita hasil purifikasi dengan metode fenol kloroform terlihat smear, hal ini menunjukkan masih terdapatnya pengotor Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 meskipun telah dilakukan proses purifikasi. Purifikasi metode Agarose gel DNA extraction kit adalah dengan menggunakan silika. Silika digunakan karena dapat mengabsorbsi DNA, tidak mengadsorbsi protein, RNA, dan oligonukleotida yang panjangnya kurang dari 50 pb sehingga dapat diperoleh DNA yang cukup murni. Purifikasi metode fenol kloroform merupakan campuran pelarut organik yang dapat mendenaturasi protein dan melarutkan komponen lipid, sehingga antara protein dan DNA dapat dipisahkan. Fenol dapat mengikat protein dan sebagian kecil RNA, sedangkan kloroform dapat membersihkan protein dan polisakarida dari larutan (Muladno, 2010). Purifikasi dengan metode Agarose Gel DNA Extraction Kit menunjukkan hasil yang lebih baik sehingga hasilnya digunakan untuk tahapan penelitian selanjutnya. Proses pemotongan ganda dengan enzim restriksi Nco I dan Xho I dilakukan produk PCR yang telah dipurifikasi. Hasil elektroforesis yang diperoleh menunjukkan pita yang berukuran 510 pb (Gambar 4.2 nomor 7), hal ini menunjukkan proses pemotongan ganda telah berhasil dilakukan. Beberapa faktor yang perlu diperhatikan dalam pemotongan menggunakan enzim restriksi diantaranya yaitu jumlah DNA, enzim, dan bufer dalam volume reaksi total yang sesuai. Hasil dari proses pemotongan ganda selanjutnya dilakukan purifikasi menggunakan metode fenol kloroform. Hasil purifikasi dengan metode fenol kloroform dilakukan pengecekan dengan dielektroforesis pada gel agarosa 1%. 1 2 3 750 pb 500 pb 510 pb Gambar 4.3 Elektroforesis Hasil Purifikasi Metode Fenol Kloroform Setelah Pemotongan Ganda. Keterangan: (1) Marker; (2) Kosong; (3) Hasil purifikasi. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Hasil elektroforesis menunjukkan terdapat hanya satu pita tanpa smear yang berukuran 510 pb (Gambar 4.3). Hal ini menunjukkan bahwa tahap akhir purifikasi dengan menggunakan metode fenol kloroform untuk penyiapan DNA insert telah berhasil dilakukan dengan baik. DNA insert (gen human ifn α2a) hasil purifikasi tahap akhir diatas telah siap digunakan pada tahapan selanjutnya untuk diligasikan ke vektor pET-32b(+) . 4.2 Penyiapan Vektor pET-32b(+) Vektor pET-32b(+) yang mempunyai konsentrasi 300 ng/µl terlebih dahulu dilakukan pemotongan dengan menggunakan enzim restriksi. Untuk mengetahui kemampuan pemotongan enzim restriksi Nco I pada vektor, maka dilakukan pemotongan ganda dengan menggunakan enzim restriksi lainnya yaitu Mlu I. Hasil pemotongan menggunakan enzim restriksi Nco I dan Mlu I akan terlihat dua pita yang ukurannya berbeda. Vektor pET-32b(+) selanjutnya dilakukan pemotongan ganda menggunakan enzim restriksi Nco I dan Xho I. Hasil dari pemotongan kemudian dielektroforesis pada agel agarosa 1%, dilanjutkan pewarnaan dengan larutan EtBr dan diamati diatas sinar UV. 1 2 3 5845 pb 4 5 5899 pb 4590 pb 1309 pb 1500 pb 1000 pb Gambar 4.4 Elektroforesis Hasil Pemotongan Vektor pET-32b(+). Keterangan: (1) Vektor pET-32b(+) dipotong Xho I dan Nco I; (2) Vektor pET-32b(+) dipotong Nco I dan Mlu I; (3) Kosong; (4) Marker; (5) Vektor pET-32b(+). Hasil elektroforesis menunjukkan bahwa enzim Xho I dan Nco I kondisinya masih bagus dan mampu untuk memotong vektor pET-32b(+) Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 (Gambar 4.4). Hal ini terbukti dari hasil pemotongan ganda menggunakan enzim Mlu I menghasilkan dua pita yang ukurannya benar. Vektor pET-32b(+) yang dipotong dengan enzim Nco I dan enzim Mlu I diperoleh dua pita yang berukuran 4590 pb dan 1309 pb. Hasil pemotongan ganda vektor pET-32b(+) dengan enzim Xho I dan Nco I terlihat pita yang mempunyai ukuran 5845 pb. Pita kecil yang berukuran 54 pb tidak dapat terlihat dikarenakan konsentrasinya yang kecil dan sewaktu proses elektroforesis pita tersebut bermigrasi dengan cepat pada gel agarosa 1% sehingga keluar dari gel. Terdapatnya pita yang berukuran 5845 pb menunjukkan bahwa pemotongan ganda pada vektor pET-32b(+) dengan enzim Xho I dan Nco I telah berhasil dilakukan. Vektor pET-32b(+) yang telah dipotong ganda menggunakan enzim Xho I dan Nco I selanjutnya dilakukan purifikasi menggunakan metode fenol kloroform. Purifikasi dilakukan pada vektor pET-32b(+) yang telah terpotong bertujuan untuk menghilangkan sisa-sisa enzim, dan pengotor-pengotor lainnya. Pemurnian DNA yang umum dilakukan yaitu melalui penambahan larutan fenol dan kloroform dengan perbandingan 1:1. Pengendapan DNA dilakukan menggunakan cara presipitasi etanol absolut dan dipisahkan dengan cara disentrifus (Radji, 2011). Hasil purifikasi vektor pET-32b(+) dengan metode fenol kloroform dilakukan pengecekan dengan dielektroforesis pada gel agarosa 1%. 1 2 3 4 5899 pb 5845 pb Gambar 4.5 Elektroforesis Hasil Purifikasi Vektor pET-32b(+) Setelah Pemotongan Ganda dengan Enzim Xho I dan Nco I. Keterangan: (1) Marker; (2) Vektor pET-32b(+) dicek 2 µl; (3) Vektor pET32b(+) dipotong Xho I dan Nco I lalu dipurifikasi dicek 2 µl; (5) Vektor pET32b(+) dipotong Xho I dan Nco I dicek 3 µl. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Elektroforesis hasil purifikasi metode fenol kloroform pada vektor pET32b(+) yang telah dilakukan pemotongan ganda dengan enzim Xho I dan Nco I menunjukkan hanya satu pita dan tidak ada smear (Gambar 4.5). Hal ini menunjukkan bahwa proses purifikasi vektor pET-32b(+) telah berhasil dilakukan. Vektor pET-32b(+) telah bersih dan siap digunakan untuk diligasikan ke DNA insert (gen human ifn α2a) yang telah dipersiapkan. Tahapan ligasi harus menggunakan DNA insert dan vektor yang bersih supaya proses ligasinya benar dan tidak mengalami gangguan. 