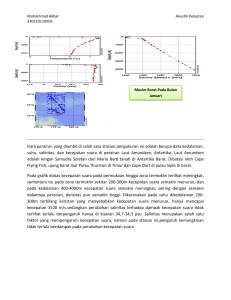
- Balai Riset dan Observasi Laut
advertisement

Vol. 2, No. 2, Desember 2010 ISSN Cetak : 2087-9423 ISSN Electronik: 2085-6695 JURNAL ILMU DAN TEKNOLOGI KELAUTAN TROPIS Diterbitkan Oleh: Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan Fakultas Perikanan dan Ilmu Kelautan Institut Pertanian Bogor http://www.itk.fpik.ipb.ac.id/ej_itkt22 Jurnal Ilmu dan Teknologi Kelautan Tropis Jurnal Ilmu dan Teknologi Kelautan Tropis merupakan pengembangan dari E-Jurnal Ilmu dan Teknologi Kelautan Tropis yang mulai terbit semenjak Juni 2009, mulai Desember 2010 diterbitkan secara cetak dan elektronik. Jurnal ini merupakan jurnal ilmiah dibidang ilmu dan teknologi kelautan tropis dan diterbitkan secara berkala sebanyak dua kali dalam setahun. Redaksi menerima paper dalam bahasa Indonesia atau bahasa Inggris yang diketik di atas kertas A4 dalam 1 spasi, Times New Roman, Font 12, termasuk gambar dan tabel dan dikirim melalui email ke alamat redaksi. Pedoman penulisan paper dapat dilihat pada bagian belakang jurnal ini. Diterbitkan oleh : Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Penanggung Jawab : Ketua Ikatan Sarjana Oseanologi Indonesia dan Ketua Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Pemimpin Redaksi : Bisman Nababan, Ph.D. Penyunting Ahli (Mitra Bebestari): Augy Syahailatua, Ph.D. (Marine Fisheries and Marine Biology, P2O-LIPI) Bambang Yulianto, Ph.D. (Marine Ecotoxicology, FPIK-UNDIP) Prof. Dr. Bonar Pasaribu (Marine Acoustic, ITK-IPB) Prof. Dr. Feliatra (Marine Microbiology, ITK-UNRI) Dwi Djoko Eny Setyono, Ph.D. (Marine Biology and Aquaculture, P2O-LIPI) Prof. Dr. Inneke Rumengan (Marine Biotechnology, FPIK-UNSRAT) Iskaq Iskandar, Ph.D. (Physical Oceanography, NOAA-PMEL, USA) Iqbal Djawad, Ph.D. (Marine Aquaculture & Physiology., FIKP-UNHAS) John I. Pariwono, Ph.D. (Physical Oceanography, ITK-IPB) Joko Santoso, Ph.D. (Aquatic Product Processing Technology, THP-IPB) Dr. Jonson L. Gaol (Marine Remote Sensing & GIS, ITK- IPB) Prof. Dr. Mulia Purba (Physical Oceanography, ITK-IPB) Neviaty P. Zamani, Ph.D. (Marine Biology, Coral Reef, ITK-IPB) Suhartati M. Natsir, Ph.D. (Marine Ecology, P2O-LIPI) Wahyu Pandoe, Ph.D. (Physical Oceanography, Modeling, BPPT) Zainal Arifin, Ph.D. (Marine Pollution, Chemical Oceanography, P2O-LIPI) Tri Prartono, Ph.D. (Chemical Oceanography, ITK-IPB) Penyunting Pelaksana: Sri Ratih Deswati, M.Si., Meutia Samira Ismet, M.Si., Jafar Elly, M.Si., Muhammad Subhan, S.Pi., dan Sahat M.R. Tampubolon, Amd. Alamat Redaksi : Departemen Ilmu dan Teknologi Kelautan, FPIK Jl. Lingkar Akademik, Kampus IPB Darmaga, Bogor 16680 e-mail: [email protected] Isi dapat dikutip dengan menyebutkan sumbernya ___________________________________________________________________ KATA SAMBUTAN Saya sebagai Ketua Umum Ikatan Sarjana Oseanologi Indonesia (ISOI) sangat senang dan bangga atas terbitnya Jurnal Ilmu dan Teknologi Kelautan Tropis (ITKT) Volume 2 Nomor 2 Desember 2010 ini yang merupakan pengembangan dari E-Jurnal Ilmu dan Teknologi Kelautan Tropis yang sudah terbit semenjak Juni 2009. Dengan terlaksananya kerjasama antara ISOI dengan Departemen Ilmu dan Teknologi Kelautan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor maka Jurnal ITKT dapat diterbitkan dan menjadi jurnal cetak sekaligus jurnal elektronik serta diharapkan menjadi jurnal andalan ISOI. Seperti disepakati bersama, Jurnal ITKT akan terbit dua kali dalam setahun yaitu pada bulan Juni dan Desember setiap tahun. Dengan manajemen yang baik, diharapkan Jurnal ITKT ini dapat terbit tepat waktu. Melalui mitra bebestari yang professional dan ahli dalam bidangnya diharapkan mutu artikel (paper) yang terbit dalam Jurnal ITKT ini memiliki mutu ilmiah yang tinggi dan dapat dipertanggung jawabkan. Semoga dalam waktu dekat, Jurnal ITKT ini dapat memperoleh akreditasi dari Kementerian Pendidikan Nasional dan Lembaga Ilmu Pengetahuan Indonesia (LIPI) untuk menjaga dan meningkatkan mutu ilmiah dari jurnal ini. Ucapan terima kasih secara khusus saya sampaikan kepada Kementerian Pendidikan Nasional yang telah bersedia menyediakan dana untuk penerbitan Jurnal ini. Penghargaan sebesar-besarnya juga saya sampaikan kepada Pemimpin Redaksi, Mitra Bebestari, dan Penyunting Pelaksana yang telah bekerja keras untuk dapat menyelesaikan proses penerbitan Jurnal ini. Akhir kata, saya sebagai Ketua Umum ISOI mengucapkan selamat atas terbitnya Jurnal ITKT Volume 2 Nomor 2 Desember 2010 ini. Semoga Jurnal ITKT ini dapat menjadi rujukan utama dalam bidang ilmu dan teknologi kelautan serta bidang perikanan tropis di Indonesia maupun di dunia. Jakarta, Desember 2010 Prof. Dr. Indroyono Susilo Ketua Umum ISOI KATA SAMBUTAN Puji Syukur kehadirat Tuhan Yang Maha Kuasa atas bimbingan-Nya sehingga Jurnal Ilmu dan Teknologi Kelautan Tropis (ITKT) Volume 2 Nomor 2 Desember 2010 dapat terbit. Jurnal ini merupakan pengembangan dari E-Jurnal Ilmu dan Teknologi Kelautan Tropis yang sudah terbit semenjak Juni 2009 melalui kerjasama antara Ikatan Sarjana Oseanologi Indonesia (ISOI) dengan Departemen Ilmu dan Teknologi Kelautan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Seperti E-Jurnal ITKT, Jurnal ITKT akan terbit dua kali dalam setahun yaitu pada bulan Juni dan Desember setiap tahun dan diharapkan dapat terbit tepat waktu. Dengan mitra bebestari yang professional dan ahli dalam bidangnya, diharapkan mutu paper dalam Jurnal ITKT ini dapat ditingkatkan dan diharapkan dapat memperoleh akreditasi dari Kementerian Pendidikan Nasional dan Lembaga Ilmu Pengetahuan Indonesia (LIPI). Saya mengucapkan terima kasih banyak kepada Kementerian Pendidikan Nasional yang telah bersedia menyediakan dana untuk penerbitan Jurnal ini. Begitu juga kepada Pemimpin Redaksi, Mitra Bebestari, dan Penyunting Pelaksana yang telah bekerja keras untuk dapat menyelesaikan proses penerbitan Jurnal ini. Akhir kata, saya mengucapkan selamat atas terbitnya Jurnal ITKT ini dan semoga Jurnal ini dapat digunakan sebagai media diseminasi hasil-hasil penelitian dalam bidang ilmu dan teknologi kelautan serta bidang perikanan tropis di Indonesia maupun di dunia. Bogor, Desember 2010 Prof. Dr. Ir. Setyo Budi Susilo, M.Sc. Ketua Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB KATA PENGANTAR Puji syukur kehadirat Tuhan Yang Maha Kuasa atas rahmat-Nya sehingga Jurnal Ilmu dan Teknologi Kelautan Tropis Volume 2 Nomor 2 ini dapat terbit. Melalui seleksi dan review oleh tim reviewer (mitra bebestari), dari sejumlah paper yang masuk sebanyak 11 (sebelas) paper dapat diterima dengan perbaikan untuk diterbitkan pada edisi ini dengan judul sebagai berikut: (1) Pengaruh calsium dan fosfor terhadap pertumbuhan, efisiensi pakan, kandungan mineral dan komposisi tubuh juvenil ikan kerapu macan (epinephelus fuscoguttatus); (2) Identifikasi jenis ikan anemon (amphiprioninae) dan anemon simbionnya di kepulauan spermonde, sulawesi selatan; (3) Keragaan reproduksi ikan kerapu bebek (cromileptes altivelis) dari alam (f-0), induk generasi pertama (f-1), dan induk generasi ke dua (f-2); (4) Pemeliharaan gelondongan kerapu sunu (plectropomus leopardus) dengan persentase pergantian air yang berbeda; (5) Karakteristik senyawa bioaktif bakteri simbion moluska dengan gc-ms; (6) Akumulasi logam berat pb, cu, dan zn di hutan mangrove muara angke, jakarta utara; (7) Kajian stabilitas empat tipe kasko kapal pole and line; (8) Struktur komunitas ikan pada padang lamun yang berbeda di pulau barrang lompo; (9) Distribusi foraminifera bentik resen di laut Arafura; (10) Perencanaan waktu tetas telur ikan kerapu dengan penggunaan suhu inkubasi yang berbeda; dan (11) Massa air subtropical di perairan hamahera. Semoga Jurnal Ilmu dan Teknologi Kelautan Tropis ini dapat meningkatkan diseminasi dan ketersediaan informasi akan hasil-hasil penelitian di bidang ilmu dan teknologi kelautan serta perikanan tropis. Pemimpin Redaksi ISSN Cetak : 2087-9423 ISSN Elektronik : 2085-6695 Jurnal Ilmu dan Teknologi Kelautan Tropis Vol. 2, No. 2, Desember 2010 DAFTAR ISI PENGARUH CALSIUM DAN FOSFOR TERHADAP PERTUMBUHAN, EFISIENSI PAKAN, KANDUNGAN MINERAL DAN KOMPOSISI TUBUH JUVENIL IKAN KERAPU MACAN (Epinephelus fuscoguttatus) (THE EFFECT OF CALCIUM AND PHOSPHOROUS ON GROWTH, FEED EFFICIENCY, MINERAL CONTENT AND BODY COMPOSITION OF BROWN MARBLED GROUPER (Epinephelus fuscoguttatus) JUVENILE) Zainuddin ............................................................................................................................................................................... 1 IDENTIFIKASI JENIS IKAN ANEMON (Amphiprioninae) DAN ANEMON SIMBIONNYA DI KEPULAUAN SPERMONDE, SULAWESI SELATAN (IDENTIFICATION OF ANEMONEFISHES (Amphiprioninae) AND THEIR SIMBIONT IN SPERMONDE ARCHIPELAGO, SOUTH SULAWESI) Inayah Yasir, Syafiuddin, dan Sumarjito ............................................................................................................................ 10 KERAGAAN REPRODUKSI IKAN KERAPU BEBEK (Cromileptes altivelis) DARI ALAM (F-0), INDUK GENERASI PERTAMA (F-1), DAN INDUK GENERASI KE DUA (F-2) (REPRODUCTION PERFORMANCE OF HUMPBACK GROUPER (Cromileptes altivelis) FROM WILD BROODSTOCK (F-0), FIRST GENERATION BROODSTOCK (F-1), AND SECOND GENERATION BROODSTOCK (F2)) Tridjoko .................................................................................................................................................................................. 17 PEMELIHARAAN GELONDONGAN KERAPU SUNU (Plectropomus leopardus) DENGAN PERSENTASE PERGANTIAN AIR YANG BERBEDA (DIFFERENT PERCENTAGE OF WATER EXCHANGE ON GROWTH OF CORAL TROUT GROUPER FINGERLING (Plectropomus leopardus)) Titiek Aslianti ......................................................................................................................................................................... 26 KARAKTERISTIK SENYAWA BIOAKTIF BAKTERI SIMBION MOLUSKA DENGAN GC-MS (CHARACTERISTIC BIOACTIVE COMPOUND OF THE MOLLUSC SYMBIOTIC BACTERIA BY USING GC-MC) Delianis Pringgenies ............................................................................................................................................................... 