BAB I PENDAHULUAN A. Latar Belakang Penyakit
advertisement

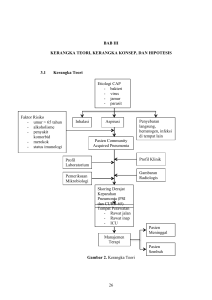
1 BAB I PENDAHULUAN A. Latar Belakang Penyakit kardiovaskular sebagai salah satu penyebab utama mortalitas lebih banyak diderita oleh subyek laki-laki daripada subyek perempuan, sehingga jenis kelamin laki-laki dikenal sebagai salah satu faktor risiko klasik penyakit kardiovaskular. Konsentrasi hormon androgen, khususnya testosteron, yang jauh lebih tinggi pada subyek laki-laki dibandingkan subyek perempuan menimbulkan dugaan adanya peran testosteron dalam penyakit kardiovaskular (Kaushik et al., 2010; Spoletini et al., 2011). Trombosit memainkan peran penting pada hemostasis dan penyakit kardiovaskular. Trombosit berperan pada hemostatis primer melalui pembentukan sumbat trombosit. Pada penyakit kardiovaskular, pada saat terjadi ruptur plak aterosklerotik, trombosit mengalami aktivasi, adhesi dan agregasi yang tidak semestinya (inappropiate), sehingga menyokong terbentuknya trombus (Nieswandt et al., 2011; Swieringa et al., 2014). Aktivasi trombosit melalui dua mekanisme, yaitu aktivasi tirosin kinase dan aktivasi G protein coupled receptors (GPCR). Tirosin kinase diaktivasi oleh kolagen subendotel, sedangkan GPCR diaktivasi oleh agonis trombosit soluble, seperti trombin, epinefrin, asam arakhidonat, adenosin diphosphate (ADP), dan tromboksan A2 (TXA2). Aktivasi trombosit menyebabkan peningkatan konsentrasi kalsium intratrombosit, yang selanjutnya memicu aktivasi glikoprotein (GP) 2 IIb/IIIa pada membran trombosit. GPIIb/IIIa yang teraktivasi menyebabkan agregasi trombosit dengan diperantarai fibrinogen (Broos et al., 2011). Penelitian terdahulu menunjukkan kecenderungan testosteron meningkatkan agregasi trombosit. Johnson et al. (1977) melaporkan agregasi trombosit tikus menurun dengan kastrasi, akan tetapi kembali meningkat dengan pre-treatment testosteron. Secara ex vivo, trombosit tikus yang diinkubasi dengan testosteron (110 ng/ml) mengalami peningkatan agregasi. Johnson et al. (1977) menunjukkan bahwa peningkatan agregasi trombosit oleh hormon androgen berkorelasi dengan tingkat androgenitas hormon tersebut. Pilo et al. (1981) melaporkan bahwa pemberian testosteron pada trombosit manusia dan tikus meningkatkan agregasi trombosit yang diinduksi ADP, epinefrin, kolagen, dan asam arakhidonat. Ditunjukkan pula paparan testosteron terhadap suspensi trombosit meningkatkan pelepasan TXA2. Rosenblum et al. (1987) melaporkan bahwa pemberian implan testosteron atau dihidrotestosteron (DHT) meningkatkan agregasi trombosit pada vena mesenterika tikus. Matsuda et al. (1994) menunjukkan testosteron meningkatkan densitas reseptor TXA2 pada trombosit manusia dan tikus. Ajayi dan Halushka (2005) melaporkan bahwa kastrasi pada penderita karsinoma prostat menurunkan densitas reseptor TXA2 dan menekan agregasi trombosit. Penelitian lain menunjukkan bahwa hormon androgen justru menghambat agregasi trombosit. ShiJun et al. (2007a, 2007b) melaporkan bahwa kastrasi meningkatkan pembentukan trombus. Pemberian DHT, yang merupakan androgen kuat turunan testosteron, mampu menghambat agregasi trombosit pada tikus. Penelitian terkini oleh Campelo et al. (2012) dan Cutini et al. (2012) 3 menunjukkan paparan testosteron pada kultur sel endotel aorta tikus menghambat agregasi trombosit melalui peningkatan produksi NO endotel. Sel endotel diketahui berperan menghambat aktivasi trombosit, antara lain melalui