4.3 Kloning Gen human ifn α2a Kloning diawali dengan memasukkan DNA insert ke dalam vektor. Ligasi dilakukan antara vektor pET-32b(+) yang dilinearkan dengan gen human ifn α2a (insert) yang telah dipersiapkan sebelumnya. Proses ligasi menggunakan enzim ligase yang berfungsi untuk mengkatalis pembentukan ikatan kovalen DNA untuk menyisipkan sepotong DNA ke dalam vektor (Radji, 2011). Pada penelitian ini reaksi ligasi dilakukan dengan menggunakan enzim T4 DNA ligase yang diinkubasi pada suhu 4⁰C semalam. Komposisi vektor dan insert yang digunakan untuk proses ligasi adalah sekitar 1:3. Hasil dari proses ligasi diharapakan diperolehnya vektor rekombinan pET-32b(+)-IFN α2a yang sirkuler. Reaksi ligasi dikerjakan sedemikian rupa sehingga akan meghasilkan rekombinan yang banyak (Sudjadi, 2008) Hasil ligasi kemudian ditransformasikan pada sel kompeten E. coli DH5α sebagai inang kloning. Sel E. coli DH5α mempunyai laju rekombinasi yang rendah sehingga memungkinkan digunakan untuk pemeliharaan yang stabil plasmid DNA (Tolia dan Joshua-Tor, 2006). Faktor penting yang mempengaruhi efisiensi transformasi adalah kualitas dari sel kompeten (Xiaowei Li et al., 2010). Sel E. coli dibuat kompeten supaya permeabilitas dinding sel meningkat sehingga vektor rekombinan lebih mudah masuk ke dalam sel. Sel E. coli dibuat kompeten dengan penambahan larutan CaCl2 dingin. Laruan CaCl2 dalam keadaan dingin efektif menyebabkan perubahan permeabilitas dinding sel bakteri menjadi lebih permiabel (Radji, 2011). Perlakuan heat shock yang diikuti dengan pendinginan akan menjadikan dinding sel kembali seperti semula. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Proses transformasi dilakukan dengan metode heat shock pada suhu 42⁰C selama 90 detik (Radji, 2011). Metode heatshock dipilih karena metodenya sederhana, pori-pori membran sel E. coli dapat terbuka dalam waktu yang singkat dan siap untuk menerima vektor rekombinan yang akan masuk. Hasil dari transformasi dicek keberhasilannya dengan cara ditumbuhkan pada media LB padat yang ditambahkan ampisilin. Sebagai kontrol negatif digunakan suspensi sel kompeten yang ditumbuhkan pada media LB padat yang ditambahkan ampisilin. Klon transforman E. coli diseleksi dengan menggunakan marker antibiotik ampisilin. Hanya klon E. coli yang mengandung vektor rekombinan pET-32b(+)IFN α2a yang dapat tumbuh pada media padat LB yang ditambah dengan ampisilin. A B C Gambar 4.6 Hasil Transformasi pada E. coli DH5α. Keterangan: (A) Inokulasi 80 µl; (B) Inokulasi 350 µl; (C) Kontrol negatif. Hasil transformasi ke inang kloning E. coli DH5α diperoleh 13 klon transforman (Gambar 4.6). Tiga klon transforman tumbuh pada inokulasi 80 µl Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 dan sepuluh klon transforman tumbuh pada inokulasi 350 µl. Proses transformasi ke sel kompeten E. coli DH5α telah berhasil dilakukan yang mana ditunjukkan dengan terdapatnya klon transforman yang tumbuh pada media LB padat yang ditambah ampisilin meskipun efisiensinya rendah. Efisiensi transformasi yang rendah kemungkinan disebabkan oleh kurang optimalnya proses ligasi yang dilakukan. Suhu optimum untuk ligasi merupakan kompromi antara kecepatan aksi enzim dan penyambungan kedua ujung yang berdasarkan percobaan sekitar 4-15⁰C (Sudjadi, 2008). Tingkat efisiensi transformasi juga dipengaruhi oleh konsentrasi DNA dan kondisi sel inang. Klon transforman yang diperoleh masih perlu dilakukan skrining dan verifikasi untuk mengetahui masuknya vektor rekombinan ke dalam sel E. coli DH5α. Pada kontrol negatif terlihat tidak ada yang tumbuh pada media LB padat yang telah ditambah ampisilin, hal ini menunjukkan sel kompeten E. coli DH5α yang digunakan untuk transformasi kondisinya baik. 4.4 Skrining dan Verifikasi Klon Transforman di E. coli DH5α Skrining dan verifikasi klon transforman pada penelitian ini dilakukan dengan beberapa metode yaitu PCR colony screening, isolasi vektor rekombinan pET-32b(+)-IFN α2a, analisis restriksi dan juga dilakukan sekuensing (pET Sistem Manual, 2006). Beberapa metode skrining dan verifikasi dilakukan bertujuan untuk memastikan keberhasilan ligasi gen human ifn α2a pada vektor pET-32b(+) membentuk vektor rekombinan (pET-32b(+)-IFN α2a) dan juga keberhasilan proses transformasi ke inang kloning E. coli DH5α. Proses seleksi klon bakteri ini tergolong sulit karena dari ribuan sel bakteri, hanya beberapa sel bakteri saja yang kemunginan menerima plasmid DNA rekombinan yang masuk ke dalam sel bakteri (Radji, 2011). Skrining koloni rekombinan E. coli untuk mengetahui terdapatnya gen target dapat dilakukan dengan metode PCR koloni. Metode PCR koloni merupakan metode yang mudah dan sederhana untuk skrining klon bakteri (Azhahianambi et al., 2008). Metode PCR colony screening dilakukan dengan menggunakan sepasang primer upstream dan downstream dari DNA insert. Sepasang primer dibuat yaitu primer pET_Screen_F dan pET_Screen_R yang Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 digunakan untuk mengetahui terdapatnya insert pada vektor pET-32b(+) sehingga membentuk vektor rekombinan (pET-32b(+)-IFN α2a) dan juga untuk mengetahui masuknya vektor rekombinan tersebut ke dalam sel E. coli DH5α. Skrining koloni dengan metode PCR juga dilakukan dengan menggunakan primer IFN_NcoI_F dan IFN_XhoI_R. Hasil skrining menggunakan metode PCR dan pada klon transforman 1-11 dicek dengan dielektroforesis pada agarosa 1%. 1 2 3 4 5 6 7 8 9 10 11 12 1500 pb 1000 pb 1100 pb Gambar 4.7 Elektroforesis Hasil PCR dengan Primer pET_Screen_F dan pET_Screen_R pada Klon Transforman 1-11 E. coli DH5α. Keterangan: (1) Marker; (2) Klon 1; (3) Klon 2; (4) Klon 3; (5) Klon 4 (6) Klon 5; (7) Klon 6; (8) Klon 7; (9) Klon 8; (10) Klon 9; (11) Klon 10; (12) Klon 11. 1 2 3 4 5 6 7 8 9 10 11 12 750 pb 500 pb 510 pb Gambar 4.8 Elektroforesis Hasil PCR dengan Primer IFN_NcoI_F dan IFN_XhoI_R pada Klon Transforman 1-11 E. coli DH5α. Keterangan: (1) Marker; (2) Klon 1; (3) Klon 2; (4) Klon 3; (5) Klon 4 (6) Klon 5; (7) Klon 6; (8) Klon 7; (9) Klon 8; (10) Klon 9; (11) Klon 10; (12) Klon 11. Elektroforesis hasil PCR colony screening dengan primer pET_Screen_F dan pET_Screen_R pada klon transforman 1-11 adalah positif semuanya dengan terdapatnya pita yang berukuran sekitar 1100 pb (Gambar 4.7). Hal ini Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 menunjukkan bahwa insert telah masuk pada vektor pET-32b(+) membentuk vektor rekombinan dan vektor rekombinan (pET-32b(+)-IFN α2a) telah masuk ke sel E. coli DH5α. Elektroforesis hasil PCR dengan primer IFN_NcoI_F dan IFN_XhoI_R pada klon transforman 1-11 menunjukkan positif semuanya dengan terdapatnya pita yang berukuran 510 pb (Gambar 4.8). Hasil keseluruhan menunjukkan bahwa pada 11 klon transforman terdapat insert gen human ifn α2a pada vektor rekombinan (pET-32b(+)-IFN α2a) yang telah masuk ke inang kloning E. coli DH5α. 4.5 Kultivasi dan Isolasi Vektor Rekombinan Klon transforman 1-11 hasil transformasi pada inang kloning E. coli DH5α selanjutnya dikultivasi pada media LB cair yang ditambah ampisilin untuk proses miniprep. Isolasi vektor rekombinan (miniprep) pada 11 klon transforman dilakukan dengan metode lisis alkali. Tahap pelisisan dilakukan pada suasana alkali (pH 12) karena molekul DNA kromosom terfragmentasi dan terdenaturasi, sedangkan DNA plasmid terdenaturasi tetapi masih saling terpaut antara masingmasing untai dan tidak terpotong karena ukurannya kecil (Sudjadi, 2008). Tujuan kultivasi dan isolasi plasmid adalah untuk memperoleh vektor rekombinan yang selanjutnya dilakukan analisis restriksi dan disekuensing. 