34 AKUMULASI LOGAM BERAT PB, CU, DAN ZN DI HUTAN MANGROVE MUARA ANGKE, JAKARTA UTARA (ACCUMULATION OF HEAVY METALS PB, CU, AND ZN IN THE MANGROVE FOREST OF MUARA ANGKE, NORTH JAKARTA) Faisal Hamzah dan Agus Setiawan ....................................................................................................................................... 41 KAJIAN STABILITAS EMPAT TIPE KASKO KAPAL POLE AND LINE (STABILITY ANALYSIS OF FOUR TYPES OF POLE AND LINER) St. Aisyah Farhum ................................................................................................................................................................. 53 STRUKTUR KOMUNITAS IKAN PADA PADANG LAMUN YANG BERBEDA DI PULAU BARRANG LOMPO (FISH COMMUNITY STRUCTURE IN DIFFERENT SEAGRASS BEDS OF BARRANG LOMPO ISLAND) Rohani Ambo Rappe .............................................................................................................................................................. 62 DISTRIBUSI FORAMINIFERA BENTIK RESEN DI LAUT ARAFURA (THE DISTRIBUTION OF RECENT BENTHIC FORAMINIFERA IN THE ARAFURA SEA) Suhartati M. Natsir dan Rubiman ........................................................................................................................................ 74 PERENCANAAN WAKTU TETAS TELUR IKAN KERAPU DENGAN PENGGUNAAN SUHU INKUBASI YANG BERBEDA (PLANNING ON HATCHING TIME OF GROUPER EGGS THROUGH DIFFERENT INCUBATION TEMPERATURES) Regina Melianawati, Philip Teguh Imanto, dan Made Suastika ..................................................................................... 83 MASSA AIR SUBTROPICAL DI PERAIRAN HALMAHERA (SUBTROPICAL WATER MASSES IN HALMAHERA WATERS) Hadikusumah ......................................................................................................................................................................... 92 Pedoman Penulisan Naskah Artikel ..................................................................................................................................... 109 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 1-9, Desember 2010 PENGARUH CALSIUM DAN FOSFOR TERHADAP PERTUMBUHAN, EFISIENSI PAKAN, KANDUNGAN MINERAL DAN KOMPOSISI TUBUH JUVENIL IKAN KERAPU MACAN (Epinephelus fuscoguttatus) THE EFFECT OF CALCIUM AND PHOSPHOROUS ON GROWTH, FEED EFFICIENCY, MINERAL CONTENT AND BODY COMPOSITION OF BROWN MARBLED GROUPER (Epinephelus fuscoguttatus) JUVENILE Zainuddin Fakultas Ilmu Kelautan dan Perikanan Universitas Hasanuddin Email: [email protected] ABSTRACT The objectives of this study were to know concentration of calcium (Ca) and posphorus (P) in feed for growth, feed efficiency, proximate composition of the body and mineral content of brown marbled grouper juvenile. The study was conducted in the Center for Brackiswater Aquaculture Development, Takalar with randomized completed design 6 x 3 with the treatment of Ca and P supplement in feed i.e., (A) the supplement of 0 g/kg Ca and 0 g/kg P, (B) the supplement of 6 g/kg Ca and 0 g/kg P, (C) the supplement of 0 g/kg Ca and 6 g/kg P, (D) the supplement of 6 g/kg Ca and 6 g/kg P, (E) the supplement of 12 g/kg Ca and 6 g/kg P, and (F) the supplement of 18 g/kg Ca and 6 g/kg P. The result showed that P supplement with doses of 6 g/kg and Ca of 0 g/kg in feed are significantly affects on relative growth, feed efficiency, proximate composition and mineral content of brown marbled grouper juvenile. Keywords: growth, feed efficiency, proximate composition, brown marbled grouper juvenile ABSTRAK Penelitian ini bertujuan untuk mengetahui konsentrasi calcium (Ca) dan fosfor (P) dalam pakan terhadap pertumbuhan, efisiensi pakan, komposisi proksimat dan kandungan mineral tubuh juvenil ikan kerapu macan. Penelitian dilakukan di Balai Budidaya Air Payau Takalar. Penelitian dirancang dengan Rancangan Acak Lengkap 6 x 3 dengan perlakuan penambahan Ca dan P dalam pakan. Perlakuan A : penambahan Ca 0 g/kg pakan dan P 0 g/kg pakan, perlakuan B : penambahan Ca 6 g/kg pakan dan P 0 g/kg pakan, perlakuan C : penambahan Ca 0 g/kg pakan dan P 6 g/kg pakan, perlakuan D : penambahan Ca 6 g/kg pakan dan P 6 g/kg pakan, perlakuan E : penambahan Ca 12 g/kg pakan dan P 6 g/kg pakan dan perlakuan F : penambahan Ca 18 g/kg pakan dan P 6 g/kg pakan. Hasil penelitian menunjukkan bahwa penambahan P sebesar 6 g/kg dan 0 g/kg ke dalam pakan secara nyata berpengaruh terhadap pertumbuhan relatif, efisiensi pakan, kompisisi prosimat dan kandungan mineral tubuh juvenil ikan kerapu macan. Kata Kunci: pertumbuhan, efisiensi pakan, komposisi proksimat, juvenile ikan kerapu macan ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 1 Pengaruh Calsium dan Fosfor Terhadap Pertumbuhan, Efisiensi Pakan... I. PENDAHULUAN Calcium(Ca) dan phosphor (P) merupakan makro mineral yang berhubungan langsung dengan perkembangan dan pemeliharaan sistem skeleton serta berpartisipasi dalam berbagai proses fisiologis tubuh organisme. Kebutuhan Ca pada ikan dipengaruhi oleh kimia air, level P dalam pakan dan spesies (Lall, 2002). Hossain dan Furuichi (2000a, b, c) melaporkan bahwa penambahan Ca dalam pakan sangat diperlukan untuk pertumbuhan ikan belanak merah, Japanese flounder dan ikan scorpion. Pada kebanyakan spesies ikan, defisiensi P berakibat pada pertumbuhan yang lambat, efisiensi pakan yang jelek, mineralisasi tulang yang buruk, kandungan lipid tubuh yang tinggi serta kadar abu yang rendah (Tacon, 1992; Zainuddin et al., 2000; dan Lall, 2002). Ca dan P merupakan mineral yang saling sinergis (Zainuddin, 2001, 2004a) dan dalam bentuk hydroxyapatite dalam membentuk kristal-kristal tulang (Ye et al., 2006). Beberapa spesies ikan kerapu sangat potensial dibudidayakan karena pertumbuhannya cepat, konversi pakan yang efisien dan nilai jualnya yang tinggi (Millamena, 2002; Zainuddin et al., 2004b). Sebagai contoh kerapu macan (Epinephelus fuscoguttatus) merupakan salah satu jenis yang umum dibudidayakan di Indonesia (Zainuddin et al., 2008). Penelitian tentang aspek nutrisi ikan kerapu yang dilaporkan umumnya pada komponen utama pakan diantaranya kebutuhan protein (Shiau dan Lan, 1996), karbohidrat (Shiau dan Lin, 2001) dan kebutuhan lipid (Lin dan Shiau, 2003). Informasi tentang kebutuhan mineral bagi ikan kerapu sangat terbatas. Penelitian Zhou et al. (2004) melaporkan kebutuhan P dalam pakan kerapu E. coioides sebesar 0,86% dari total pakan, serta Ye et al. (2006) melaporkan 2 penambahan Ca dan P masing-masing 6 g/kg pakan memberikan pertumbuhan yang optimum pada spesies yang sama. Namun demikian, penelitian kebutuhan akan Ca dan P dalam pakan pada jenis kerapu macan E. fuscoguttatus masih sangat terbatas. Penelitian ini bertujuan untuk mengetahui kadar Ca dan P dalam pakan yang sesuai untuk pertumbuhan, efisiensi pakan juvenil kerapu macan, komposisi proksimat tubuh, morfometri dan kandungan mineral tubuh juvenil kerapu macan. Manfaat yang dapat diperoleh dari penelitian ini adalah diperolehnya informasi baru bagi pengembangan iptek khususnya dalam bidang nutrisi dengan penentuan komposisi pakan juvenil kerapu macan yang tepat, akurat dan efisien serta sebagai informasi baru bagi industri/perusahaan pakan khususnya pakan kerapu macan. II. METODE PENELITIAN 2.1. Tempat dan Waktu Penelitian berlangsung dari Juli – Nopember 2009 di Balai Budidaya Air Payau Takalar. 2.2. Pakan uji Pakan uji yang digunakan sebanyak 6 pakan dengan dua level P (dengan dan tanpa penambahan P). Pakan uji merupakan pakan dengan formulasi sendiri dengan formulasi pakan dasar seperti yang disajikan pada Tabel 1. Pakan A dan B tanpa penambahan P, pakan C, D, E dan F dengan penambahan P masing-masing 6 g/kg (sumber P digunakan NaH2PO42H2O). Kedalam masing-masing pakan tersebut (A, B, C, D, E, dan F) masingmasing ditambahkan Ca berturut-turut sebesar 0,0; 6,0; 0,0; 6,0; 12,0; dan 18,0 g/kg (Ca-carbonat sebagai sumber Ca) (Tabel 2). http://www.itk.fpik.ipb.ac.id/ej_itkt22 Zainuddin Tabel 1. Komposisi pakan dasar Komposisi bahan Tepung ikan Tepung kedelai Tepung jagung Minyak jagung Minyak ikan Mineral mix tanpa Ca & P Vitamin mix Sellulosa g/kg pakan kering 400 150 100 70 65 20 20 175 Tabel 2. Komposisi pakan uji setelah ditambahkan Ca dan P Bahan pakan Tepung ikan Tepung kedelei Tepung jagung Minyak ikan Minyak jagung Mineral mix Vitamin mix Ca P Sellulosa A 400 150 100 65 70 20 20 0 0 175 B 400 150 100 65 70 20 20 6 0 169 Semua bahan kering yang digunakan ditimbang dan dicampur dalam mixer selama 15 menit, minyak ikan dan minyak jagung ditambahkan ke dalam mixer lalu diaduk selama 15 menit. Selanjutnya ditambahkan air sebanyak 250 mL/kg pakan kering dan diaduk selama 15 menit. Selanjutnya dibuat pakan dalam bentuk semimoist. Pakan ini disimpan dalam lemari pendingin pada suhu –20oC sebelum digunakan. Setelah analisis proksimat diperoleh komposisi nutrisi pakan uji seperti yang disajikan pada Tabel 3. 2.3. Prosedur penelitian Juvenil kerapu macan E. fuscoguttatus diperoleh dari pendederan Balai Budidaya Air Payau Takalar. Jumlah juvenil yang disiapkan sebanyak 300 ekor dengan panjang rata-rata 12 cm dan Pakan uji (g/kg pakan) C D E 400 400 400 150 150 150 100 100 100 65 65 65 70 70 70 20 20 20 20 20 20 0 6 12 6 6 6 169 163 157 F 400 150 100 65 70 20 20 18 6 151 bobot rata-rata 40 g/ekor. Juvenil-juvenil ini diadaptasikan di dalam bak-bak berkapasitas 5 ton pada kondisi lingkungan suhu 31oC dan salinitas 30 ppt. Selama masa adaptasi ikan uji diberi pakan uji A (pakan tanpa penambahan Ca dan P) hingga juvenil terbiasa memakan pakan buatan secara total. Aklimatisasi dilakukan selama 3 hari supaya P yang terdeposit masih sangat rendah. Juvenil yang telah diaklimatisasi dan diseleksi menurut ukuran panjang dan bobot yang relatif sama ditebar secara acak ke dalam 18 buah ember plastik berkapasitas 80 L. Setiap wadah diisi juvenil sebanyak 5 ekor dengan panjang tubuh rata-rata 12 ± 0,3 cm dan bobot rata-rata 40 ± 0,85 g per ekor. Air yang digunakan adalah air laut yang telah disaring dengan salinitas 30 ± 0,5 ppt. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 3 Pengaruh Calsium dan Fosfor Terhadap Pertumbuhan, Efisiensi Pakan... Tabel 3. Komposisi nutrisi pakan uji Komposisi (%) Protein Lemak Serat BETN Kadar Ca P kasar kasar kasar Abu A 41,31 22,83 2,11 16,19 17,56 2,44 1,36 B 40,96 23,11 2,32 15,79 17,83 2,99 1,70 C 41,12 22,97 2,08 15,73 18,10 3,18 1,86 D 41,28 23,04 2,18 15,37 18,13 3,58 1,89 E 40,88 22,86 2,29 16,14 17,83 3,68 1,91 F 41,42 23,19 2,13 15,30 17,96 3,79 1,94 Ket. Hasil analisis Lab. Kimia Makanan Ternak, Fak. Peternakan Unhas,2009 Pakan Cahaya alami digunakan selama pemeliharaan.Pemberian pakan dilakukan secara satiasi (kenyang) yang diberikan dua kali perhari yakni pukul 09.00 dan 16.00 selama 8 minggu. 1. Pertumbuhan bobot, WG = 100x 2.4. Sampling dan metode analisa Pada awal percobaan, sebanyak 5 ekor juvenil diambil secara acak untuk analisa awal komposisi tubuh.Pada akhir penelitian (hari ke-60 pemeliharaan), 5 ekor juvenil diambil secara acak dari setiap tangki untuk analisa komposisi tubuh. Kandungan mineral pada pakan dan tubuh juvenil ikan diukur dengan menggunakan plasma atomic emission spectrophotometer. Kadar air, protein kasar, lemak kasar dari pakan uji, dan tubuh ikan diukur dengan metode standar (AOAC, 1984). Kadar air diukur melalui pengeringan dalam oven pada 105oC selama 24 jam; protein kasar dianalisa dengan metode Kjeldahl; lemak kasar dianalisa dengan metode ekstraksi ether melalui system Soxlec. Analisa kadar abu dilakukan dengan pengabuan pada suhu 550oC selama 24 jam dalam muffle furnace. 3. Kandungan mineral Ca dan P pada tubuh 4. Komposisi proksimat tubuh yang diuji pada seluruh tubuh 5. Morfometri meliputi: a. Faktor kondisi, CF = 100x 2.5. Parameter uji Paramater yang akan diuji dalam penelitian ini sama dengan yang dilakukan oleh Ye et al. (2006) sebagai berikut: 4 2. Efisiensi pakan, FE = 100% b. Indeks somatic organ dalam, VSI = 100x 2.6. Rancangan Perlakuan Adapun perlakuan yang diterapkan dalam penelitian ini adalah penambahan Ca dan P dalam pakan masing-masing sebagai berikut: Perlakuan A: penambahan Ca 0 g/kg pakan dan P 0 g/kg pakan, Perlakuan B: penambahan Ca 6 g/kg pakan dan P 0 g/kg pakan, Perlakuan C: penambahan Ca 0 g/kg pakan dan P 6 g/kg pakan, Perlakuan D: penambahan Ca 6 g/kg pakan dan P 6 g/kg pakan, Perlakuan E: penambahan Ca 12 g/kg pakan dan P 6 g/kg pakan, dan Perlakuan F: penambahan Ca 18 g/kg pakan dan P 6 g/kg pakan http://www.itk.fpik.ipb.ac.id/ej_itkt22 Zainuddin Setiap perlakuan diberi ulangan sebanyak 3 kali sehingga terdapat 18 satuan percobaan.Rancangan percobaan yang digunakan adalah rancangan acak lengkap (RAL). 2.7. Analisis Statistik Seluruh data dipresentasikan secara rata-rata dan dianalisis varians satu arah (SPSS for Windows ver 11,5) untuk menguji perbedaan antara perlakuan. Jika terdapat pengaruh perlakuan terhadap parameter uji, maka dilanjutkan dengan uji beda nilai tengah BNT pada taraf kepercayaan 5%. III. HASIL DAN PEMBAHASAN 3.1. Pertumbuhan Bobot Relatif dan Efisiensi Pakan Data hasil pertumbuhan bobot relatif dan efisiensi pakan juvenil ikan kerapu macan selama pemeliharaan disajikan pada Tabel 4. Berdasarkan Tabel 4 menunjukkan bahwa rata-rata pertumbuhan bobot relatifjuvenil kerapu macan tertinggi diperoleh pada perlakuan C sebesar 66,25% diikuti oleh perlakuan F 65,25%, perlakuan B 65,00%, perlakuan E 52,00%, perlakuan D 44,75% dan terendah pada perlakuan A sebesar 42,25%. Sementara itu, efisiensi pakan tertinggi diperoleh pada perlakuan C sebesar 12.30% diikuti perlakuan B 11.30%, perlakuan F 10.61%, perlakuan E 8.88%, perlakuan D 7.88% dan terendah pada perlakuan A sebesar 7.70%. Hasil penelitian ini menunjukkan bahwa pertumbuhan bobot dan efisiensi pakan juvenile yang tidak ditambahkan Ca dan P dalam pakannya (perlakuan A) secara signifikan lebih rendah dibandingkan dengan perlakuan lainnya yang di dalam pakannya ditambahkan Ca dan P. Hal ini disebabkan karena pakan A (tanpa Ca dan P) mengakibatkan juvenile ikan kerapu macan mengalami defisiensi terhadap Ca dan P. Defisiensi terhadap Ca dan P tidak hanya mengakibatkan pertumbuhan yang terhambat tetapi efisiensi pakan juga jelek serta dalam jangka panjang ikan akan mengalami malformation (Steffens, 1989). Hasil penelitian menunjukkan perlakuan C (penambahan P 6g/kg) menyebabkan pertambahan bobot dan efisiensi secara nyata signifikan (P<0,05) lebih tinggi dibandingkan tanpa penambahan P. Hal ini menunjukkan bahwa pada pakan uji dengan penambahan P sebesar 6g/kg telah mencukupi kebutuhan juvenile sehingga Tabel 4. Pertumbuhan bobot relatif dan efisiensi pakanjuvenil kerapu macan pada semua perlakuan Perlakuan Penambahan Rerata Pertumbuhan Efisiensi Pakan Ca dan P dalam pakan bobot relatif (%) (g/kg pakan) (%) A (0 Ca, 0 P) 42,25±5,303a 7.70±1,041a b B (6 Ca, 0 P) 65,00±4,419 11.30±0,352bc C (0 Ca, 6 P) 66,25±1,767bc 12.30±0,288b a D (6 Ca, 6 P) 44,75±0,353 7.88±0,354a E (12 Ca, 6 P) 52,00±9,899acd 8.88±1,346ac bd F (18 Ca, 6 P) 65,25±9,545 10.61±1,823bc Ket. Huruf superscript yang berbeda dibelakang angka pada kolom yang sama menunjukkan perlakuan berbeda nyata (P<0,05) Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 5 Pengaruh Calsium dan Fosfor Terhadap Pertumbuhan, Efisiensi Pakan... dapat bertumbuh dengan baik. Demikian pula halnya dengan kondisi lingkungan pemeliharaan yang sesuai dengan pertumbuhan dan efisiensi pakan juvenile kerapu macan. Selama masa pemeliharaan kondisi lingkungan meliputi oksigen terlarut sebesar 4,5 ± 0,15 ppm, suhu 31 ± 0,5 oC, salinitas 30 ± 0,5 ppt dan pH 8 ± 0,5. Kondisi lingkungan seperti ini sangat mendukung kelangsungan hidup organisme budidaya. Sebagai perbandingan dengan peneliti lain yang menunjukkan terjadinya pertumbuhan yang lambat dan efisiensi pakan yang rendah akibat defisiensi P pada ikan sea bass Eropa (Oliva-Teles and Pimentel-Rodrigues, 2004), dan haddock (Roy and Lall, 2003). 3.2. Komposisi Proksimat Tubuh dan Morfometri Komposisi proksimat tubuh juvenil kerapu macan yang diberi pakan dengan kandungan Ca dan P yang berbeda disajikan pada Tabel 5. Hasil penelitian menunjukkan bahwa pakan uji tanpa penambahan Ca dan P memiliki kandungan lemak tertinggi dibandingkan dengan perlakuan lainnya. Kandungan lemak tubuh semakin menurun seiring dengan peningkatan level penambahan Ca dan P ke dalam pakan meskipun tidak berbeda secara signifikan. Menurunnya kandungan lemak ini sebagai respon terhadap peningkatan jumlah P yang ada di dalam pakan dan hal ini terlihat pada setiap perlakuan penambahan Ca dan P di dalam pakan. Pakan uji tanpa penambahan P menyebabkan kadar abu pada tubuh lebih rendah, demikian pula halnya dengan kandungan BETN (Bahan Ekstraks Tanpa Nitrogen) meskipun keduanya tidak berbeda secara signifikan. Hal ini disebabkan karena fungsi P dalam pembentukan tulang sangat signifikan. Dalam proses mineralisasi tulang Ca dan P memiliki peran yang penting karena 6 sekitar 80 – 90% unsur tulang tersusun dari Ca, P dan Mg. Pakan uji tanpa penambahan P menyebabkan faktor kondisi, indeks somatic organ dalam dan kandungan lemak tubuh tinggi. Hasil yang sama diperoleh pada penelitian-penelitian sebelumnya (Skonberg et al., 1997; Zainuddin dkk. 2000; Vielma et al., 2002; Roy dan Lall, 2003). Akumulasi lemak dalam tubuh sebagai dampak dari defisiensi P berhubungan dengan perubahan metabolisme intermedier yang lebih cepat dari pakan yang dikonsumsi (Vielma et al., 2002). Dengan demikian deposit lemak akan berlangsung lebih cepat jika pakan kekurangan atau defisiensi P. Berdasarkan penelitian ini, konsumsi pakan pada perlakuan tanpa penambahan P juvenil mengkonsumsi pakan lebih sedikit. Hal ini membuktikan bahwa akumulasi lipid tidak berhubungan dengan banyaknya pakan yang dimakan. 3.3. Kadar Abu dan Kandungan Ca dan P pada Tulang Kadar abu dan kandungan mineral pada tulang belakang juvenil kerapu macan disajikan pada Tabel 6. Hasil penelitian menunjukkan kadar abu, kandungan Ca dan P pada tulang ikan kerapu macan secara signifikan (P<0,05) lebih rendah pada pakan uji tanpa penambahan P. Kecuali perlakuan A, semua perlakuan lainnya mengalami peningkatan kadar abu tulang juvenil ikan kerapu macan. Hal ini menunjukkan bahwa penambahan Ca dan P berpengaruh positif terhadap peningkatan kadar abu tulang. Demikian pula halnya dengan kandungan Ca dan P pada tulang menunjukkan peningkatan yang signifikan pada semua perlakuan penambahan Ca dan P dibandingkan dengan perlakuan tanpa penambahan Ca dan P (Tabel 7). Hal ini mempertegas http://www.itk.fpik.ipb.ac.id/ej_itkt22 Zainuddin Tabel 5 Komposisi tubuh dan morfometri juvenil kerapu macan yang diberi pakan dengan kandungan Ca dan P yang berbeda Komposisi (%) Tubuh Protein Lemak Sera kasar BETN Kadar Abu Morfometri CF (g/cm3) VSI Awal Perlakuan Penambahan Ca dan P dalam pakan (g/kg pakan) A B C D E F 76.02 8.04 5.51 1.98 8.45 79.75 7.44 1.95 2.16 8.71 79.27 6.65 1.27 2.60 10.21 78.21 6.90 2.75 2.69 9.46 78.06 6.56 2.30 2.88 10.21 79.63 6.42 1.79 2.22 9.95 77.73 6.42 3.32 3.32 10.06 2,30 3.18 2.34 4.09 2.29 3.21 2.27 4.23 2.55 4.22 2,42 3.93 2,57 4.65 Ket. Perlakuan tidak berpengaruh nyata (P>0,05); CF = faktor kondisi; VSI = indeks somatic organ dalam; BETN = Bahan Ekstrak Tanpa Nitrogen Tabel 6. Kadar abu dan kandungan mineral pada tulang juvenil kerapu macan pada semua perlakuan Perlakuan Penambahan Ca dan P dalam pakan (g/kg pakan) Konsentrasi Awal (%) A B C D E F a b b b b Kadar Abu 31.18 26.44 32.94 31.50 31.11 32.27 31.55b a b b b b Ca 10.39 8.96 10.96 10.92 10.67 11.17 11.04b P 5.02 5.10a 6.44b 6.16b 6.23b 6.57b 6.43b Ket. Huruf superscript yang berbeda dibelakang angka pada baris yang sama menunjukkan perlakuan berbeda nyata (P<0,05) bahwa P sangat dibutuhkan dalam proses mineralisasi tulang, karena 80 – 90% tulang tersusun oleh P, selain Ca dan Mg. Hasil yang sama pada penelitian Roy dan Lall (2003) yang melaporkan penggunaan pakan tanpa penambahan P menyebabkan kadar abu, Ca dan P secara nyata lebih rendah pada tulang dan operculum ikan haddock. Penambahan Ca dalam pakan berpengaruh signifikan terhadap kadar abu, Ca dan P tulang pada dua level P yang berbeda. Ketika pakan tidak ditambahkan dengan P, penambahan Ca pada pakan basal memberikan pengaruh yang sama terhadap kadar abu, Ca dan P tulang. Hal ini menunjukkan bahwa jika P tidak tersedia maka penambahan Ca juga tidak akan mampu memperbaiki proses