biosintesis dan pelepasan agen tromboregulator, seperti endotheliumderived relaxing factor (EDRF) atau nitric oxide (NO) dan prostasiklin (PGI2). PGI2 adalah antiagregator trombosit yang paling poten, selain sebagai vasodilator. PGI2 endotel akan berikatan dengan reseptor PGI2 (IP1) pada membran trombosit. Ligasi IP1-yang merupakan GPCR-oleh PGI2 menyebabkan aktivasi enzim adenilat siklase, yang mengkatalisis perubahan ATP menjadi cyclic adenosine monophosphate (cAMP), sehingga terjadi peningkatan cAMP intratrombosit, aktivasi protein kinase A (PKA), penurunan konsentrasi ion kalsium, dan akhirnya penghambatan agregasi trombosit (Broos et al., 2011; Feletou et al., 2011; Majed & Khalil, 2012). Sebagai prostaglandin (PG) turunan fosfolipid membran sel, biosintesis PGI2 memerlukan kerja beberapa enzim, yaitu enzim fosfolipase A2 (PLA2) yang mengkatalisis pemecahan fosfolipid membran sel menjadi asam arakhidonat, enzim siklooksigenase (COX) yang mengkatalisis konversi asam arakhidonat menjadi PGD2 dan PGH2, dan enzim PGI2 sintase (PGIS) yang mengkatalisis perubahan PGH2 menjadi PGI2. Terdapat dua isomer COX, yaitu COX-1 dan COX-2. COX-1 berperan pada sel endotel dalam keadaan basal atau istirahat. COX-2 berperan pada sel endotel dalam keadaan terstimulasi. Sintesis PGI2 memerlukan enzim COX-2, yang diinduksi shear stress, glukosa tinggi, hiperkolesterolemia, lipopolysaccharide (LPS), dan sitokin pro-inflamasi, seperti 4 interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), dan interferon-γ (IFN-γ) (Feletou et al., 2011; Majed & Khalil, 2012). Penelitian terbaru menunjukkan bahwa sel endotel mengandung reseptor androgen (AR). Sel endotel juga telah diketahui mengandung enzim 5α- reduktase, yang mengkatalisis perubahan testosteron (5-10% dari bioavailable testosteron) menjadi DHT, serta enzim aromatase, yang mengkatalisis perubahan testosteron (0,1-0,2% dari bioavailable testosteron) menjadi estradiol. Sel endotel juga diketahui mengandung reseptor estrogen (ER), baik ER-α dan ER-β. Dengan adanya AR, ER, dan enzim-enzim metabolisme testosteron, endotel merupakan salah satu sel target testosteron (Wu & von Eckardstein, 2003 cit. Kelly & Jones, 2013; Liu et al., 2003 cit. Torres-Estay et al., 2015). Dipertanyakan apakah testosteron mempengaruhi sintesis PGI2 pada sel endotel? Testosteron dilaporkan menurunkan sintesis PGI2 pada aorta yang diperfusi (Chang et al., 1982), akan tetapi pemaparan testosteron pada kultur sel endotel tidak mempengaruhi (Myers et al., 1995) atau meningkatkan (Seillan et al., 1983) sintesis PGI2. Diduga bahwa konversi testosteron menjadi estrogen berlangsung di dalam sel endotel, dan hal tersebut berkontribusi terhadap peningkatan sintesis PGI2 endotel. Telah dilaporkan bahwa ER, baik ER-α (Sobrino et al., 2010) ataupun ER-β (Garcia-Martinez et al., 2003), berperan meningkatkan sintesis PGI2 endotel yang diinduksi IL-1β. Sel endotel juga berperan menghambat adhesi trombosit. Lapisan endotelium yang intak melindungi trombosit dari paparan kolagen subendotel. Selain itu, sel endotel dalam keadaan tenang tidak atau sedikit mengekspresikan molekul adhesi. 