1 2 3 4 5 6 7 8 9 10 11 12 6355 pb Gambar 4.9 Elektroforesis Hasil Miniprep Klon Transforman di E. coli DH5α. Keterangan: (1) Marker; (2) Klon 1; (3) Klon 2; (4) Klon 3; (5) Klon 4; (6) Klon 5; (7) Klon 6; (8) Klon 7; (9) Klon 8; (10) Klon 9; (11) Klon 10; (12) Klon 11. Elektroforesis hasil miniprep pada klon transforman 1-11 menunjukkan positif semua dengan terdapatnya pita yang berukuran 6355 pb pada sumuran 2 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 sampai 12 (Gambar 4.9). Hal ini menunjukkan bahwa proses miniprep pada klon transforman 1-11 telah berhasil dilakukan dengan baik. Dalam melakukan isolasi DNA plasmid harus memperhatikan keberadaan DNA genom yang berasal dari sel bakteri. Adanya kontaminasi dari DNA genom bakteri dapat mempengaruhi keberhasilan kloning DNA (Radji, 2011). Hasil isolasi vektor rekombinan (pET32b(+)-IFN α2a) pada klon transforman 1, 2, 3, 4, 11 dipilih untuk dilakukan pengecekan lebih lanjut menggunakan metode PCR. Proses PCR menggunakan primer gabungan antara primer IFN_NcoI_F dengan pET_Screen_R dan pET_Screen_F dengan IFN_XhoI_R. Hasil PCR yang diperoleh selanjutnya dilakukan cek dengan dielektroforesis pada gel agarosa 1,5%. 1 2 3 4 5 6 7 8 9 10 11 1000 pb 1000 pb 600 pb 500 pb Gambar 4.10 Elektroforesis Skrining Metode PCR Menggunakan Primer (IFN_NcoI_F dengan pET_Screen_R) dan (Primer pET_Screen_F dengan IFN_XhoI_R) pada Hasil Miniprep Klon 1, 2, 3, 4, dan Klon 11 E. coli DH5α. Keterangan: (1) Klon 1; (2) Klon 2; (3) Klon 3; (4) Klon 4; (5) Klon 11; (6) Marker; (7) Klon 1; (8) Klon 2 ; (9) Klon 3; (10) Klon 4; (11) Klon 11. Elektroforesis hasil PCR dengan primer IFN_NcoI_F dan pET_Screen_R pada hasil miniprep klon transforman 1, 2, 3, 4, dan klon 11 menunjukkan positif semuanya dengan terdapatnya pita yang berukuran benar disekitar 600 pb (Gambar 4.10 nomor 1-5). Elektroforesis hasil PCR dengan primer pET_Screen_F dengan IFN_XhoI_R pada hasil miniprep klon transforman 1, 2, 3, 4, dan klon 11 juga menunjukkan hasil positif semuanya dengan terdapatnya pita yang berukuran Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 benar disekitar 1000 pb. Hal ini menunjukkan bahwa pada klon transforman 1, 2, 3, 4, dan klon 11 terdapat vektor rekombinan (pET-32b(+)-IFN α2a) yang telah masuk ke dalam inang E. coli DH5α. Hasil minirep vektor rekombinan pada klon transforman 2 selanjutnya dipilih dan dipurifikasi dengan menggunakan metode fenol kloroform. Proses purifikasi bertujuan untuk menghilangkan RNAse pada proses isolasi DNA rekombinan. RNAse digunakan pada proses miniprep berfungsi untuk membersihkan DNA dari RNA (Radji, 2011). Metode analisis restriksi dilakukan pada hasil miniprep klon transforman 2 yang sudah dilakukan purifikasi. Hasil miniprep pada klon transforman 2 dilakukan pemotongan ganda dengan menggunakan enzim restriksi Xho I dan Nco I. Vektor rekombinan (pET-32b(+)IFN α2a) yang telah dipotong selanjutnya dilakukan pengecekan dengan dielektroforesis pada gel agarosa 1%. 1 2 3 5899 pb 5845 pb 750 pb 500 pb 510 pb Gambar 4.11 Elektroforesi Hasil Pemotongan Vektor Rekombinan Klon 2. Keterangan: (1) Marker; (2) Vektor pET-32b(+); (3) Vektor rekombinan klon 2 dipotong enzim Xho I dan Nco I. Pemotongan hasil miniprep vektor rekombinan klon transforman 2 yang sudah dipurifikasi dengan metode menghasilkan dua buah pita yang berukuran 5845 pb dan 510 pb (Gambar 4.11). Pita yang berukuran 5845 pb menunjukkan vektor pET-32b(+), sedangkan pita yang berukuran 510 pb menunjukkan terdapatnya insert. Hasil yang diperoleh menunjukkan bahwa vektor rekombinan pada klon transforman 2 telah berhasil dipotong oleh enzim Xho I dan Nco I dan mempunyai insert (gen human ifn α2a). Klon transforman 2 selanjutnya dipilih Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 untuk dikultivasi pada media LB cair 200 mL yang telah ditambah ampisilin untuk perbanyakan vektor rekombinan (pET-32b(+)-IFN α2a). Isolasi dan purifikasi vektor rekombinan dilakukan dengan menggunakan Qiagen Plasmid Maxi Kit. Hasil Qiagen Plasmid Maxi Kit selanjutnya diukur konsentrasinya dengan menggunakan alat Nanophotometer. Hasil pengukuran menunjukkan A260/A280: 1,851 dan diperoleh konsentrasi 405 ng/µl. Kemurnian larutan DNA dihitung dengan membandingkan absorbansi pada panjang gelombang 260 nm dan absorbansi pada panjang gelombang 280 nm. Nilai kemurnian DNA hasil isolasi dan purifikasi menggunakan Qiagen Plasmid Maxi Kit adalah 1,851 sehingga telah murni dan dapat digunakan untuk tahapan selanjutnya. Rasio A 260/A280 antara 1,8-2,0 mencerminkan DNA yang relatif murni dan terbebas dari kontaminan protein (Muladno, 2010). 1 2 6355 pb Gambar 4.12 Elektroforesis Hasil Qiagen Plasmid Maxi Kit. Keterangan: (1) Marker; (2) Hasil Qiagen Plasmid Maxi Kit Klon 2. Elektroforesis hasil Qiagen Plasmid Maxi Kit pada gel agarosa 1% menunjukkan terdapatnya pita yang berukuran 6355 pb (Gambar 4.12). Hal ini menunjukkan bahwa proses isolasi dan purifikasi menggunakan Qiagen Plasmid Maxi Kit telah berhasil dilakukan dan diperoleh vektor rekombinan dengan konsentrasi 405 ng/ µl. Hasil elektroforesis vektor rekombinan terdapat dua pita menunjukkan vektor yang dicek mempunyai 2 bentuk yaitu supercoil dan relaxed circular. Urutan migrasi bentuk-bentuk vektor pada elektroforesis gel dari yang paling cepat adalah supercoil, supercoiled denaturated, relaxed circular, linear, Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 dan nicked open circular. Vektor rekombinan (pET-32b(+)-IFN α2a) dari klon 2 hasil Qiagen Plasmid Maxi Kit selanjutnya dilakukan analisis sekuensing untuk mengetahui urutan nukleotidanya. Sekuensing DNA merupakan teknik yang dapat dimanfaatkan untuk menentukan urutan basa nukleotida suatu gen ataupun sekuen genom total suatu sel atau organisme (Radji, 2011). Analisis urutan nukleotida dengan metode sekuensing dilakukan di perusahaan Macrogen Korea dengan menggunakan dua primer yaitu Stag-18mer-primer dan T7terminator-primer. Gambar 4.13 Hasil Alignment Sekuen Klon 2 dengan Stag-18mer-primer. Gambar 4.14 Hasil Alignment Sekuen Klon 2 dengan T7terminator-primer. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Urutan nukleotida hasil sekuensing vektor rekombinan klon transforman 2 selanjutnya disejajarkan dengan urutan nukleotida human IFN α2a pada GenBank menggunakan program MultAlin (Multiple sequence alignment). Berdasarkan analisis urutan nukleotida menggunakan primer Stag-18mer-primer dan T7terminator-primer menunjukkan bahwa urutan nukleotida DNA insert (gen human ifn α2a) klon transforman 2 memiliki urutan yang sama dengan urutan nukleotida human IFN α2a pada GenBank DI084466.1 dan tidak terdapat mutasi (Gambar 4.13 dan 4.14). Berdasarkan data yang diperoleh, maka dapat disimpulkan bahwa pada klon transforman 2 telah membawa gen human ifn α2a yang benar dalam vektor rekombinan sehingga vektor rekombinan (pET-32b(+)IFN α2a) dapat digunakan untuk tahapan penelitian selanjutnya. 4.6 Transformasi Vektor Rekombinan ke E. coli BL21(DE3) Vektor rekombinan (pET-32b(+)-IFNα2a) dari klon transforman 2 dengan insert yang benar selanjutnya ditransformasikan ke sel E. coli galur BL21(DE3) sebagai inang ekspresi. Sistem ekspresi untuk produksi protein rekombinan dalam E. coli melalui kombinasi antara vektor dan galur E. coli sebagai inang ekspresi (Sørensen dan Mortensen, 2005). Proses transformasi dilakukan dengan perlakuan kejut panas (heat shock) pada suhu 42⁰C selama 90 detik (Radji, 2011). Hasil dari transformasi selanjutnya ditumbuhkan pada media padat LB yang telah ditambahkan ampisilin. A B Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 C D E Gambar 4.15. Hasil Transformasi Vektor Rekombinan pada E. coli BL21(DE3). Keterangan: (A) Inokulasi 20 µl; (B) Inokulasi 50 µl; (C) Inokulasi 80 µl; (D) Inokulasi 350 µl; (E) Kontrol negatif. Hasil transformasi vektor rekombinan ke inang ekspresi sel kompeten E. coli BL21(DE3) diinokulasikan pada media LB padat yang mengandung ampisilin dengan metode sebar. Hasil transformasi vektor rekombinan (pET-32b(+)IFNα2a) dari klon transforman 2 ke sel kompeten E. coli BL21(DE3) menunjukkan diperolehnya banyak klon transforman (Gambar 4.15). Banyaknya klon transforman yang tumbuh menunjukkan bahwa proses transformasi yang dilakukan berhasil dengan baik. Escherichia coli merupakan inang umum yang digunakan untuk produksi protein rekombinan, meskipun terdapat gen yang kurang efisien apabila diekspresikan pada organisme ini (Srivastva et al., 2005). 4.7 Skrining dan Verifikasi Klon Transforman di E coli BL21 (DE3) Klon transforman E. coli BL21(DE3) yang diperoleh diseleksi menggunakan media padat LB selektif yang mengandung ampisilin. Verifikasi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 masuknya vektor rekombinan pET-32b(+)-IFN α2a pada inang ekspresi E. coli BL21(DE3) dilakukan dengan menggunakan metode PCR colony screening. Klon transforman dipilih 7 secara acak dan dilakukan PCR skrining menggunakan primer pET_Screen_F dengan pET_Screen_R dan IFN_NcoI_F dengan IFN_XhoI_R. Hasil dari PCR colony screening kemudian dilakukan pengecekan dengan metode elektroforesis pada medium agarosa 1%. 1 2 3 4 5 6 7 8 1500 pb 1000 pb 1100 pb Gambar 4.16 Elektroforesis Hasil PCR Skrining dengan Primer pET_Screen_F dan pET_Screen_R pada Klon Transforman E. coli BL21(DE3). Keterangan: (1) Marker; (2) Klon 1; (3) Klon 2; (4) Klon 3; (5) Klon 4; (6) Klon 5; (7) Klon 6; (8) Klon 7. 1 2 3 4 5 6 7 8 750 pb 500 pb 510 pb Gambar 4.17 Elektroforesis Hasil PCR Skrining dengan Primer IFN_NcoI_F dan IFN_XhoI_R pada Klon Transforman E. coli BL21(DE3). Keterangan: (1) Marker; (2) Klon 1; (3) Klon 2; (4) Klon 3; (5) Klon 4; (6) Klon 5; (7) Klon 6; (8) Klon 7. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Metode PCR colony screening pada ketujuh klon transforman di E. coli BL21(DE3) dilakukan menggunakan metode yang serupa dengan PCR colony screening pada klon transforman di E. coli DH5α. Elektroforesis hasil PCR skrining dengan primer pET_Screen_F dan pET_Screen_R pada ketujuh klon transforman diperoleh pita yang berukuran sekitar 1100 pb (Gambar 4.16). Hal ini menunjukkan bahwa vektor rekombinan (pET-32b(+)-IFNα2a) telah masuk ke inang ekspresi E. coli BL21(DE3). Ketujuh klon transforman juga dilakukan PCR skrining dengan menggunakan primer IFN_NcoI_F dan IFN_XhoI_R diperoleh pita berukuran 510 pb yang mana merupakan ukuran gen human ifn α2a (Gambar 4.17). Keselruhan hasil yang diperoleh menunjukkan bahwa vektor rekombinan (pET-32b(+)-IFNα2a) telah berhasil ditranformasikan pada inang ekspresi E. coli BL21(DE3) dengan baik. 4.8 Ekspresi, Isolasi dan Karakterisasi Protein Rekombinan Human IFN α2a Klon transforman E. coli BL21(DE3) yang telah dipastikan mempunyai vektor rekombinan (pET-32b(+)-IFNα2a) selanjutnya dilakukan uji ekspresi protein. Ketujuh klon transforman dilakukan uji ekspresi protein pada skala kecil dengan menggunakan medium produksi yaitu media LB cair 10 ml yang ditambah ampisilin dan diinkubasi pada suhu 37⁰C. Induksi dilakukan dengan penambahan IPTG pada kultur sel yang telah mencapai OD600 sekitar 0,5 dengan konsentrasi akhir IPTG 1 mM. Reagen IPTG berfungsi melepas represi promotor sehingga diperoleh ekspresi protein rekombinan dalam jumlah yang tinggi (Grompe et al., 1998). Pemanenan kultur dilakukan pada waktu 1, 2, dan 3 jam setelah inkubasi. Isolasi protein rekombinan human IFN α2a dilakukan pada protein yang terlarut pada medium kultivasi (supernatan) dan protein tidak terlarut (pelet). Pelet selanjutnya diresuspensi menggunakan bufer PBS 1x. Hasil isolasi protein pada supernatan dan pelet kemudian dianalisis menggunakan SDS-PAGE yang dilanjutkan dengan Western blot atau pewarnaan CBB. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 120 kDa 80 kDa 60 kDa 36 kDa 35 kDa 30 kDa 20 kDa Gambar 4.18 Western Blot Klon 1 Inkubasi 37⁰C IPTG 1 mM. Keterangan: (1) Pelet 0 jam; (2) Supernatan 0 jam; (3) Marker; (4) Pelet 1 jam; (5) Supernatan 1 jam; (6) Pelet 2 jam; (7) Supernatan 2 jam; (8) Pelet 3 jam; (9) Supernatan 3 jam. Hasil SDS PAGE yang dilanjutkan Western blot pada klon 1 yang diinkubasi pada suhu 37⁰C dengan induksi IPTG 1 mM menunjukkan terdapatnya pita tebal pada pelet dan pita tipis pada supernatan yang mempunyai berat molekul sekitar 36 kDa (Gambar 4.18). Pita tersebut merupakan berat molekul dari protein rekombinan IFN α2a yang terfusi dengan thioredoxin dan 6x histidin. Pendeteksian terdapatnya protein rekombinan IFN α2a pada Western blot menggunakan antibodi monoklonal anti interferon α yang spesifik. Waktu 0 jam merupakan level basal ekspresi protein sebelum dilakukan induksi. Hasil yang diperoleh juga menunjukkan bahwa inkubasi pada suhu 37⁰C dengan induksi IPTG 1 mM maka protein rekombinan human IFN α2a lebih dominan diekspresikan dalam bentuk tidak terlarut. Ekspresi protein rekombinan sebagai protein terfusi mempunyai manfaat dapat dibuat konstruksi gen yang lebih menguntungkan, memungkinkan ekspresi level tinggi protein terlarut (Kapust dan Waugh, 1999) dengan menurunkan kecenderungan pembentukan badan inklusi (Lilie et al., 1998). Formasi pelipatan dan pembentukan ikatan disulfida pada protein target dapat ditingkatkan dengan fusi thioredoxin pada E. coli galur defisiensi thioredoxin reductase (trxB) (Fathallah et al., 2009). Banyak protein yang normalnya diproduksi dalam bentuk tidak terlarut pada E. coli menjadi lebih terlarut jika difusikan dengan sekuen Nterminal thioredoxin (LaValline et al., 1993; Novy et al., 1995). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.19 Pewarnaan CBB Klon 2, 3, 6, dan 7 Inkubasi 37⁰C IPTG 1 mM. Keterangan: (1) Pelet 3 jam klon 7; (2) Supernatan 3 jam klon7; (3) Supernatan 3 jam klon 6; (4) Pelet 3 jam klon 6; (5) Supernatan 3 jam klon 2; (6) Pelet 3 jam klon 2; (7) Supernatan 3 jam klon 3; (8) Pelet 3 jam klon 3; (9) Pelet 0 jam klon 3; (10) Marker. 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.20 Pewarnaan CBB Klon 4 dan Klon 5 Inkubasi 37⁰C IPTG 1 mM. Keterangan: (1) Supernatan 4 jam klon 5; (2) Supernatan 3 jam klon 5; (3) Supernatan 4 jam klon 4; (4) Supernatan 3 jam klon 4; (5) Marker; (6) Pelet 4 jam klon 5; (7) Pelet 3 jam klon 5; (8) Pelet 4 jam klon 4; (9) Pelet 3 jam klon 4; (10) Pelet 0 jam klon 4. Hasil elektroforesis SDS-PAGE dengan pewarnaan CBB pada klon 2, 3, 6, dan klon 7 dengan membandingkan waktu inkubasi 3 jam setelah induksi IPTG juga menunjukkan adanya pita tebal yang mempunyai berat molekul sekitar 36 kDa pada pelet (Gambar 4.19). Hasil elektroforesis SDS-PAGE dengan pewarnaan CBB pada klon 4 dan klon 5 dengan membandingkan waktu inkubasi 3 jam dan 4 jam setelah induksi IPTG juga menunjukkan terdapatnya pita tebal yang mempunyai berat molekul sekitar 36 kDa pada pelet (Gambar 4.20). Hasil yang diperoleh menunjukkan bahwa inkubasi pada suhu 37⁰C dengan induksi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 IPTG 1 mM klon 2, 3, 4, 5, 6, dan klon 7 maka protein rekombinan human IFN α2a yang lebih dominan diekspresikan dalam bentuk tidak terlarut sama seperti hasil ekspresi pada klon 1. Hasil uji ekspresi skala kecil pada klon 1, 2, 3, 4, 5, 6, dan klon 7 menunjukkan bahwa protein rekombinan human IFN α2a telah berhasil diekspresikan pada sel E. coli BL21(DE3). Pita tebal protein terdapat pada E. coli setelah dilakukan induksi dengan IPTG. Hal ini menunjukkan bahwa overproduksi protein rekombinan human IFN α2a telah berhasil dilakukan. Protein rekombinan yang diperoleh dalam bentuk protein terfusi sehingga mengalami penambahan berat molekul. Protein rekombinan human IFN α2a mempunyai berat molekul 19 kDa dengan fusi tagnya mempunyai berat molekul 17 kDa sehingga berat molekul total adalah 36 kDa. Penelitian Yu-Ling Sun et al. (2011) tentang ekspresi rabbit neutrophil peptide-1 dalam vektor pET-32b(+) pada inang E. coli Rosetta-gami(DE3)pLysS menyatakan bahwa fusi Trx-(His)6tag mempunyai ukuran sekitar 17 kDa setelah dilakukan pemotongan menggunakan enterokinase. Ekspresi rabbit neutrophil peptide-1 dengan induksi IPTG 1 mM pada suhu 37⁰C dihasilkan protein terlarut setelah dilakukan ekstraksi dengan disonikasi. Klon 3 selanjutnya dipilih dan dilakukan uji ekspresi lebih lanjut menggunakan variasi suhu 28⁰C, 30⁰C, dan 37⁰C dengan induksi IPTG 1 mM. Pemanenan kultur setelah induksi dilakukan pada waktu 1, 2, 3, 4, 5, 6, 7 jam, dan kultur semalam. Variasi suhu dilakukan pada penelitian ini bertujuan untuk mencari optimasi suhu pada ekspresi protein rekombinan human IFN α2a terfusi sehingga diperoleh hasil ekspresi protein rekombinan human IFN α2a yang maksimal. Pemanenan kultur dilakukan pada berbagai waktu inkubasi bertujuan untuk mencari waktu yang tepat untuk pemanenan sehingga diperoleh hasil ekspresi maksimal protein rekombinan human IFN α2a. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 36 kDa Gambar 4.21 Pewarnaan CBB Pelet Klon 3 Inkubasi 37⁰C IPTG 1 mM. Keterangan: (1) Pelet 10 jam; (2) Pelet 9 jam; (3) Pelet 8 jam; (4) Pelet 7 jam; (5) Pelet 6 jam; (6) Pelet 5 jam; (7) Pelet 4 jam; (8) Pelet 3 jam; (9) Marker. 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.22 Pewarnaan CBB Supernatan Klon 3 Inkubasi 37⁰C IPTG 1 mM. Keterangan: (1) Marker; (2) Supernatan 0 jam; (3) Supernatan 3 jam; (4) Supernatan 4 jam; (5) Supernatan 5 jam; (6) Supernatan 6 jam; (7) Supernatan 7 jam; (8) Supernatan 8 jam; (9) Supernatan 9 jam; (10) Supernatan 10 jam. Hasil SDS PAGE dengan pewarnaan CBB pada klon 3 yang diinkubasi pada suhu 37⁰C dengan induksi IPTG 1 mM menunjukkan terdapatnya pita yang mempunyai berat molekul sekitar 36 kDa (Gambar 4.21 dan Gambar 4.22). Perbandingan hasil ekspresi protein pada supernatan dan pelet menunjukkan bahwa ekspresi protein rekombinan human IFN α2a dengan waktu pemanenan 3, 4, 5, 6, 7, 8, 9, dan 10 jam setelah induksi lebih dominan diekspresikan dalam bentuk tidak terlarut yaitu terdapat pada pelet sel. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.23 Western Blot Klon 3 Inkubasi 30⁰C IPTG 1 mM. Keterangan: (1) Marker; (2) Pelet 0 jam; (3) Pelet 5 jam; (4) Supernatan 5 jam; (5) Pelet 6 jam; (6) Supernatan 6 jam; (7) Pelet 7 jam; (8) Supernatan 7 jam; (9) Pelet semalam; (9) Supernatan semalam. 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.24 Pewarnaan CBB Klon 3 Inkubasi 30⁰C IPTG 1 mM. Keterangan: (1) Pelet 4 jam; (2) Pelet 3 jam; (3) Pelet 2 jam; (4) Pelet 1 jam; (5) Marker; (6) Supernatan 4 jam; (7) Supernatan 3 jam; (8) Supernatan 2 jam; (9) Supernatan 1 jam; (10) Supernatan 0 jam. Hasil SDS PAGE dengan pewarnaan CBB pada klon 3 yang diinkubasi pada suhu 30⁰C dengan induksi IPTG 1 mM menunjukkan terdapatnya pita yang mempunyai berat molekul sekitar 36 kDa (Gambar 4.23 dan Gambar 4.24). Perbandingan hasil ekspresi protein pada supernatan dan pelet menunjukkan bahwa ekspresi protein rekombinan human IFN α2a dengan waktu pemanenan 1, 2, 3, 4, 5, 6, 7 jam dan semalam setelah induksi lebih dominan diekspresikan dalam bentuk tidak terlarut pada pelet sel. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.25 Pewarnaan CBB Pelet Klon 3 Inkubasi 28⁰C IPTG 1 mM. Keterangan: (1) Pelet semalam; (2) Pelet 8 jam; (3) Pelet 7 jam; (4) Pelet 6 jam; (5) Pelet 5 jam; (6) Pelet 4 jam; (7) Pelet 3 jam; (8) Pelet 2 jam; (9) Marker; (10) Pelet 0 jam. 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.26 Pewarnaan CBB Supernatan Klon 3 Inkubasi 28⁰C IPTG 1 mM. Keterangan: (1) Pelet 1 jam; (2) Marker; (3) Supernatan 2 jam; (4) Supernatan 3 jam (5) Supernatan 4 jam; (6) Supernatan 5 jam; (7) Supernatan 6 jam (8) Supernatan 7 jam; (9) Supernatan 8 jam; (10) Supernatan semalam. Hasil SDS PAGE dengan pewarnaan CBB pada klon 3 yang diinkubasi pada suhu 28⁰C dengan induksi IPTG 1 mM menunjukkan terdapatnya pita yang mempunyai berat molekul sekitar 36 kDa (Gambar 4.25 dan Gambar 4.26). Perbandingan hasil ekspresi protein pada supernatan dan pelet menunjukkan bahwa ekspresi protein rekombinan human IFN α2a dengan waktu pemanenan 1, 2, 3, 4, 5, 6, 7, 8 jam dan semalam setelah induksi lebih dominan diekspresikan dalam bentuk tidak terlarut yaitu terdapat pada pelet sel. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Keberhasilan ekspresi protein rekombinan dipengaruhi oleh banyak faktor diantaranya plasmid dengan fusi tag yang digunakan, galur E. coli untuk inang ekspresi dan sifat protein yang diekspresikan. Beberapa strategi yang berbeda dapat digunakan tergantung pada kebutuhan protein untuk diekspresikan. Ekspresi protein yang toksik pada sel inang dapat menyebabkan lisisnya sel sehingga hasil yang diperoleh kurang optimal (Tolia dan Joshua-Tor, 2006). Pada penelitian ini telah berhasil dilakukan ekspresi protein rekombinan human IFN α2a pada E. coli galur BL21(DE3). 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.27 Pewarnaan CBB Pelet Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM. Keterangan: (1) Marker; (2) Pelet 3 jam (28⁰C); (3) Pelet 4 jam (28⁰C); (4) Pelet 5 jam (28⁰C); (5) Pelet 3 jam (30⁰C); (6) Pelet 4 jam (30⁰C); (7) Pelet 5 jam (30⁰C); (8) Pelet 3 jam (37⁰C); (9) Pelet 4 jam (37⁰C); (10) Pelet 5 jam (37⁰C). 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.28 Western Blot Pelet Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM. Keterangan: (1) Marker; (2) Pelet 3 jam (28⁰C); (3) Pelet 4 jam (28⁰C); (4) Pelet 5 jam (28⁰C); (5) Pelet 3 jam (30⁰C); (6) Pelet 4 jam (30⁰C); (7) Pelet 5 jam (30⁰C); (8) Pelet 3 jam (37⁰C); (9) Pelet 4 jam (37⁰C); (10) Pelet 5 jam (37⁰C). Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 1 2 3 4 5 6 7 8 9 10 36 kDa Gambar 4.29 Western Blot Supernatan Klon 3 Suhu 28⁰C, 30⁰C, 37⁰C IPTG 1 mM. Keterangan: (1) Marker; (2) Supernatan 3 jam (28⁰C); (3) Supernatan 4 jam (28⁰C); (4) Supernatan 5 jam (28⁰C); (5) Supernatan 3 jam (30⁰C); (6) Supernatan 4 jam (30⁰C); (7) Supernatan 5 jam (30⁰C); (8) Supernatan 3 jam (37⁰C); (9) Supernatan 4 jam (37⁰C); (10) Supernatan 5 jam (37⁰C). Hasil SDS PAGE klon 3 dengan pewarnaan CBB (Gambar 4.27) dan Western blot (Gambar 4.28) yang membandingkan hasil ekspresi protein rekombinan human IFN α2a pada pelet sel yang diinduksi IPTG 1 mM dengan variasi suhu 28⁰C, 30⁰C, dan 37⁰C menunjukkan bahwa ekspresi optimal diperoleh pada suhu inkubasi 37⁰C dengan rentang waktu pemanenan antara 3 sampai 5 jam setelah induksi. Hasil SDS PAGE klon 3 dengan Western blot (Gambar 4.29) yang membandingkan hasil ekspresi protein rekombinan human IFN α2a pada supernatan yang diinduksi IPTG 1 mM dengan variasi suhu 28⁰C, 30⁰C, dan 37⁰C menunjukkan bahwa ekspresi maksimal diperoleh pada suhu inkubasi 30⁰C dengan waktu pemanenan 3 jam setelah induksi. Kuantifikasi data tingkat ekspresi pada penelitian ini dilakukan dengan cara analisis densitometri pita hasil Western blot pada pelet (Gambar 4.28) dan supernatan (Gambar 4.29) menggunakan sistem digitalisasi automatik program UN-SCAN-IT Gel versi 6.1. Kuantifikasi yang dilakukan yaitu dengan membandingkan hasil piksel total pada masing-masing sampel yang terdeteksi sehingga dapat dibandingkan tingkat ekspresi pada masing-masing perlakuan dan diketahui hasil yang maksimal. Piksel total merupakan jumlah total keseluruhan piksel dalam area pita. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Tabel 4.1 Hasil Kuantifikasi Tingkat Ekspresi pada Pelet dan Supernatan. Perlakuan 3 jam (28⁰C) 4 jam (28⁰C) 5 jam (28⁰C) 3 jam (30⁰C) 4 jam (30⁰C) 5 jam (30⁰C) 3 jam (37⁰C) 4 jam (37⁰C) 5 jam (37⁰C) Piksel Total Pelet Supernatan 143960 71505 154760 77770 149549 82115 142012 129810 135055 87573 150057 65391 148327 39584 173122 24274 181773 18872 Hasil kuantifikasi menggunakan program UN-SCAN-IT menunjukkan bahwa piksel total tertinggi pada pelet diperoleh pada suhu inkubasi 37⁰C dengan waktu pemanenan 5 jam, sedangkan piksel total tertinggi pada supernatan diperoleh pada suhu inkubasi 30⁰C dengan waktu pemanenan 3 jam (Tabel 4.1). Oleh karena itu, dapat diambil kesimpulan bahwa ekspresi protein rekombinan human IFN α2a yang diinduksi IPTG 1 mM dengan variasi suhu 28⁰C, 30⁰C, dan 37⁰C menunjukkan hasil ekspresi maksimal pada pelet sel diperoleh pada suhu inkubasi 37⁰C dengan waktu pemanenan 5 jam setelah induksi. Hasil ekspresi maksimal pada supernatan diperoleh pada suhu inkubasi 30⁰C dengan waktu pemanenan 3 jam setelah induksi. Berdasarkan keseluruhan hasil penelitian, maka hasil terbaik ditunjukkan pada ekspresi protein rekombinan human IFN α2a yang diinduksi IPTG 1 mM pada suhu inkubasi 37⁰C dengan waktu pemanenan 5 jam setelah induksi. Isolasi protein human IFN α2a pada pelet sel pada penelitian ini dilakukan dengan pemecahan dinding sel bakteri menggunakan metode sonikasi. Sonikasi merupakan aplikasi gelombang ultrasonik untuk mengaduk partikel dalam suatu sampel yang dapat digunakan untuk mempercepat pelarutan suatu materi dengan memecah reaksi intermolekuler. Isolasi pada pelet sel bertujuan untuk mengetahui protein human IFN α2a diekspresikan dalam bentuk terlarut atau bentuk agregrat yang tidak terlarut (badan inklusi) pada sitoplasma. Isolasi badan inklusi Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 dilakukan dengan metode sentrifugasi dan solubilisasi menggunakan bufer yang mengandung guanidine hydrochloride dan merkaptoetanol. 