mineralisasi tulang atau deposit Ca dan P. Ca dan P merupakan mineral yang saling sinergis (Zainuddin dkk. 2000). Penelitian yang menunjukkan hasil yang sama telah dilakukan diantaranya pada ikan black sea bream (Hossain and Furuichi, 1999) dan atlantic salmon (Vielma and Lall, 1998a). Lebih lanjut hasil penelitian ini menunjukkan bahwa penambahan Ca di atas 12 g/kg pakan akan menurunkan kadar abu, Ca dan P tulang. Hasil penelitian serupa pada kerapu batik (Ye et al., 2006). Porn-Ngam et al. (1993) melaporkan bahwa absorbsi P dapat terhalang akibat adanya peningkatan Ca dalam pakan. Hal ini terjadi karena pada ikan laut mampu menyerap lebih banyak Ca dari lingkungannya. Akan tetapi, penelitian Vielma and Lall (1998a) menunjukkan bahwa penambahan Ca dalam pakan tidak memberikan dampak negatif terhadap kadar abu, Ca dan P Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 7 Pengaruh Calsium dan Fosfor Terhadap Pertumbuhan, Efisiensi Pakan... tulang ketika P juga ditambahkan. Berbeda halnya di perairan tawar yang membutuhkan Ca lebih tinggi di dalam pakannya. Hal ini bisa saja terjadi karena kondisi media air tawar dan air laut yang digunakan berbeda, dimana diketahui bahwa kandungan Ca pada air laut jauh lebih banyak dibandingkan pada air tawar. IV. KESIMPULAN Berdasarkan Pertumbuhan bobot relatif,efisiensi pakan, komposisi proksimat tubuh dan kandungan mineral tubuh juvenil ikan kerapu macan secara signifikan mengalami perubahan setelah ditambahkan Ca dan P di dalam pakannya. Perlakuan terbaik dari penelitian ini adalah penambahan P sebesar 6g/kg pakan dan Ca 0g/kg ke dalam pakan. DAFTAR PUSTAKA Hossain, M.A., and M. Furuichi. 1999. Necessity of dietary calcium supplement in black sea bream. Fish. Sci., 65:893-897. Hossain, M.A., and M. Furuichi. 2000a. Essentiality of dietary calcium supplement in redlip mullet Liza haematocheila. Aquac. Nutr., 6:33– 38. Hossain, M.A., and M. Furuichi. 2000b. Necessity of calcium supplement to the diet of Japanese flounder. Fish. Sci., 66:660–664. Hossain, M.A., and M. Furuichi. 2000c. Essentiality of dietary calcium supplement in fingerling scorpion fish (Sebastiscus marmoratus). Aquaculture, 189:155–163. Lall, S.P. 2002. The minerals. In: Halver, J.E., Hardy, R.W. (Eds.), Fish Nutrition, 3rd ed. Academic Press, San Diego, CA, 259–308pp. 8 Lin, Y.H. and S.Y. Shiau. 2003. Dietary lipid requirement of grouper, Epinephelus malabaricus, and effects on immune responses. Aquaculture, 225:243–250. Millamena, O.M. 2002. Replacement of fish meal by animal byproduct meals in a practical diet for growout culture of grouper Epinephelus coioides. Aqua-culture, 204:75–84. Oliva-Teles, A. and A. PimentelRodrigues. 2004. Phosphorous requirement of European sea bass (Dicentrarchus labrax L.) juveniles. Aquac. Res., 35:636-642. Porn-Ngam, N., S. Satoh, T. Takeuchi, and T. Watanabe. 1993. Effect of the ratio of phosphorous to calcium on zinc availability to rainbow trout in high phosphorous diet. Nippon Suisan Gakkaishi, 59:2065-2070 Roy, P.K. and S.P. Lall. 2003. Dietary phosphorous requirement of juvenile haddock (Melanogrammus aeglefinus L.). Aquaculture, 221:451-468. Shiau, S.Y. and C.W. Lan. 1996. Optimum dietary protein level and proteinto energy ratio for growth of grouper (Epinephelus malabaricus). Aquaculture, 145: 259–266. Shiau, S.Y. and Y.H. Lin. 2001. Carbohydrate utilization and its proteinsparing effect in diets for grouper, Epinephelus malabaricus. Anim. Sci., 73:299–304. Skonberg, D.I., L. Yogev, R.W. Hardy, and F.M. Dong. 1997. Metabolic response to dietary phosphorous intake in rainbow trout (Oncorhynchus mykiss). Aquaculture, 157:11-24. Steffens, W. 1989. Principles of fish nutrition. John Wiley & Sons. New York. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Zainuddin Suderajad, P. 2003. Pengaruh penambahan kalsium dalam pakan terhadap pertumbuhan dan sintasan juvenil ikan bandeng (Chanos chanos). Skripsi. Fakultas Ilmu Kelautan dan Perikanan. Universitas Hasanuddin. Makassar. Tacon, A.G. 1992. Nutritional fish pathology. Morphological signs of nutrient deficiency and toxicity in farmed fish. FAO Fisheries Technical Paper, vol. 330. FAO, Rome, Italy. 75 pp. Vielma, J. and S.P. Lall. 1998a. Phosphorus utilization by Atlantic salmon (Salmo salar) reared in freshwater is not influenced by higher dietary calcium intake. Aquaculture, 160:117–128. Vielma, J. and S.P. Lall. 1998b. Control of phosphorous homeostasis of Antalantic salmon (Salmo salar) in fresh water. Fish Physiol. Biochem., 19:83-93. Vielma, J., J. Koskela, and K. Ruohonen. 2002. Growth, bone mineralizeation, and heat and low oxygen toelance in European whitefish (Coregonus lavaretus L.) fed with graded levels of phosphorous. Aquaculture, 212:321-333. Ye, C.X., Y. Liu, L. Tian, K. Mai, Z. Y. Du, H. Yang, and J. Niu. 2006. Effect of dietary calcium and phosphorus on growth, feed efficiency, mineral content and body composition of juvenil grouper, Epinephelus coioides. Aquaculture, 255:263–271 Zainuddin, I. Mokoginta, R. Affandi, dan D. Yusadi. 2000. Kadar Fosfor Optimum dalam Pakan Benih Ikan Jambal Siam (Pangasius sutchi Fowler), Hayati, 7(2):41-44. Zainuddin. 2001. Pengaruh Pemberian Mineral Fosfor dalam Pakan terhadap Pertumbuhan dan Kelangsungan Hidup Juvenil Udang Windu (Penaeus monodon), Lembaga Penelitian, Universitas Hasanuddin. Makassar. Zainuddin. 2004a. Pengaruh CalsiumFosfor Dengan Rasio Berbeda Terhadap Pertumbuhan dan Efisiensi Pakan Udang Windu (Penaeus Monodon Fabr.). Lembaga Penelitian, Universitas Hasanuddin, Makassar. Zainuddin, A. Niartiningsih, Arifin, dan Supriadi. 2004. Pembesaran ikan kerapu macan (Epinephelus fuscoguttatus) dalam Karamba Jaring Apung. Adaptive Research and Extention. Proyek Pengelolaan Sumberdaya Pesisir dan Laut (MCRMP) Kabupaten Pangkajene dan Kepulauan. Zainuddin, M.N. Nessa, M.I. Djawad, dan D. Dh. Trijuno. 2008. Deposit glikogen juvenil ikan kerapu macan (Epinephelus fuscogu-ttatus) pada frekeunsi pemberian pakan yang berbeda. Torani,18(2):179–186. Zhou, Q.C., Y.J. Liu, K.S. Mai, and L.X. Tian. 2004. Effect of dietary phosphorus level on growth, body composition, muscle and bonemineral concentrations for orangespotted grouper Epinephelus coioides reared in floating cages. J. World Aquac. Soc., 35:427–435. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 9 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 10-16, Desember 2010 IDENTIFIKASI JENIS IKAN ANEMON (Amphiprioninae) DAN ANEMON SIMBIONNYA DI KEPULAUAN SPERMONDE, SULAWESI SELATAN IDENTIFICATION OF ANEMONEFISHES (Amphiprioninae) AND THEIR SIMBIONT IN SPERMONDE ARCHIPELAGO, SOUTH SULAWESI Inayah Yasir, Syafiuddin, dan Sumarjito Jurusan Ilmu Kelautan, Fakultas Ilmu Kelautan dan Perikanan, Universitas Hasauddin Jln. Perintis Kemerdekaan Km. 10 Makassar-90245 email: [email protected] ABSTRACT The study was conducted in June 2009 around the waters of Samalona, Barranglompo, Koedingareng Keke, Badi, Langkai and Kapoposang islands, representing four zones of Spermonde Archipelago of South Sulawesi. Seven species of anemonefish from two genera were found living symbiotically with 7 species of sea anemones. Those fishes were Amphiprion clarkii, A. melanopus, A. ocellaris, A. sandaracinos, A. perideraion, A. polymnus and Premnas biaculeatus. Three of these fishes were simbiotically found with one species of anemone (specific symbiont), and one species of fish was symbiotically found with five species of anemones. Keywords: Amphiprioninae, Spermonde Archipelago, anemone symbiont ABSTRAK Penelitian dilakukan pada bulan Juni 2009 di perairan pulau Samalona, P. Barranglompo, P. Koedingareng Keke, P. Badi, P. Langkai dan P. Kapoposang yang mewakili empat zona perairan Kepulauan Spermonde Sulawesi Selatan. Pada penelitian ini ditemukan 7 jenis ikan giru yang berasal dari 2 genera dan hidup bersimbiosis dengan 7 jenis anemon laut. Ketujuh jenis ikan giru tersebut adalah Amphiprion clarkii, A. melanopus, A. ocellaris, A. sandaracinos, A. perideraion, A. polymnus dan Premnas biaculeatus. Tiga diantara ikan giru ini ditemukan bersimbiosis hanya dengan 1 jenis anemon saja (simbion spesifik), sedangkan 1 jenis lainnya ditemukan bersimbiosis dengan 5 jenis anemon. Kata Kunci: Amphiprioninae, Kepulauan Spermode, anemon I. PENDAHULUAN Ikan merupakan organisme yang jumlah biomassanya terbesar dan juga organisme besar yang mencolok yang dapat ditemui di ekosistem terumbu karang. Banyaknya celah dan lubang yang terdapat di daerah terumbu karang memberikan tempat tinggal, perlindungan, tempat mencari makan dan berkembang biak bagi ikan dan hewan invertebrata yang berada disekitarnya (Nybakken dan Bertness, 2004). Lebih dari 4000 species ikan (atau sekitar 18% 10 dari jumlah species ikan yang ada di seluruh dunia) dapat ditemukan di daerah terumbu karang. Umumnya ikan-ikan yang hidup di daerah terumbu karang ini berukuran kecil dan menetap sepanjang hidupnya di daerah tersebut. Salah satu jenis ikan karang yang hidup di daerah terumbu karang adalah ikan-ikan dari Familia Pomacentridae, subfamilia Amphiprioninae. Semua ikan dalam subfamilia Amphiprioninae hidup bersimbiosis dengan anemon laut (Dunn, 1981; Fautin, 1991) dalam hubungan simbiosis ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Yasir et al. mutualisme (Fautin dan Allen, 1992) sehingga kelompok ikan ini juga dikenal sebagai ikan anemon (anemonefish). Pola warnanya yang indah, kemampuannya untuk hidup dalam akuarium dan hubungan simbiosis yang menarik dengan anemon membuat ikan ini sangat diminati oleh pencinta ikan hias laut (Fautin dan Allen, 1997). Akibat kepopularannya, ikan anemon dijuluki juga ‘ikan mas’ akuarium air laut (Hoff, 1996). Kepulauan Spermonde,yang oleh masyarakat di kepulauan ini dikenal dengan nama Pulau-pulau Sangkarang, adalah gugusan pulau-pulau yang terletak di pesisir barat daya Pulau Sulawesi. Mata pencaharian utama penduduk di Kepulauan Spermonde adalah sebagai nelayan yang memanfaatkan sumberdaya wilayah pesisir. Menurunnya hasil tangkapan ikan di daerah sekitar Kepulauan Spermonde memaksa hanya nelayan dengan modal menengah hingga besar yang dapat tetap melaut, sedangkan yang hanya bermodalkan perahu kecil