5 Beberapa molekul adhesi ditengarai berperan dalam adhesi trombosit-endotel, antara lain selektin-platelet (selektin-P) dan intercellular adhesion molecule-1 (ICAM-1). Selektin-P diperlukan untuk rolling trombosit, sedangkan ICAM-1 diperlukan untuk firm adhesion trombosit dengan sel endotel (Palomo et al., 2012). Molekul cluster of differentiation-40 (CD40) adalah reseptor pada permukaan sel yang termasuk dalam keluarga TNF. CD40 pertama kali dikenali dan diketahui fungsinya pada limfosit B, sedangkan CD40 ligand (CD40L) diekspresikan pada sel limfosit T CD4+. Interaksi antara ligan dan reseptor tersebut (CD40L-CD40) berperan penting pada respon sel B yang tergantung sel T (T cell-dependent B cell responses). Saat ini diketahui bahwa CD40 diekspresikan secara luas pada monosit, sel dendritik, sel epitel, dan sel endotel, sedangkan CD40L diekspresikan pada sel mast, basofil, eosinofil, limfosit B, sel natural killer, monosit, makrofag, sel otot polos vaskular, sel endotel, dan trombosit. Dilaporkan bahwa ligasi CD40 endotel oleh CD40L trombosit akan menstimulasi sel endotel dan memicu peningkatan ekspresi ICAM-1 endotel (Li et al., 2009). Endotelium berhadapan langsung dengan darah, sehingga perubahan biokimiawi darah dapat mempengaruhi metablisme sel endotel. Pada kondisi normal konsentrasi glukosa darah 3,8-5,6 mM, akan tetapi dalam kondisi homeostasis terganggu konsentrasi glukosa darah dapat meningkat mencapai >10 mM (hiperglikemia). Lingkungan glukosa tinggi dapat mengganggu metabolisme dan homeostasis sel endotel (Popov, 2010). Cosentino et al. (2003) melaporkan bahwa glukosa tinggi (>10 nM) dapat memodulasi sel endotel berupa peningkatan 6 ekspresi COX-2 dan penurunan sintesis PGI2. Lingkungan glukosa tinggi juga dilaporkan meningkatan ekspresi molekul adhesi dan sitokin pro-inflamasi sel endotel (Basta et al., 2007). Hiperglikemia meupakan hallmark diabetes mellitus (DM), baik DM tipe 1 maupun DM tipe 2 (Ozougwu et al., 2013). Dilaporkan bahwa trombosit pada pasien DM, dengan adanya stimulus tertentu cenderung lebih reaktif, yaitu lebih mudah mengalami aktivasi dan agregasi (Kakouros et al., 2011). Disfungsi endotel (Basha et al., 2012) dan kelainan sinyal pada trombosit (Schneider, 2009) berkontribusi pada hiperreaktivitas trombosit pada penderita DM. Hal ini banyak berperan terhadap peningkatan terjadinya penyakit kardiovaskular pada penderita DM (Dokken et al., 2008). Beberapa penelitan terdahulu menunjukkan hasil yang berbeda mengenai pengaruh testosteron terhadap agregasi trombosit. Endotel-yang diketahui mempunyai kapasitas menghambat adhesi dan agregasi trombosit-merupakan merupakan jaringan target testosteron. Hal ini karena sel endotel mengandung AR, enzim-enzim yang memetabolisme testosteron, dan ER. Dengan demikian, menarik untuk ditelaah apakah testosteron menghambat agregasi trombosit melalui COX-2 dan ER-β endotel, serta menghambat potensi adhesi trombosit pada sel endotel, melalui CD40 dan ICAM-1 pada sel endotel. Aspek tersebut belum pernah ditinjau oleh peneliti lain. Pada posisi yang selalu terpapar aliran darah, endotel sebagai tromboregulator dapat dipengaruhi oleh kondisi hiperglikemia atau glukosa tinggi. Dengan demikian, menarik pula untuk dikaji bagaimana peran testosteron dalam adhesi dan agregasi trombosit melalui endotel 7 dalam perubahan kondisi lingkungan glukosa tinggi. Terdapat dua pendapat yang berseberangan mengenai peran testosteron dalam penyakit kardiovaskular. Pendapat pertama mengatakan testosteron tidak bersifat kardioprotektif karena testosteron dalam dosis suprafisiologis menimbulkan efek merugikan bagi tubuh. Pendapat konsentrasi kedua mengatakan testosteron bersifat kardioprotektif testosteron yang rendah merupakan faktor risiko karena penyakit kardiovaskular (Cheung et al., 2015; Torres-Estay et al., 2015). B. Rumusan Masalah Berdasarkan latar belakang di atas, dapat dijumput hal-hal yang sekiranya dapat menjadi dasar untuk menentukan rumusan masalah, yaitu: 1. Trombosit mempunyai peran penting dalam penyakit kardiovaskular melalui aktivasi, adhesi dan agregasi. 