1 2 3 4 5 36 kDa Gambar 4.30 Pewarnaan CBB Hasil Isolasi Protein dari Pelet. Keterangan: (1) Badan inklusi; (2) Pencucian ke 2; (3) Pencucian ke 1; (4) Protein terlarut; (5) Marker. Hasil SDS PAGE dengan pewarnaan CBB menunjukkan bahwa pita protein rekombinan human IFN α2a yang mempunyai berat molekul sekitar 36 kDa lebih banyak diekspresikan dalam bentuk terlarut dibandingkan bentuk badan inklusi (Gambar 4.30). Pada proses pencucian yang pertama juga masih menunjukkan terdapatnya protein rekombinan human IFN α2a (Gambar 4.30 no 3). Hal ini bisa disebabkan pada proses pengambilan supernatan masih terdapatnya sisa-sisa pada tabung. Hasil penelitian yang telah diperoleh menunjukkan bahwa protein rekombinan human IFN α2a lebih dominan diekspresikan dalam bentuk terlarut di sitoplasma. Ekspresi protein rekombinan dalam bentuk terlarut mempunyai keuntungan diantaranya yaitu proses purifikasinya lebih menghemat biaya dan tidak memakan waktu daripada refolding dan purifikasi pada badan inklusi (Fathallah et al., 2009). Data peneletian yang sebelumnya menyatakan bahwa protein IFN α yang diekspresikan pada E.coli seringnya dalam bentuk badan inklusi dalam sitoplasma (Swaminathan dan Khanna, 1999; Bedarrain et al., 2001; Srivasta et al., 2005). Berbagai strategi untuk meningkatkan kelarutan protein rekombinan yang diproduksi pada E. coli dapat dilakukan dengan cara pembatasan agregrasi invivo protein yang diantaranya melaui kultivasi pada temperatur yang lebih rendah, Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 penurunan konsentrasi penginduksi, dan sistem ekspresi dengan promotor yang indusibel. Level induksi yang rendah juga dilaporkan dapat meningkatkan kelarutan protein yang diekspresikan (Baneyx dan Mujacic, 2004). Oleh karena itu, perlu dilakukan optimasi faktor-faktor lingkungan seperti temperatur, media, dan konsentrasi penginduksi dalam mengekspresikan suatu gen. Pada penelitian ini telah berhasil diperoleh protein terfusi rekombinan human IFN α2a dalam bentuk terlarut di sitoplasma yang diekspresikan pada inang E. coli BL21(DE3) menggunakan media LB dengan suhu inkubasi 37⁰C dan waktu pemanenan 5 jam setelah induksi IPTG 1 mM. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 BAB 5 KESIMPULAN DAN SARAN 5.1 Kesimpulan Berdasarkan hasil penelitian secara keseluruhan menunjukkan bahwa gen human ifn α2a telah berhasil dimasukkan ke dalam vektor pET-32b(+) membentuk vektor rekombinan pET-32b(+)-IFN α2a. Vektor rekombinan juga telah berhasil ditransformasikan pada inang kloning E. coli DH5α dan selanjutnya ditansformasikan pada inang ekspresi E. coli BL21(DE3). Ekspresi protein rekombinan human IFN α2a dengan induksi IPTG 1 mM pada variasi suhu 28⁰C, 30⁰C, dan 37⁰C menunjukkan bahwa hasil optimal diperoleh pada suhu inkubasi 37⁰C dengan waktu pemanenan 5 jam setelah induksi. Protein rekombinan human IFN α2a yang diperoleh lebih dominan diekspresikan dalam bentuk terlarut di sitoplasma sebagai protein terfusi dengan berat molekul sekitar 36 kd. 5.2 Saran Protein rekombinan human IFN α2a pada penelitian masih perlu dilakukan pemisahan fusi tag dan purifikasi sehingga didapatkan protein rekombinan yang murni. Protein rekombinan human IFN α2a yang telah murni perlu diuji aktivitasnya secara in vitro mengunakan sel mamalia dan uji in vivo menggunakan hewan uji dengan dibandingkan IFN α2a komersial. Hasil penelitian ini juga perlu dilakukan scale up untuk mendapatkan protein rekombinan human IFN α2a dalam jumlah yang lebih banyak. 68 Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 DAFTAR PUSTAKA Arbabi M., Alasti F., Sanati M.H., Hosseini S., Deldar A., and Maghsoudi N. (2003). Kloning and expression of human gamma-interferon cDNA in E. Coli. Iraian Journal of Biotechnology, 1 (2), 87-94. Aulanni’am. (2004). Prinsip dan Teknik Analisis Biomolekul. Fakultas Pertanian Universitas Brawijaya Press. Azhahianambi P., Ghosh S., Kumar A., and Suryanaraya V.V.S. (2008). Cost effectiveness of colony lysis and colony PCR methods for screening of recombinant Escherichia coli colonies--a comparative study. Indian J Exp Biol. 46 (10), 731-735. Baneyx F., and Mujacic M. (2004). Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol., 22, 1399-1408. Bedarrain A., Cruz Y., Cruz O., Navarro M., and Gil M. (2001). Purification and conformational properties of a human interferon alpha2b produced in Escherichia coli. Biotechnol. Appl. Biochem., 33, 173-182. Brassard D.L., Grace M.J., and Bordens R.W. (2002). Interferon-α as an immunotherapeutic protein. Journal of Leukocyte Biology, 71, 565-581. Chiti F., Stefani M., Taddei N., Ramponi G., and Dobson C.M. (2003). Rationalization of the effects of mutations on peptide and protein aggregation rates. Nature, 424, 805-808. Fathallah M.D., Carthage T., Rabhi-Essafi I., and Coteaux M. (2009). Method for the production of high-level soluble human recombinant interferon alpha in E. coli and vectors useful for such a production. US Patent Application Publication. US 2009/0258394 A1. Fuh G., Mulkerrin M. G., Bass S., McFarland N., Brochier M., Bourell J. H., Light D. R., and Wells J. A. (1990). The human growth hormone receptor. Secretion from Escherichia coli and disulfide bonding pattern of the extracellular binding domain. J. Biol. Chem., 265, 3111-31153. Grompe M., Johnson W., and Jameson L. (1998). Recombinant DNA and genetic techniques. In Principles of molecular medicine. Edited by J. Larry Jameson. Humana Press Inc. Totowa, New Jersey. Hodgson J. (1993). Expression sistems: a user’s guide. Bio/Technology, 11, 887893. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Jeong W., and Shin H.C. (1998). Supply of the argU gene product allows highlevel expression of recombinant human interferon-α2a in Escherichia coli. Biotechnology Letters, 20 (1), 19-22. Jonasch E., and Haluska F.G. (2001). Interferon in oncological practice : review of interferon biology, clinical application, and toxicities. The Oncologist, 6, 34-55. Kapust R.B., and Waugh D.S. (1999). Escherichia coli maltose-binding protein is uncommonly effective at promoting the solubility of polypeptides to which it is fused. Protein Sci., 8, 1668-1674. Klaus W., Gsella B., Labhardta A.M., Wipfa B., Senn H. (1997). The threedimensional high relarutan structure of human interferon α-2a determined by heteronuclear NMR spectroscopy in larutan. Journal of Molecular Biology, 274 (4), 661-675. Kiefhaber T., Rudolph R., Kohler H.H., and Buchner J. (1991). Protein aggregation in vitro and in vivo: a quantitative model of the kinetic competition between folding and aggregation. Biotechnology (NY), 9, 825829. LaVallie E.R., DiBlasio E.A., Kovacic