tanpa motor harus mencari sekitar pulau saja. Alternatif lain muncul seiring dengan meningkatnya jumlah pemelihara ikan hias air laut di seluruh dunia (Ziemann, 2001). Permintaan yang meningkat akhirnya berdampak juga terhadap ikanikan terumbu karang yang tadinya tidak dieksploitasi karena tidak dikonsumsi. Ikan-ikan anemon termasuk yang paling terkena imbas dari trend ini, karena permintaan dunia yang memang tinggi untuk kelompok ikan ini (Wood, 2001; Wabnitz, 2003). Selain itu, ketergantungannya terhadap anemon membuat ikan ini mudah untuk ditangkapi karena di alam ikan ini tidak pernah jauh meninggalkan anemon simbionnya. Meskipun telah dikenal dan dieksploitasi untuk keperluan ekspor, belum ada data akurat tentang jenis ikan anemon yang dapat ditemukan di pulau-pulau Kepulauan Spermonde. Diketahui di dunia terdapat 28 jenis ikan anemone dari 2 genera yaitu genus Amphiprion dengan 27 species dan genus Premnas dengan 1 species (Allen, 1991) yang tersebar di seluruh dunia. Menurut Allen (1991) di Indonesia ditemukan 9 species yaitu Amphiprion akallopsis, A. clarkii, A. ephippium, A. frenatus, A. melanopus, A. ocellaris, A. periderion, A. polymnus, A. sebae, dan A. sandaracinos, lalau oleh Kuiter dan Tonozuka (2001) ditambahkan dengan A. percula. Semua ikan anemon hidup bersimbiosis mutualistik dengan anemon tertentu (Allen, 1991). Dalam simbiosis ini, ikan mendapat proteksi dan memakan material non-metabolik yang dikeluarkan oleh anemon. Di sisi lain, anemon ‘dibersihkan’ dan dilindungi dari predator oleh ikan simbionnya (Randall dan Fautin, 2002). Upaya identifikasi suatu organisme diperlukan dalam pengenalan jenis berdasarkan sifat-sifat morfologi, anatomi, bahkan perilaku organisme tersebut. Identifikasi didasarkan pada karak-ter fisik dari bagian-bagian tubuh. Untuk ikan kelompok Amphiprioninae, karakter yang umum digunakan adalah perpaduan antara penggunaan karakter morfometrik, meristik dan pola pewarnaan tubuh (Allen, 1991). II. METODE PENELITIAN Penelitian dilaksanakan dari bulan Juni sampai bulan Agustus 2009 pada 4 zona Kepulauan Spermonde yang berada dalam kawasan pemerintahan Provinsi Sulawesi Selatan. Pengambilan sampel dilakukan pada tanggal 6 hingga 10 Juni 2009. Untuk zona 1 diwakili oleh Pulau Samalona, zona 2 diwakili oleh P. Barrang lompo dan P. Koedingareng Keke sedangkan zona 3 diwakili oleh P. Badi. Pulau Kapoposang dan P. Langkai adalah pulau-pulau yang mewakili zona 4 (Gambar 1). Sampling dilakukan dengan Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 11 Identifikasi Jenis Ikan Anemon (Amphiprioninae) dan Anemon Simbionnya... cara menyelam menggunakan peralatan selam hingga kedalaman 25 meter. Satu pulau pengamatan dibagi ke dalam 4 daerah untuk memudahkan sampling. Sampling dimulai dengan langsung turun ke kedalaman 25 meter sebagai posisi awal (starting point), lalu bergerak ke satu arah hingga batas daerah sampling. Sampling dilanjutkan dengan berbalik ke arah diagonal mengikuti kontur perairan naik ke kedalaman 15 meter. Lanjut lagi dengan cara yang sama ke kedalaman 5 meter dst. Ikan anemon yang ditemukan diamati pola warna tubuh dan kecenderungan bentuk tubuh. Pola warna ikan yang ditemukan kemudian dibandingkan dengan pola warna ikan pada gambar yang dibawa serta. Setelah proses identifikasi visual selesai, ikan kemudian diambil gambarnya. Bila identifikasi dengan pola warna masih meragukan, 2 ekor dari kelompok tersebut kemudian ditangkap dengan menggunakan serok lalu dipasangi label dan dimasukkan ke dalam tas jaring untuk kemudian diadakan identifikasi lanjutan di laboratorium Biologi Laut, Jurusan Ilmu Kelautan, Fakultas Ilmu Kelautan dan Perikanan. Gambar 1. Peta lokasi penelitian di Kepulauan Spermonde, Sulawesi Selatan 12 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Yasir et al. Untuk identifikasi jenis, anemon yang berasosiasi dengan ikan giru dicatat lalu diambil gambarnya sebagai bahan re-check nantinya untuk keperluan identifikasi di laboratorium. Pengukuran diameter anemon kemudian dilakukan lalu dilanjutkan dengan menghitung jumlah ikan yang ada dalam kelompok itu. Data penunjang lainnya termasuk organisme lain yang ikut menghuni anemon (bila ada), kondisi ikan Amphiprion (bertelur atau tidak) dan kondisi ekologi sekitar anemon juga dicatat dan diamati. Ikan yang identifikasi lapangannya dianggap meragukan, ditangkap untuk kemudian diidentifikasi lebih lanjut di laboratorium biologi laut dengan meng- gunakan kombinasi metode morfometrik dan meristik (Allen, 1991). III. HASIL DAN PEMBAHASAN Di lokasi penelitian ditemukan tujuh jenis dari dua genera dari subfamilia Amphiprioninae, yaitu Amphiprion clarkii, A.melanopus, A. ocellaris, A. perideraion, A. polymnus, A. sandaracinos dan Premnas biaculeatus. Ketujuh jenis Amphioprioninae yang ditemukan di lokasi penelitian dapat ditemukan di semua pulau lokasi penelitian yang masuk pada zona 1, 2 dan 3, namun pada zona 4, hanya ada 5 jenis yang ditemukan (Tabel 1). Tabel 1. Penyebaran Amphiprioninae berdasarkan zona di Kepulauan Spermonde Zona Pulau 1 Samalona 2 Kodingareng Keke dan Barrang Lompo 3 Pulau Badi 4 Kapoposang & Langkai Jenis Amphiprioninae Amphiprion clarkii Amphiprion melanopus Amphiprion ocellaris Amphiprion perideraion Amphiprion polymnus Amphiprion sandaracinos Premnas biaculeatus Amphiprion clarkii Amphiprion melanopus Amphiprion ocellaris Amphiprion perideraion Amphiprion polymnus Amphiprion sandaracinos Premnas biaculeatus Amphiprion clarkii Amphiprion melanopus Amphiprion ocellaris Amphiprion perideraion Amphiprion polymnus Amphiprion sandaracinos Premnas biaculeatus Amphiprion clarkii Amphiprion ocellaris Amphiprion perideraion Amphiprion polymnus Premnas biaculeatus Kedalaman (m) 1-10 1-10 1- 10 1- 10 16 1- 2 1- 10 1- 19 6 1- 13 1- 12 23 2 1- 15 1-19 4-6 4- 11 1- 15 9 3- 9 2- 12 3-11 5 -15 4-29 4-14 5-11 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 13 Identifikasi Jenis Ikan Anemon (Amphiprioninae) dan Anemon Simbionnya... Pada zona 1, zona 2 dan zona 3 ditemukan Amphiprion clarkii, A. melanopus, A. ocellaris, A. Polymnus, A. Sandaracinos, A. perideraion dan Premnas biaculeatus. Pada Zona 4 yang merupakan zona terluar, hanya 5 jenis Amphiprioninae yang ditemukan tanpa A. sandaracinos dan A. melanopus. Pola penyebaran yang tidak merata pada kelompok ikan kemungkinan besar disebabkan oleh kondisi lingkungan pulau, seperti gelombang, arus, kedalaman perairan dan kompleksitas terumbu karang pada masing-masing zona. Pada zona 1, 2 dan 3, kontur kedalaman terumbu sekeliling pulau berupa reef flat yang dilanjutkan dengan reef slope. Kompleksitas terumbu karang yang relatif bagus dan beragam jenisnya di hampir semua sisi pulau memungkinkan beragam anemon hidup yang pada akhirnya menyediakan tempat berlindung bagi ikan Amphiprioninae (Allen, 1998). Zona-zona ini relatif berbeda dibandingkan dengan zona 4 dengan areal yang cenderung sempit, dan keberagaman kondisi terumbunya yang kurang. Beberapa bagian pulau yang berupa drop off dengan kondisi arus yang cukup kuat kemungkinan besar menjadi kendala bagi anemon simbion ikan Amphiprioninae untuk hidup. Keberadaan jenis anemon juga turut mempengaruhi sebaran ikan Amphiprioninae (Allen, 1972 dan Dunn, 1981). Jenis dan jumlah anemon yang ditemukan di setiap pulau pada zona 1, 2 dan 3 cukup banyak bila dibandingkan dengan jenis dan jumlah anemon yang ditemukan di pulau-pulau yang mewakili zona 4. Adanya beberapa jenis ikan Amphiprioninae yang anemon simbionnya spesifik seperti A. sandaracinos yang hanya berasosiasi dengan Stichodactyla mertensii dan Premnas biaculeatus dengan Entacmaea quadricolor (Allen, 1991), membuat penyebarannya tidak 14 merata.A. clarkiiditemukan di setiap zona penelitian karena mampu menerima berbagai jenis anemon sebagai simbionnya. Dengan kata lain, jenis ini tidak memiliki 'host' yang spesifik (Allen, 1991; Dunn, 1981). Jenis anemon yang berbeda mempunyai jenis toksin yang berbeda pula, sehingga beberapa anemon memiliki toksin yang lebih kuat daya racunnya dibandingkan dengan jenis yang lain (Mebs, 1994). Di lokasi penelitian ditemukan A. clarkii yang mampu bersimbiosis dengan 5 (lima) jenis anemon yaitu Stichodactyla mertensii, S. gigantea, Heteractis crispa, H. aurora dan Entacmaea quadricolor. Dari hasil analisis kimia lendir yang menyelimuti A. clarkii disimpulkan kalau jenis ini menghasilkan lendir sendiri yang spesifik yang menyebabkan nematocyst anemon tidak ditembakkan (Mebs, 2009). Kemampuan ini menyebabkan A. clarkii dapat hidup di banyak jenis anemon. A. clarkii dapat hidup di tujuh jenis anemon (Tabel 1, Fautin dan Allen, 1997), namun di lokasi penelitian, hanya ditemukan berasosiasi dengan lima jenis anemon saja (Tabel 2). Berbeda dengan A. clarkii, A. sandaracinos ditemukan hanya berasosiasi dengan anemon jenis Stichodactyla mertensii. Anemon jenis ini ditemukan di semua pulau di zona 1, 2 dan 3, sedangkan di zona 4 hanya ditemukan di P. Langkai dengan ko-simbion A. clarkii. Premnas biaculeatus dan A. melanopus di lokasi penelitian hanya ditemukan berasosiasi dengan anemon Entacmaea quadricolor. Padahal menurut Allen (1991), A. melanopus juga ditemukan bersimbiosis dengan H. crispa dan H. magnifica. Kedua jenis anemon ini juga dijumpai di lokasi penelitian. Tidak dihuninya kedua jenis anemon ini oleh A. melanopus di lokasi penelitian membutuhkan penelitian lebih lanjut. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Yasir et al. H. crispa dan E. quadricolor masing-masing dapat bersimbiosis dengan 3 jenis Amphiprioninae (Tabel 2), namun H. crispa adalah jenis yang paling banyak ditemukan dan juga paling banyak dihuni oleh ikan Amphiprioninae. Hal ini disebabkan karena H. crispa dapat hidup di semua habitat yang bisa di tempati anemon untuk hidup. Berdasarkan pengamatan di lapangan, jenis ini umumnya menghuni habitat dengan kondisi pecahan karang hancur, pasir dan celah-celah karang. Kondisi ekologi ini pula yang mendominasi lokasi penelitian. Selain ikan dari subfamilia Amphiprioninae, di lokasi penelitian ditemukan beberapa jenis biota lain yang juga hidup berasosiasi dengan anemon. Yang paling umum ditemukan adalah ikan damsel Dascyllus trimaculatus, kepiting Neopetrolisthes maculatus, dan udang Periclemenes tosaensis. Namun jenis ini ditemukan tidak di semua jenis anemon. Untuk Dascyllus trimaculatus hanya ditemukan di anemon jenis H. magnifica, S. mertensii, H. crispa, S. haddoni dan S. gigantea. Neopetrolisthes maculatus hanya di anemon jenis S. mertensii, E. quadricolor, S. haddoni dan S. gigantea. Udang Periclimenes tosaensis hanya ditemukan bersimbiosis dengan anemon H. crispa, H. magnifica dan S. haddoni (Tabel 2). Berdasarkan hasil uji statistik dengan analisis regresi menunjukan bahwa tidak ada pengaruh yang nyata antara besarnya koloni dengan ukuran anemon simbionnya (P>0,05). Besar atau kecilnya koloni ikan simbionnya sepertinya lebih tergantung pada perilaku dari masing-masing jenis daripada ukuran anemon simbionnya. Anemon dari jenis S. haddoni dengan diameter 51 cm bersimbiosis dengan A. polymnus dengan jumlah hingga 31 ekor. Anemon jenis E. quadricolor yang umumnya berukuran besar, hanya dihuni oleh sepasang P. biaculeatus. Hal ini kemungkinan besar disebabkan karena pola perilaku jenis yang beragam. Sebagai contoh P. biaculeatus cenderung sangat agresif terhadap apapun yang mendekati anemon simbionnya termasuk peneliti sendiri. Jenis ini juga terkait sangat dekat dengan anemonnya. Tabel 2. Amphiprioninae dan jenis anemon simbionnya 7 No Simbiosis 1 2 3 4 5 6 1 Amphiprion clarkii X X X X X X X 2 Amphiprion melanopus 3 Amphiprion ocellaris X X 4 Amphiprion perideraion X X 5 Amphiprion polymnus X X 6 Amphiprion sandaracinos X X 7 Premnas biaculeatus 8 Dascyllus trimaculatus X X X X X 9 Neopetrolisthes maculatus X X X X 10 Periclimenes tosaensis X X X Keterangan: 1. Heteractis magnifica, 2. H. crispa, 3. H. aurora, 4. Stichodactyla haddoni, 5. S. gigantea, 6. S. mertensii, 7. Entacmaea quadricolor Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 15 Identifikasi Jenis Ikan Anemon (Amphiprioninae) dan Anemon Simbionnya... Hal berbeda diamati pada A. polymnus. Bila merasa terancam, koloni A. polymnus yang mendiami anemon cenderung meninggalkan anemone simbionya meskipun tidak terlalu jauh. Tetapi dua ikan dari koloni tersebut yang berukuran paling besar akan berlindung dengan cara masuk ke dalam mulut anemon untuk kemudian keluar lagi bila sumber gangguan telah hilang. IV. KESIMPULAN Terdapat tujuh jenis dari dua genera Amphiprioninae yang ditemukan di Kepulauan Spermonde berdasarkan pembagian zona yaitu Amphiprion clarkii, A. melanopus, A. perideraion, A. polymnus, A. ocellaris, A. sandaracinos dan Premnas biaculeatus. Ketujuh jenis Amphioprioninae yang ditemukan di lokasi penelitian berada pada zona 1, 2 dan 3, sedangkan di zona 4, hanya ditemukan lima jenis. Terdapat tujuh jenis anemon yang hidup bersimbiosis dengan genera Amphiprioninae di Kepulauan Spermode yaitu Heteractis aurora, H.crispa, H. magnifica, Stichodactyla mertensii, S. haddoni, S. gigantea dan Entacmaea quadricolor. DAFTAR PUSTAKA Allen, G. R. 1991. Damselfishes of the world. Germany, Hans A. Baensch. Dunn, D. F. 1981. The clownfish sea anemones: Stichodactylidae (Coelenterata: Actiniaria) and other sea anemones symbiotic with pomacentrid fishes. Transactions of the American Philosophical Society, 71:115. Fautin, D. G. 1991. The anemonefish symbiosis: what is known and what is not. Symbiosis, 10:23-46. 16 Fautin, D.G. and G.R. Allen. 1992. Field guide to anemonefishes and their host sea anemones. Australia, Western Australian Museum. Fautin, D.G. and G.R. Allen. 1997. Anemone fishes and their host sea anemones: a guide for aquarists and divers. Western Australian Museum. Hoff, F.H. 1996. Conditioning, spawning and rearing of fish with emphasis on marine clownfish. Florida, Dade City, Aquaculture Consultants, Inc. Kuiter R.H. and T. Tonozuka. 2001. Pictorial guide to Indonesian reef fishes – Zoonetics- Australia Mebs, D. 1994. Anemonefish symbiosis: vulnerability and resistance of fish to the toxin of the sea anemone. Toxicon, 32:1059-1068. Mebs, D. 2009. Chemical biology of the mutualistic relationships of sea anemones with fish and crustaceans, Toxicon, doi:10.1016/ j.toxicon. 2009.02.027 Nybakken, J. W. and M. D. Bertness. 2004. Marine Biology: An Ecological Approach. Randall, J. E dan D.G. Fautin. 2002. Fishes other than anemonefishes that associate with sea anemones. Coral Reefs, 21:188–190 Wabnitz, C.; M. Taylor; E. Green and T. Razak. 2003. From Ocean to Aquarium. Cambridge, UK, UNEPWCMC: 64. Wood, E.M. 2001. Collection of coral reef fish for aquaria: global trade, conservation issues and management strategies. Marine Conservation Society, UK. 80pp. Ziemann, D. A. 2001. The potential for the restoration of marine ornamental fish populations through hatchery releases. Aquarium Sciences and Conservation, 3:107–117. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 17-25, Desember 2010 KERAGAAN REPRODUKSI IKAN KERAPU BEBEK (Cromileptes altivelis) DARI ALAM (F-0), INDUK GENERASI PERTAMA (F-1), DAN INDUK GENERASI KE DUA (F-2) REPRODUCTION PERFORMANCE OF HUMPBACK GROUPER (Cromileptes altivelis) FROM WILD BROODSTOCK (F-0), FIRST GENERATION BROODSTOCK (F-1), AND SECOND GENERATION BROODSTOCK (F-2) Tridjoko Balai Besar Riset Perikanan Budidaya Laut – Gondol, Email: [email protected] ABSTRACT Grouper seeding technology still relies on the broodstock from the ocean, eventhough, the existence of the broodstock is rare and difficult to obtain. The broodstock takes a long time to mature and spawn. Therefore, to anticipate the scarcity humpback grouper from wild, it is important to produce broodfish from aquaculture. This study was aimed to know the reproduction performance of humpback grouper wild broodstock F-0, first generation broodstock (F-1) and second generation broodstock (F-2). The cultured of humback grouper broodstock used three tanks 75 m3 in volume (tank I, II and tank III). Tank I: reared 35 fishes broodstock F-0. Tank II and tank III for reared each 50 fishes F-1 and F-2 broodstock. Food for broodstock from the wild (F-0) was given from fresh trash fish, squid, and added vitamin mixture, vitamin C and vitamin E. Meanwhhile, food given to F-1 and F-2 broodstock was a commercial dry pellet (PG 9-10) that contains following nutrients: protein content of 43%, min. fat of 9%, ash content of 13%, fiber levels of 2%, and moisture content of 12%. Mixture of vitamin C and E were also added. The results showed that the humpback grouper F-0, F-1 and F-2 were cultured in 75 m3 concrete tank was successfully spawn with the fertilization rate of 87%, 78% and 45% respectively. This result proves that the broodstock from F-0 gaved the best results. Keywords: Humpback grouper reproduction, wild broodstock (F-0), first generation (F-1), seccond generation (F-2) ABSTRAK Teknologi pembenihan ikan kerapu bebek masih mengandalkan induk dari laut. Padahal induk dari laut sudah sulit didapatkan dan hanya ada di perairan tertentu saja. Disamping itu induk dari laut diperlukan waktu yang cukup lama untuk bisa matang gonad hingga memijah. Oleh karena itu untuk mengantisipasi kelangkaan induk ikan kerapu bebek hasil tangkapan dari laut perlu diupayakan produksi induk ikan kerapu yang berasal dari budidaya/generasi pertama (F-1) maupun induk generasi ke dua (F-2). Penelitian ini bertujuan untuk melihat keragaan reproduksi induk ikan kerapu bebek F-0, F-1 dan F-2. Bak pemeliharaan induk ikan kerapu bebek menggunakan 3 tangki volume 75 m3 (tangki I, II dan III). Untuk tangki I diisi 35 ekor induk kerapu bebek F-0, sedangkan pada tangki II dan III diisi induk F-1 dan F-2 masing-masing 50 ekor. Pakan induk F-0 yaitu ikan rucah segar, cumi-cumi dan juga ditambahkan vitamin mix, vitamin C dan vitamin E. Sedangkan pakan untuk induk F-1 dan induk F-2 adalah pellet kering komersial (PG 9-10) dengan kandungan nutrisi sebagai berikut: kadar protein min. 43%, kadar lemak min 9%, kadar abu max. 13%, kadar serat max. 2% dan kadar air max. 12%, juga ditambahkan vitamin mix, vitamin C dan vitamin E. Hasil penelitian menunjukkan bahwa induk ikan kerapu bebek F-0, F-1 dan F-2 yang dipelihara pada tangki volume 75 m3 secara terkontrol sudah berhasil memijah dengan tingkat fertilisasi tertinggi 87%, 78% dan 45%. Hal ini membuktikan bahwa induk F-0 memperlihatkan hasil yang terbaik. Kata Kunci: reproduksi, induk kerapu alam (F-0), induk generasi pertama (F-1), induk generasi ke dua (F-2) ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 17 Keragaan Reproduksi Ikan Kerapu Bebek (Cromileptes Altivelis) dari Alam (F-0)... I. PENDAHULUAN Dari beberapa jenis ikan kerapu, ikan kerapu bebek (Cromileptes altvelis) adalah satu diantara jenis ikan keluarga Serranidae yang bernilai ekonomis tinggi, karena banyak diminati oleh konsumen baik sebagai ikan hias maupun sebagai ikan konsumsi. Upaya untuk membudidayakan melalui pembenihan telah dilakukan oleh Balai Besar Riset Perikanan Budidaya Laut, Gondol dan telah berhasil memproduksi benih kerapu bebek dengan kelangsungan hidup yang relatif tinggi (Tridjoko et al., 1999; Sugama et al., 2001). Salah satu faktor yang menjadi kendala dalam usaha pembenihan ikan kerapu khususnya ikan kerapu bebek adalah ketersediaan induk. Selama ini induk ikan kerapu yang dipijahkan berasal dari alam yang biasa ditangkap oleh para nelayan. Untuk memperoleh induk ikan kerapu bebek ini relatif sulit, karena hanya ada pada perairan-perairan tertentu saja. Untuk menanggulangi tantangan tersebut, maka sebagai alternatif sudah saatnya dilakukan kajian dan usaha-usaha untuk menyediakan calon induk untuk dijadikan induk dari hasil budidaya. Dengan tersedianya induk hasil budidaya ini diharapkan dapat diproduksi induk yang berkualitas baik dan tidak terjadi penurunan genetik serta bebas penyakit. Produksi benih di hatcheri sangat dipengaruhi oleh kondisi lingkungan (suhu, salinitas, variasi pakan, kandungan nutrisi, kepadatan larva) dan faktor genetik yang berbeda. Akhir-akhir ini telah banyak dilakukan penelitian mengenai pakan buatan terutama kandungan nutrisinya pada ikan kerapu bebek ( Giri et al., 1999 ; Suwirya et al., 2001; Suwirya et al., 2002). Selanjutnya keragaman genetik ikan perlu dipertahankan dalam proses penggunaan induk dalam perbenihan, karena terjadinya 18 reduksi