2. Trombosit teraktivasi oleh agonis trombosit, seperti kolagen, epinefrin, asam arakhidonat, ADP, TXA2, dan trombin, sehingga terjadi perubahan metabolisme trombosit menuju jalur bersama, yaitu peningkatan konsentrasi kalsium intratrombosit. 3. Dengan peningkatan konsentrasi kalsium terjadi aktivasi molekul GP pada membran trombosit, dan dengan adanya fibrinogen maka terjadi pelekatan antartrombosit yang teraktivasi (agregasi trombosit). 4. Sel endotel berperan menghambat agregasi trombosit melalui sintesis agen tromboregulator, seperti NO dan PGI2. 8 5. PGI2 menghambat aktivasi trombosit melalui aktivasi adenilat siklase, peningkatan cAMP, aktivasi PKA, serta penurunan konsentrasi kalsium intratrombosit. 6. Biosintesis PGI2 oleh sel endotel memerlukan enzim COX-2. 7. Trombosit melekat (adhesi) pada sel endotel yang teraktivasi, antara lain melalui ICAM-1. 8. CD40 pada sel endotel berperan meningkatkan ekspresi ICAM-1 endotel. 9. Penyakit kardiovaskular lebih sering didapatkan pada subyek laki-laki daripada perempuan, yang diduga terkait dengan hormon androgen, khususnya testosteron. 10. Beberapa penelitian menunjukkan testosteron meningkatkan agregasi trombosit, akan tetapi penelitian yang lain mengindikasikan sebaliknya. 11. Sel endotel merupakan jaringan target testosteron karena mengandung AR dan enzim-enzim metabolisme testosteron. 12. Sebagian testosteron bebas yang masuk ke dalam sel endotel dikonversi menjadi estradiol oleh aromatase; sel endotel diketahui mengandung ER. 13. Testosteron diduga mempengaruhi potensi sel endotel dalam menghambat adhesi dan agregasi trombosit setelah dikonversi menjadi estrogen. 14. Glukosa tinggi memodulasi fungsi endotel, antara lain meningkatkan ekspresi COX-2, menurunkan sintesis PGI2, dan meningkatkan agregasi trombosit. 15. Glukosa tinggi juga dilaporkan meningkatkan ekspresi ICAM-1 endotel. 9 Berdasarkan hal-hal tersebut di atas, maka diajukan pertanyaan penelitian sebagai berikut. 1. Apakah testosteron bersifat kardioprotektif dengan menghambat agregasi trombosit melalui endotel? 2. Apakah testosteron bersifat kardioprotektif dengan menghambat potensi adhesi trombosit pada sel endotel? Adapun rincian sub masalah penelitian adalah sebagai berikut. 1.1 Apakah persentase agregasi trombosit pada suspensi trombosit (platelet rich plasma; PRP) yang diinkubasi dengan sel endotel (human umbilical vein endothelial cell culture; HUVEC) yang terpapar testosteron lebih rendah daripada persentase agregasi trombosit pada PRP yang diinkubasi dengan HUVEC yang tidak terpapar testosteron? 1.2 Apakah konsentrasi cAMP trombosit pada PRP yang diinkubasi dengan HUVEC yang terpapar testosteron lebih tinggi daripada konsentrasi cAMP trombosit pada PRP yang diinkubasi dengan HUVEC yang tidak terpapar testosteron? 1.3 Apakah ekspresi COX-2 pada HUVEC yang terpapar testosteron lebih tinggi daripada ekspresi COX-2 pada HUVEC yang tidak terpapar testosteron? 1.4 Apakah ekspresi ER-β pada HUVEC yang terpapar testosteron lebih tinggi daripada ekspresi ER-β pada HUVEC yang tidak terpapar testosteron? 