S., Grant K.L., Schendel P.F., and McCoy J.M. (1993). A thioredoxin gene fusion expression system that circumvents inclusion body formation in the E. coli cytoplasm. Bio/Technology, 11, 18793. Lilie H., Schwarz E., and Rudolph,R. (1998). Advances in refolding of proteins produced in E. coli. Curr. Opin. Biotechnol., 9, 497-501. Makrides S.C. (1996). Strategies for achieving high-level expression of genes in Escherichia coli. Microbiol Rev., 60 (3), 512-538. Meager A. (2006). The Interferons: characterization and application. WILEYVCH Verlag GmbH & Co. KGaA, Weinheim. Middelberg A. (2002). Preparative protein refolding. Trend Biotechnol., 20, 437. Muladno. (2010). Teknologi rekayasa genetika, Edisi Kedua. Penerbit IPB Press. Bogor. Mustofa I., Mahaputra L., Dachlan Y.P., Rantam F.A., dan Hinting A. (2006). Analisis densitometrik protein reseptor fertilisasi (ZP3) pada zona pelusida kambing sebagai kandidat bahan imunokontrasepsi. Media Kedokteran Hewan., 22 (2). Novagen. https://wasatch.biochem.utah.edu/chris/links/pET32.pdf. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Novy R., Berg J., Yaeger K., and Mierendorf,R. (1995). inNovations, 3, 7-9. Obeid D., and Bauvois B. (2006). Interferons: mechanisms, biological activities and survey of their use in human diseases. Current Bioactive Compounds, 2 (4), 431-444. Pang K.R., Wu J.J., Huang D.B., Tyring S.K., and Baron S. (2005). Biological and clinical basis for molecular studies of interferons. In Interferon methods and protocols. Edited by Daniel J.J. Carr. Humana Press Inc., Totowa, New Jersey. pET Sistem Manual (11th ed). (2006). Novagen. EMD Biosciences Inc., an affiliate of Merck KGaA, Darmstadt, Germany. Rabhi-Essafi I., Sadok A., Khalaf S.A., and Fathallah D.M. (2007). A strategy for high-level expression of soluble and functional interferon f as a GST fusion protein in E. coli. Protein Engineering, Design & Selection, 20 (5), 201-209. Radji M. (2011). Rekayasa genetika pengantar untuk profesi kesehatan. Penerbit Sagung Seto. Jakarta. Rezvani K., Teng Y., Pan Y., Dani J.A., Lindstrom J., Gras E.A.G., McIntosh J.M., and Biasi M.D. (2009). UBXD4, a UBX-containing protein, regulates the cell surface number and stability of 3-containing nicotinic acetylcholine receptors. The Journal of Neuroscience, 29 (21), 6883-6896. Roche. 2010. Roferon-A (Interferon alpha-2a). Australia. http://www.rocheaustralia.com/fmfiles/re7229005/downloads/anti-virals/roferon-pi.pdf. Roy R.K., Sapatnekar S.M., and Deshmukh R.A. (2005). The Effect of heat shock on production of recombinant human interferon alpha 2a by Escherichia coli. Iranian Biomedical Journal, 9 (4), 155-162. Samuel C.E. (2001). Antiviral actions of interferons. Clin Microbiol Rev., 14 (4), 778-809. Siregar M.L. (2009). Peningkatan mutu standar kualitas hasil cetakan mengunakan kombinasi pengaturan. Tesis. Fakultas Teknik Universitas Indonesia Press-Jakarta. Sørensen H.P., and Mortensen K.K. (2005). Advanced genetic strategies for recombinant expression in Escherichia coli. J Biotechnol, 115,113-128. Srivasta P., Bhattacharaya P., Pandey G., and Mukherjee K.J. (2005). Overexpression and purification of recombinant human interferon alpha2b in Escherichia coli. Protein Expr. Purif., 41, 313-322. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Studier F.W., Rosenberg A.H., Dunn J.J., Dubendorff J.W. (1990). Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymol., 185, 60-89. Sudjadi. (2008). Bioteknologi kesehatan. Penerbit Kanisius. Yogyakarta, Swaminathan,S. and Khanna,N. (1999). Affinity purification of recombinant interferon-alpha on a mimetic ligand adsorbent. Protein Expr. Purif., 15, 236-242. Tae-Ok Bae, Ho-Jin Chang, Jung Ho Kim, and Soon Jae Park. (1995). Purification and Characteization of Recombinant Human Interferon Alpha 2a Produced from Saccharomyces cerevisiae. J. Biochem. Mol. Biol., 28 (6), 477-483. Tan Hoan Tjay, dan Rahardja K. (2007). Obat-Obat Penting Khasiat, Penggunaan, dan Efek-Efek Sampingya. Jakata: PT Elex Media Komputindo. Tolia N. H., and Joshua-Tor L. (2006). Strategies for protein coexpression in Escherichia coli. Nature Methods, 3 (1), 55-64. Villaverde A., and Carrio M.M. (2003). Protein aggregation in recombinant bacteria: biological role of inclusion bodies. Biotechnol Lett., 25, 13851395. Xiaowei Li, Xin sui, Yan Zhang, Yepen Sun, Yan Zhao, Ying Zhai and Qingyu Wang. (2010). An improved calcium chloride method preparation and transformation of competent cells. African Journal of Biotechnology, 9 (50), 8549-8554. Yon J.M. (2002). Protein folding in the post-genomic era. J Cell Mol Med., 6, 307-327. Yu-Ling Sun, Yi-Juain Lin, and Chih-Sheng Lin. (2011). Soluble expression and production of rabbit neutrophil peptide-1 in Escherichia coli. Romanian Biotechnological Letters, 16 (5), 6618-6629. Zhang Y., Olsen D.R., Nguyen K.B., Olson P.S., Rhodes E.T. and Mascarenhas D. (1998). Expression of eukaryotic proteins in soluble form in E. coli. Protein Exp Purif., 12, 159-165. Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Lampiran 1. Skema Kerja Penelitian Penyiapan DNA insert Penyiapan vektor pET-32b(+) Amplifikasi dengan PCR Purifikasi Pemotongan Purifikasi fenol kloroform Pemotongan Purifikasi fenol kloroform Ligasi DNA insert dan vektor pET-32b(+) Transformasi ke E. coli DH5α (inang kloning) Skrining dan verifikasi klon transforman (PCR colony screening, isolasi vektor rekombinan, analisis restriksi, sekuensing) Kultivasi dan isolasi vektor rekombinan pET-32b(+)-IFN α2a Transformasi ke E. coli BL21(DE3) (inang ekspresi) Skrining dan verifikasi klon transforman (PCR colony screening) Kultivasi dan ekspresi sel transforman (induksi dengan IPTG) Isolasi protein rekombinan human IFN α2a Analisis elektroforesis SDS-PAGE (Western blot dan Pewarnaan CBB) Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Lampiran 2. Vektor pcDNA3.1+:IFN-Gene Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Lampiran 3. Hasil Sekuen Klon 2 E. coli DH5α dengan Stag-18mer-primer Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 “Lanjutan” Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Lampiran 4. Hasil Sekuen Klon 2 E. coli DH5α dengan T7terminator primer Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 ”Lanjutan” Kloning gen..., Arizah Kusumawati, F Farmasi, 2013 Lampiran 5. Nukleotida Human Interferon Alpha 2a (GenBank: DI084466.1) Kloning gen..., Arizah Kusumawati, F Farmasi, 2013