1.5 Apakah persentase agregasi trombosit pada PRP yang diinkubasi dengan HUVEC yang terpapar testosteron dan glukosa tinggi lebih tinggi daripada 10 persentase agregasi trombosit pada PRP yang diinkubasi dengan HUVEC yang terpapar glukosa tinggi saja? 1.6 Apakah konsentrasi cAMP trombosit pada PRP yang diinkubasi dengan HUVEC yang terpapar testosteron dan glukosa tinggi lebih rendah daripada konsentrasi cAMP trombosit pada PRP yang diinkubasi dengan HUVEC yang terpapar glukosa tinggi saja? 1.7 Apakah ekspresi COX-2 pada HUVEC yang terpapar testosteron dan glukosa tinggi lebih rendah daripada ekspresi COX-2 pada HUVEC yang terpapar glukosa tinggi saja? 1.8 Apakah ekspresi ER-β pada HUVEC yang terpapar testosteron dan glukosa tinggi lebih rendah daripada ekspresi ER-β pada HUVEC yang terpapar glukosa tinggi saja? 2.1 Apakah ekspresi ICAM-1 pada HUVEC yang terpapar testosteron lebih rendah daripada ekspresi ICAM-1 pada HUVEC yang tidak terpapar testosteron? 2.2 Apakah ekspresi CD40 pada HUVEC yang terpapar testosteron lebih rendah daripada ekspresi CD40 pada HUVEC yang tidak terpapar testosteron? 2.3 Apakah ekspresi ICAM-1 pada HUVEC yang terpapar testosteron dan glukosa tinggi lebih tinggi daripada ekspresi ICAM-1 pada HUVEC yang terpapar glukosa tinggi saja? 11 2.4 Apakah ekspresi CD40 pada HUVEC yang terpapar testosteron dan glukosa tinggi lebih tinggi daripada ekspresi CD40 pada HUVEC yang terpapar glukosa tinggi saja? C. Tujuan Penelitian C.1 Tujuan umum Untuk mengkaji peran testosteron sebagai kardioprotektor dengan menghambat agregasi dan potensi adhesi trombosit melalui endotel. C.2 Tujuan khusus 1. Untuk mengukur persentase agregasi trombosit pada suspensi trombosit yang diinkubasi dengan sel endotel yang telah terpapar testosteron dengan atau tanpa glukosa tinggi. 2. Untuk mengukur konsentrasi cAMP trombosit pada suspensi trombosit yang diinkubasi dengan sel endotel yang telah terpapar testosteron dengan atau tanpa glukosa tinggi, serta korelasinya dengan persentase agregasi trombosit. 3. Untuk mengukur ekspresi COX-2 pada sel endotel yang terpapar testosteron dengan atau tanpa glukosa tinggi, serta korelasinya dengan persentase agregasi trombosit dan konsentrasi cAMP trombosit. 4. Untuk mengukur ekspresi ER-β pada sel endotel yang terpapar testosteron dengan atau tanpa glukosa tinggi, serta korelasinya dengan ekspresi COX2 endotel dan persentase agregasi trombosit. 12 5. Untuk mengukur ekspresi ICAM-1 pada sel endotel yang terpapar testosteron dengan atau tanpa glukosa tinggi. 6. Untuk mengukur ekspresi CD40 pada sel endotel yang terpapar testosteron dengan atau tanpa glukosa tinggi, serta korelasinya dengan ekspresi ICAM-1 endotel. D. Manfaat Penelitian D.1 Manfaat teoretis Memberi sumbangan pengetahuan mengenai peran testosteron dalam adhesi dan agregasi trombosit melalui endotel. D.2 Manfaat praktis 1. Memberi landasan ilmiah kemungkinan pemberian testosteron atau obat antagonis AR untuk mencegah adhesi dan agregasi trombosit. 2. Memberi landasan ilmiah kemungkinan pemberian testosteron dengan tujuan untuk memperbaiki kesehatan kardiovaskular bagi subyek laki-laki usia lanjut, subyek dengan hipogonadisme, atau subyek dengan penyakit kardiovaskular. E. Keaslian Penelitian E.1 Peran testosteron dalam agregasi trombosit yang dimediasi oleh endotel Peran sel endotel dalam menghambat agregasi trombosit telah dikaji oleh Nordoy et al. (1978), Lagarde et al. (1984), dan Altorjay et al. (1989) secara in vitro. Penelitian tersebut sebagian besar menggunakan HUVEC, dengan agonis 13 trombosit seperti trombin, asam arakhidonat, ADP, dan kolagen. Parameter yang diukur adalah persentase agregasi trombosit, tromboksan B2 (TXB2), dan cAMP trombosit. Peneliti tersebut tidak mengkaji peran testosteron dalam agregasi trombosit melalui endotel. Peran testosteron dalam agregasi trombosit telah dikaji oleh Pilo et al. (1981) secara in vitro dengan memberikan testosteron pada PRP manusia atau tikus coba. Parameter yang diukur adalah agregasi trombosit, TXA2, dan produk lipolisis. Pilo et al. tidak menambahkan endotel dalam penelitian tersebut. Peneliti yang lain juga mengkaji masalah tersebut dengan pendekatan in vivo dan in vitro sekaligus. Johnson et al. (1977) memberikan testosteron pada hewan coba yang telah dikastrasi serta DHT, testosteron, metiltestosteron, androstenedion, androsteron, dan estradiol secara in vitro pada suspensi trombosit hewan coba. Parameter yang diukur adalah agregasi trombosit. Johnson et al. (1977) menambahkan flutamide untuk mengkaji peran AR. Sementara itu, Rosenblum et al. (1987) mengkaji pengaruh implan testosteron dan DHT terhadap agregasi trombosit pada tikus coba. Matsuda et al. (1994) memberikan testosteron pada hewan coba yang dikastrasi maupun yang tidak dikastrasi. Parameter yang diukur antara lain densitas reseptor TXA2 pada trombosit dan aorta, kontraktilitas aorta, dan agregasi trombosit. ShiJun et al. (2007a) memberikan DHT tanpa atau dengan flutamide pada hewan coba sebelum dilakukan kastrasi. Parameter yang diukur adalah trombus yang terbentuk in vivo dan rasio TXB2 terhadap 6-keto-PGF1α dalam plasma pasca induksi FeCl3. Secara ex vivo ShiJun et al. (2007a) menambahkan DHT dengan atau tanpa flutamide pada PRP tikus yang dikastrasi. 14 Parameter yang diukur adalah persentase agregasi trombosit dan adhesi trombosit. ShiJun et al. (2007a) tidak mengkaji peran testosteron, estrogen, ER, dan aromatase. Penelitian tersebut belum mengkaji secara jelas peran testosteron dalam agregasi trombosit melalui endotel, konversi testosteron menjadi estrogen, dan ER. Zerr-Fouineau et al. (2007) mengkaji peran hormon steroid kelamin dalam agregasi trombosit melalui sel endotel, akan tetapi hormon yang dikaji adalah progesteron dan dalam kaitannya dengan kapasitas sel endotel mensintesis NO. Penelitian terbaru dari Campelo et al. (2012) mengkaji pengaruh paparan testosteron pada kultur sel endotel tikus terhadap agregasi trombosit, akan tetapi dilakukan pada kondisi lingkungan normoglikemia saja, serta melalui mekanisme/ jalur NO. Peneliti-peneliti tersebut di atas belum mengkaji peran testosteron dan metabolismenya dalam sel endotel dalam hubungannya dengan kapasitas sel endotel mensintesis tromboregulator PGI2. Peran testosteron dalam sintesis PGI2 telah dikaji oleh Seillan et al. (1983) dan Myers et al. (1995) pada sel endotel, Nakao et al. (1981) dan Wakasugi et al. (1989) pada sel otot polos vaskular, dan Chang et al. (1982) pada perfusi aorta. Peran testosteron dalam ekspresi COX-2 telah dikaji oleh Razmara et al. (2005) pada pembuluh darah serebral tikus coba pasca injeksi LPS. Sementara, Osterlund et al. (2010) mengkaji peran DHT dalam ekspresi COX-2 pada kultur otot polos arteri koronaria manusia. Belum ada peneliti yang mengkaji peran testosteron dalam ekspresi COX-2 endotel dan pengaruhnya terhadap agregasi trombosit. Dietrich et al. (2011) telah mengkaji peran testosteron dan konversi lokal menjadi estrogen dalam proliferasi sel endotel. Parameter yang diukur adalah 15 ekspresi aromatase, AR, dan ER pada sel endotel. Peran testosteron dalam agregasi trombosit melalui sel endotel tidak dikaji oleh peneliti tersebut. Penelitian ini mengkaji peran testosteron sebagai kardioprotektor dengan menghambat agregasi trombosit melalui kultur sel endotel manusia (HUVEC) dalam lingkungan glukosa normal atau glukosa tinggi. Parameter yang diukur adalah persentase agregasi trombosit, konsentrasi cAMP trombosit, ekspresi COX-2 dan ER-β endotel. E.2 Peran testosteron dalam potensi adhesi trombosit yang dimediasi oleh endotel Adhesi trombosit pada monolayer kultur sel endotel telah dikaji oleh Czervionke et al. (1978) dan Yagyu et al. (1991). Penelitian tersebut memperhatikan faktor-faktor aktivasi trombosit, aktivasi endotel, sintesis dan pelepasan prostasiklin oleh sel endotel. Akan tetapi, peran testosteron dalam adhesi trombosit melalui ekspresi molekul adhesi endotel belum dikaji dalam penelitian tersebut. Beberapa peneliti telah mengkaji peran testosteron (Hatakeyama et al., 2002; Mukherjee et al., 2002) dan DHT (McCrohon et al., 1999; Death et al., 2004; Norata et al., 2006) dalam ekspresi molekul adhesi pada sel endotel yang distimulasi sitokin atau LPS, melalui AR, ER, dan konversi testosteron menjadi estrogen. Molekul adhesi yang diukur adalah vascular cell-adhesion molecule-1 (VCAM-1), yang berperan dalam adhesi lekosit. Terhadap ekspresi CD40 endotel, telah dilaporkan pengaruh hormon steroid kelamin, seperti DHT (Norata et al., 2006), estradiol (Geraldes et al., 2006), atau dehidroepiandrosteron (DHEA) (Li et 16 al., 2009). Peran hormon testosteron terhadap ekspresi CD40 endotel belum diketahui. Penelitian ini mengkaji peran testosteron sebagai kardioprotektor dengan menghambat potensi adhesi trombosit pada sel endotel manusia (HUVEC) dalam lingkungan glukosa normal atau glukosa tinggi. Parameter yang diukur adalah ekspresi ICAM-1 dan CD40 endotel. Beberapa penelitian terdahulu terkait dengan keaslian topik penelitian tercantum dalam Lampiran Tabel 1.1. 361 22-25 Oktober 2014 28-30 Oktober 2014 Presentasi oral, Ikhlas M.Jenie, Soedjono Aswin, Budi Mulyono, Sri Kadarsih Soejono, “The Comparison of Platelet Aggregation in the Presence of Disperse Primary and Monolayer Secondary HUVEC Exposed to Testosterone in High Glucose Medium”, Joint International Conference 2014 APCHIERGOFUTURE-PEI-IAIFI, Universitas Udayana, Denpasar. Presentasi oral, Ikhlas M. Jenie, Sri Kadarsih Soejono, Soedjono Aswin, Budi Mulyono, “Platelet Aggregation after Exposure of Testosterone and High Glucose to Human Umbilical Vein Endothelial Cell Culture”, The 6th Indonesia Japan Joint Scientific Symposium, Universitas Gadjah Mada, Yogyakarta. PENGABDIAN 2010 2012 Penyuluhan mengenai Pemanfaatan Seduhan Kelopak Bunga Rosela Kering sebagai Obat Antihipertensi dalam Rangka Muktamar Seabad Muhammadiyah (FKIK UMY) Deteksi dan Pemantauan Tekanan Darah Tinggi pada Lansia di Posyandu Keji Beling, Kelurahan Keparakan, Yogyakarta (LP3M UMY) KEMASYARAKATAN 2007 – sekarang 2008 – 2012 2011 – sekarang Seksi Kesehatan, Takmir Masjid Darussalam, Pujokusuman Yogyakarta. Bendahara, RW 06 Kelurahan Keparakan Kota Yogyakarta. Anggota, Pimpinan Ranting Muhammadiyah Pujokusuman, Yogyakarta. Demikianlah daftar riwayat hidup ini saya buat dengan sebenarnya. Yogyakarta, 2 September 2015 Ikhlas Muhammad Jenie