gen akan mengakibatkan hilangnya sebagian karakter genetik benih turunannya (Gondie et al., 1995; Benzie dan William, 1996; Sugama et al., 1998). Tingginya keragaman genetik ini juga banyak dipengaruhi oleh jumlah induk dalam suatu populasi pembenihan dan juga jumlah induk yang effektif dalam suatu pemijahan. Benih hasil budidaya turunan pertama (F-1) yang dijadikan calon induk untuk pembenihan sangatlah penting untuk diketahui nilai rasio RNA/DNA-nya. Semakin tinggi nilai rasio RNA/DNA pada ikan tersebut, maka kualitas ikan juga semakin baik (Caldarone dan Buckley, 1997; Jung dan Clemmesen, 1997; Seginni dan Chung, 1997; Chicharo et al., 1998). Secara bertahap penelitian induk ikan kerapu bebek telah dimulai dari induk yang berasal dari alam (F-0), kemudian induk ikan hasil budidaya (F1) dan berikutnya induk generasi ke dua (F-2). Penelitian diarahkan untuk menghasilkan induk yang dapat dimanfaatkan untuk produksi benih berkualitas, sehingga tidak lagi tergantung dari hasil tangkapan di laut. II. METODE PENELITIAN Hewan uji yang digunakan adalah induk ikan kerapu bebek F-0, F-1 dan F2. Untuk pemeliharaan induk ikan kerapu bebek menggunakan 3 (tiga) bak beton (bak I, bak II dan bak III) yang berbentuk silinder , vulome 75 m3 dan kedalaman air 2 meter. Pada bak I diisi induk ikan kerapu bebek F-0 sebanyak 35 ekor, sedangkan pada bak II dan bak II diisi induk F-1 dan F-2 masing-masing 50 ekor. Pada bak pemeliharaan induk dilengkapi dengan airasi sebagai sumber oksigen dan sistem air mengalir dengan laju pergantian air antara 350 – 500%/hari. Pakan yang diberikan untuk http://www.itk.fpik.ipb.ac.id/ej_itkt22 Tridjoko induk F-0 adalah ikan rucah segar, cumicumi dan juga ditambahkan vitamin mix, vitamin C dan vitamin E. Sedangkan pakan untuk induk F-1 dan induk F-2 adalah pellet kering komersial (PG 9-10) dengan kandungan nutrisi sebagai berikut: kadar protein min. 43%, kadar lemak min 9%, kadar abu max. 13%, kadar serat max. 2% dan kadar air max. 12%, juga ditambahkan vitamin mix, vitamin C dan vitamin E. Bila pada bak pemeliharaan induk ikan kerapu terlihat adanya sisa makanan atau kotoran, maka dilakukan penyiponan. Selanjutnya antara 3 - 5 minggu sekali induk direndam air tawar selama 5 - 10 menit untuk menghilangkan parasit dan bak pemeliharaan induk dibersihkan untuk menghindari dan menanggulangi adanya berbagai kemungkinan serangan penyakit. Pada pipa pembuangan air dipasang jaring kecil berbentuk segi empat dengan ukuran panjang 1 meter, lebar 75 cm dan tinggi 60 cm sebagai tempat penampungan telur yang terbuat dari nylon monofilamen dengan ukuran mata jaring 400 m. Untuk mengetahui produksi telur ikan kerapu bebek, maka setiap pagi jam 07:30 WITA dilakukan pengamatan pada tempat penampungan telur . Selanjutnya dilihat kualitas dan kuantitas telur tersebut, yaitu jumlah telur yang mengapung, jumlah telur yang tenggelam, jumlah total telur yang dihasilkan, diameter telur, jumlah gelembung minyak pada telur, diameter gelembung minyak dan daya tetas telur. III. HASIL DAN PEMBAHASAN Hasil pengamatan pemijahan induk ikan kerapu bebek dari alam (F-0) yang dipelihara pada tangki volume 75 m3 (tangki I) selama 1 tahun dari bulan Januari sampai dengan Desember, tertera pada Tabel (1). Dari 35 ekor induk kerapu bebek dengan bobot tubuh antara 1800–3800 gram/ekor telah berhasil memijah sebanyak 33 kali. Jumlah total telur yang dihasilkan mencapai 15.349.000 butir telur. Dari jumlah total telur yang didapatkan nampaknya telur yang dibuahi berjumlah 8.350.000 butir dan daya tetasnya antara 15-85%. Sedangkan telur yang tidak dibuahi sebanyak 6.995.000 butir. Dari hasil-hasil penelitian mengenai pemijahan, bahwa telur yang mempunyai daya tetas rendah atau dibawah 35% akan berpengaruh terhadap pertumbuhan larva. Hal tersebut berakibat pertumbuhan larva lambat, dan sering terjadi kematian massal atau tidak sampai umur 45 hari. Seperti halnya yang terjadi pada ikan kerapu batik (Slamet dan Tridjoko, 1997), kerapu bebek (Tridjoko et al., 1999), dan juga pada ikan laut lainnya seperti ikan napoleon (Slamet et al., 1998), ikan cobia (Priyono et al., 2005). Dari 33 kali pemijahan, terlihat bahwa pada bulan April, Mei dan Juni tidak terjadi pemijahan (Gambar 1). Pada bulan Maret, Juli, Agustus dan September jumlah telur yang dihasilkan masing-masing 699.000, 3.350.000, 3.325.000 dan 2.825.000 butir. Sedangkan jumlah telur terbanyak terjadi pada bulan Oktober yaitu 5.150.000 butir. Beberapa hasil penelitian menunjukkan bahwa pemijahan ikan-ikan laut ekonomis penting pada umumnya jumlah telur yang dihasilkan setiap bulannya tidaklah stabil melainkan terjadi fluktuasi. Seperti halnya yang terjadi pada ikan bandeng (Vanstone et al., 1977), ikan kerapu macan (Setyadharma et al., 2003), dan ikan kerapu sunu (Suwirya et al., 2003). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 19 Keragaan Reproduksi Ikan Kerapu Bebek (Cromileptes Altivelis) dari Alam (F-0)... Tabel 1. Hasil pengamatan pemijahan induk ikan kerapu bebek dari alam (F-0) selama penelitian berlangsung. ______________________________________________________________________ Parameter Hasil Pengamatan (Tangki I) ______________________________________________________________________ Jumlah Induk (ekor) 35 Kisaran bobot tubuh (g) 1900-3800 Pemijahan (kali) 33 Jumlah total telur (butir) 15.349.000 Telur yang mengapung/dibuahi (butir) 8.350.000 Telur yg mengendap/tdk dibuahi (butir) 6.995.000 Diameter telur (m) 820-915 Diameter gelembung minyak (m) 158-215 Daya tetas telur (%) 15-87 ______________________________________________________________________ 6.000.000 Jumlah telur (butir) 5.000.000 4.000.000 3.000.000 2.000.000 1.000.000 0 Mar Apr Mei Jun Jul Agt Sep Okt Bulan Gambar 1. Jumlah total telur yang dihasilkan dari pemijahan induk ikan kerapu bebek dari alam (F-0). Selanjutnya hasil pengamatan pemijahan induk ikan kerapu bebek generasi pertama (F-1) yang dipelihara pada tangki volume 75 m3 (tangki II) selama 8 bulan dari bulan Januari sampai dengan Agustus, tertera pada Tabel (2). Dari 50 ekor induk kerapu bebek dengan bobot tubuh antara 1150– 18500 gram/ekor telah berhasil memijah sebanyak 27 kali. Jumlah total telur yang dihasilkan mencapai 5.642.000 butir telur. Dari jumlah total telur yang didapatkan nampaknya telur yang 20 dibuahi berjumlah 1.154.000 butir dan yang tidak dibuahi 4.488.000 butir. Diameter telur 799 – 818 m , sedangkan daya tetas telur antara 10-78%. Induk ikan kerapu bebek generasi pertama ini memijah setiap bulan, dan frekuensi pemijahan sebanyak 27 kali (Gambar 2). Jumlah total telur tertinggi yang dihasilkan selama pemijahan terjadi pada bulan Agustus yaitu sebanyak 1.045.000 butir, sedangkan yang terrendah pada bulan Januari yaitu 260.000 butir. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Tridjoko Tabel 2. Hasil pengamatan pemijahan induk ikan kerapu bebek generasi pertama (F-1) selama penelitian berlangsung ______________________________________________________________________ Parameter Hasil Pengamatan (Tangki II) ______________________________________________________________________ Jumlah Induk (ekor) 50 Kisaran bobot tubuh (g) 1150-1850 Pemijahan (kali) 27 Jumlah total telur (butir) 5.642.000 Telur yang mengapung/dibuahi (butir) 1.154.000 Telur yg mengendap/tdk dibuahi (butir) 4.488.000 Diameter telur (m) 799-818 Diameter gelembung minyak (m) 162-165 Daya tetas telur (%) 10-78 ______________________________________________________________________ Jumlah telur (butir) 1200000 1000000 800000 600000 400000 200000 0 Jan Peb Mar Apr Mei Jun Jul Agt Bulan pemijahan Gambar 2. Jumlah total telur yang dihasilkan dari pemijahan induk ikan kerapu bebek generasi pertama (F-1) Jumlah telur yang dihasilkan dari pemijahan induk kerapu bebek generasi pertama lebih sedikit jika dibandingkan dengan induk ikan kerapu bebek tangkapan di alam. Hal tersebut diduga bahwa ukuran induk dari alam lebih besar, disamping faktor lainnya seperti umur ikan dan lain-lain (Tridjoko et al., 1996). Dari bulan Januari sampai dengan bulan April jumlah total telur yang dihasilkan terus mengalami trend kenaikan, dan turun lagi pada bulan Mei. Pada bulan berikutnya Mei, naik lagi hingga bulan Agustus, seperti yang tertera pada Gambar (2). Berikutnya hasil pengamatan pemijahan induk ikan kerapu bebek generasi ke dua (F-2) yang dipelihara pada tangki volume 75 m3 (tangki III) selama 8 bulan dari bulan Januari sampai dengan Agustus, tertera pada Tabel (3). Dari 50 ekor induk kerapu bebek F-2 dengan bobot tubuh antara 850– 1200 gram/ekor telah berhasil memijah sebanyak 36 kali. Jumlah total telur yang dihasilkan mencapai 12.800.000 butir Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 21 Keragaan Reproduksi Ikan Kerapu Bebek (Cromileptes Altivelis) dari Alam (F-0)... telur. Dari jumlah total telur yang didapatkan, telur yang dibuahi berjumlah 1.497.000 butir dan yang tidak dibuahi 11.303.000 butir. Diameter telur 710 – 861 m, sedangkan daya tetas telur antara 0-55%. Dari 36 kali pemijahan, terlihat bahwa dari bulan Maret sampai dengan bulan Oktober terjadi pemijahan dan (Gambar 3). Jumlah telur terbanyak terjadi pada bulan Juni yaitu 2.285.000 butir. Dari hasil pengamatan pemijahan induk kerapu bebek F-2 yang masih belum stabil dan masih relatif sedikit, diduga bahwa jumlah individu jantan yang siap membuahi masih belum memadai. Hal tersebut berkaitan dengan perkembangan gonad yang secara alami membutuhkan waktu yang cukup lama terutama bagi induk kerapu bebek jantan, karena sifatnya yang "protogynous hermaphrodit". Sifat tersebut dimana betina dewasa akan mengalami perubahan kelamin menjadi jantan (Mishima dan Gonzares, 1994). Namun perubahan kelamin ikan kerapu tergantung pada ukuran, umur dan jenisnya. Oleh karena itu dalam manejemen induk kerapu bebek hasil budidaya F-1 maupun F-2 agar cepat mendapatkan induk jantan diperlukan rekayasa hormonal. Tabel 3. Hasil pengamatan pemijahan induk ikan kerapu bebek generasi pertama (F-2) selama penelitian berlangsung ______________________________________________________________________ Parameter Hasil Pengamatan (Tangki III) ______________________________________________________________________ Jumlah Induk (ekor) 50 Kisaran bobot tubuh (g) 850-1200 Pemijahan (kali) 36 Jumlah total telur (butir) 12.800.000 Telur yang mengapung/dibuahi (butir) 1.497.000 Telur yg mengendap/tdk dibuahi (butir) 11.303.000 Diameter telur (m) 710-861 Diameter gelembung minyak (m) 147-175 Daya tetas telur (%) 0-45 ______________________________________________________________________ Jumlah telur (butir) 3000000 2500000 2000000 1500000 1000000 500000 0 Mar Apr Mei Jun Jul Bulan pemijahan Agt Sep Okt Gambar 3. Jumlah total telur yang dihasilkan dari pemijahan induk ikan kerapu bebek turunan ke-2 (F-2) 22 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Tridjoko Umur (hari) Rekayasa hormonal merupakan salah satu alternatif dalam menunjang atau mempercepat proses perkembangan oosit. Untuk perkembang biakan secara buatan tersebut dapat dilakukan dengan cara implantasi (Vanstone et al., 1977; Crim, 1985; Lee et al., 1986 ; Kuo et al., 1988; Tridjoko et al., 1997). Penelitian implan hormon juga telah dilakukan pada beberapa jenis ikan, seperti pada ikan bandeng ( Prijono et al, 1990), ikan kerapu macan (Setiadharma et al, 2001) ikan napoleon (Slamet et al., 1999) ikan kerapu sunu (Suwirya et al, 2005). Dari hasil pemijahan induk kerapu bebek F-0, F-1 dan F-2, maka dilakukan pengamatan pemeliharaan larva. Pertumbuhan larva hingga mencapai umur 25 hari , seperti yang tertera pada Gambar (4) berikut ini : D-20 F-3 F-2 D-10 F-1 D-1 0 5 10 Panj. total larva (mm) Gambar 4. Pertumbuhan panjang total larva ikan kerapu bebek F-1, F-2 dan F-3 yang dipelihara selama 25 hari (D-25) IV. KESIMPULAN Hasil penelitian menunjukkan bahwa induk ikan kerapu bebek F-0, F-1 dan F-2 yang dipelihara pada tangki volume 75 m3 secara terkontrol sudah berhasil memijah dengan tingkat fertilisasi tertinggi 87%, 78% dan 45%. Hal ini membuktikan bahwa induk F-0 memperlihatkan hasil yang terbaik. UCAPAN TERIMA KASIH Penulis mengucapkan terima kasih kepada Bpk/Ibu : Bagus Winaya, M. Rivai, dan semua kelompok Peneliti/ Teknisi serta para Siswa/Mahasiswa Praktek Kerja Lapangan/Magang yang telah membantu selama penelitian ini berlangsung. DAFTAR PUSTAKA Benzie,J.A.H. and S.T.W. Williams. 1996. Limitation of the genetic variation of hatchery produced batches of Giant Clam, Tridacna gigas. Aquaculture, 139:225-241. Caldarone, E.M. and L.J. Buckley. 1997. Relationship between RNA/DNA ratio, temperature and growth rate in Atlantic cod larvae. Ichthyoplankton Ecology Fishries Society of the British Isles, 50pp. Chicharo, M.A., L. Chicharo, L. Valdes, E. Lopez-Jamar, and P. Re. 1998. Estimation of starvation and diet variation of the RNA/DNA ratios in Field-caught Sardina pilcardus larvae of the North of Spain. Mar. Ecol. Prog. Ser., 164:273-283. Crim, L.W. 1985. Methods for acute and chronic hormone administration in fish ,p : 1-9 In Proceeding for a workshop held at Tungkang Marine Laboratory Taiwan, April 22-24, 1985. Giri, N.A., K. Suwirya, dan M. Marzuqi. 1999. Kebutuhan protein, lemak, dan vitamin C untuk yuwana ikan kerapu bebek (Cromileptes altivelis). Jurnal Penelitian Perikanan Indonesia, 5(3):38-46 Goundie, C.A., Q. Liu. B.A. Simeo, and K.B. Davis. 1995. Genetic relationship of growth sex and glucose phosphate isomerase-B in channel cat fish. Aquaculture, 138:119-124. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 23 Keragaan Reproduksi Ikan Kerapu Bebek (Cromileptes Altivelis) dari Alam (F-0)... Jung, T. and C. Clemmesen. 1997. Effect of different food organism on the development and nutritional condition of cod larvae (Gadus morhua L.) in laboratory rearing experiment. Ichthyoplankton Ecology Fisheries Society of the British Isles, 50pp. Kuo, C.M., Y.Y. Thing, and S.L. Yeh. 1988. Induced sex reversal and spawning of spotted grouper, E. fario. Aquaculture, 74:113-126. Lee, C.S., C.S. Tamaru, J.E. Banno, and C.D. Kelly. 1986. Influence of administration of LHRH analoque and/or 17 methyltestoteron on maturation in milkfish, Chanos chanos. Aquaculture, 59:147-159. Mishima, H. and B. Gonzaes. 1994. Some biological and ecological aspect on Cromileptes altivelis around Palawan Island, Philippines. Suisanzoshoku, 42(2):345-349 Prijono, A., G. Sumiarsa, dan Z.I. Azwar. 1990. Implantasi hormon LHRH-a dan atau 17a MT untuk pematangan gonad induk bandeng. J. Penel. Budidaya Pantai, 6(1):2023 Priyono. A., B. Slamet, dan Asmanik. 2005. Pengamatan pemijahan alami di bak pemeliharaan, perkembangan embrio dan awal larva ikan cobia (Rachycentron canadum). Seminar akuakultur Indonesia, Hotel Sahid Jaya, Makasar, 23-25 Nopember 2005. 12 hal. Seginni, M.I., and K. S. Chung. 1997. Influence of environmental factors on the instant growth of tropical fishes assessed by the RNA/DNA relationship. Biol. Inst. Oceanogr. Venez., 36(1/2):21-29 Setiadharma,T., N.A. Giri, Wardoyo, dan A. Prijono. 2001. Pembenihan Ikan Kerapu Macan (Epinephelus fuscoguttatus) ). Pros. Lokakarya 24 Nasional Pengembangan Agribisnis Kerapu hal 165-174. Jakarta 28-29 Agustus 2001. Setiadharma, T., A. Prijono, dan N.A Giri. 2003. Aplikasi hormon LHRH-a untuk meningkatkan pemijahan dan kualitas telur induk ikan kerapu macan (Epinephelus fuscoguttatus). Laporan Teknis Proyek Riset Perikanan Budidaya Laut, Gondol Bali. P.101-109. Slamet, B., Hersapto, dan Tridjoko. 1999. Pematangan induk ikan napoleon Cheilinus undulatus dengan perbandingan pakan segar yang berbeda. Seminar Nasional Penelitian dan Deseminasi teknologi Budidaya Laut dan Pantai. Jakarta, 2 desember 1999. Slamet, B. dan Tridjoko. 1997. Pengamatam pemijahan alami, perkembangan embrio dan larva ikan kerapu batik, Epinephelus microdon dalam bak terkontrol. J. Pen. Perikanan Indonesia, 3(4):4050. Slamet, B., Hersapto, dan Tridjoko. 1998. Pengamatan panjang-bobot, kebiasaan makan dan aspek biologi reproduksi ikan napoleon, Cheilinus undulatus. Prossiding Seminar Teknologi Perikanan Pantai. Bali, 6-7 Agustus 1998: 119-123. Sugama, K., Tridjoko, Haryanti, S.B. Moria, dan F. Cholik. 1998. Genetic variation and population stucture in the Humback grouper, Cromileptes altivelis throughout its range in Indonesian waters. Indonesian Fisheries Research Journal, 5(1)1:32-38 Sugama, K., Tridjoko, B. Slamet, S. Ismi, E. Setadi, dan S. Kawahara. 2001. Petunjuk teknis produksi benih ikan kerapu bebek, Cromileptes altivelis. BBRPBL, Pusris., DKP dan JICA. 40p http://www.itk.fpik.ipb.ac.id/ej_itkt22 Tridjoko Suwirya, K., N.A. Giri, dan M. Marzuqi. 2001. Pengaruh n-3 HUFA terhadap pertumbuhan dan efisiensi pakan yuwana ikan kerapu bebek, Cromileptes altivelis. In Sudrajad, A., E.S. Heruwati, A. Poernomo, A. Rukyani, J. Widodo, dan E. Danakusuma (Eds) Teknologi Budidaya Laut dan Pengembangan Sea Farming di Indonesia. Departemen Kelautan dan Perikanan p.201-206. Suwirya, K., N.A. Giri, M. Marzuqi, dan Tridjoko. 2002. Kebutuhan karbohidrat untuk pertumbuhan yuwana ikan kerapu bebek, Cromileptes altivelis. JPPI, Edisi Akuakultur, 8:9-14. Suwirya,K., A. Prijono, N.A Giri, B. Slamet, dan Marzuqi. 2003. Pematangan induk induk kerapu sunu, Plectropomus leopardus dengan penambahan vitamin C pada pakan. Laporan Teknis Proyek Riset Perikanan Budidaya Laut, Gondol Bali. hal. 314-223 Suwirya, K., A. Priyono, M. Marzuqi, N.A. Giri, dan R. Andamari. 2005. Pemijahan dan pemeliharaan larva kerapu sunu halus, Plectropomus leopardus. Warta Penelitian Perikanan Indonesia. Edisi Akuakultur, 2(3):7-10. Tridjoko, B. Slamet, D. Makatutu, dan K. Sugama. 1996. Pengamatan pemijahan dan perkembangan telur ikan kerapu bebek (Chromileptes altivelis) pada bak secara terkontrol. J. Penel. Perikanan Indonesia, 2(2):55-62. Tridjoko, B. Slamet, dan D. Makatutu. 1997. Pematangan induk kerapu bebek (Cromileptes altivelis) dengan rangsangan suntikan hormon LHRH-a dan 17 alphamethyltestoteron. J. Penel. Perikanan Indonesia, 3(4):30-34. Tridjoko, B. Slamet., T. Aslianti, Wardoyo, S. Ismi, J.H. Hutapea, K.M. Setiawati, I. Rusdi, D. Makatutu, A. Prijoni, T. Setiadharma, H. Matsuda, and S. Kumagai. 1999. The seed production technique of humback grouper, Cromileptes altivelis. Japan International Cooperation Agency (JICA) and Gondol Research Station For Coastal Fisheries (GRSCF). 56p. Vanstone, W.E., Tiro, Jr., L.B. Villaluz, A.C. Ramsingh, D.C. Kumagai, S. Dulduco, P.T. Barnes, M.M. L., and C.E. Duenas. 1977. Breeding and Larval rearing of the milkfish Chanos-chanos (Pisces Chanidae) SEAFDEC, Aquculture Deparment Tech. Report, 3:3-17. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 25 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 26-33, Desember 2010 PEMELIHARAAN GELONDONGAN KERAPU SUNU (Plectropomus leopardus) DENGAN PERSENTASE PERGANTIAN AIR YANG BERBEDA DIFFERENT PERCENTAGE OF WATER EXCHANGE ON GROWTH OF CORAL TROUT GROUPER FINGERLING (Plectropomus leopardus) Titiek Aslianti Balai Besar Riset Perikanan Budidaya Laut, Gondol PO Box 140 Singaraja 81101, Bali E-mail: [email protected] ABSTRACT Coral trout grouper (Plectropomus leopardus) is the prospective aquaculture commodities with a high demand. Fingerling culture is the one of shortly rearing time methods for grow out in net-cages in order to produce continuously. Two hundred fishes with initial total length 16.6±0.5 cm and 72.2±7.6 g of body weight were stocked in each of three of concrete tanks of 4m3 seawater. Water replace as a treatment was applied i.e. A (200%/day), B (300%/day) and C (400%/day).respectively. Feeding frequency of pellet was done twice a day of about 3 - 5% of body weight. Survival and growth of fishes were monitored during two-month period. The result showed that the different percentage of water replace had a significant role in increasing the growth and survival rate of the fishes. Water replaced of 400% daily (treatment C) gave the best survival rate (97%) and growth performance (TL 21.61±0,54 cm; BW 156.84±1.05 g). In contrast, the treatment B (300%/day) and A (200%/day) resulted in a lower survival and growth , i.e. (SR 95.5%; TL 19.51±0.52 cm; BW 140.96±0.08 g) and A (SR 93%; TL 19.08±0.30 cm; BW 132.2±2.65 g)., respectively. Keywords: Coral trout grouper, growth, survival rate, water exchange ABSTRAK Ikan kerapu sunu (Plectropomus leopardus) termasuk komoditas budidaya yang prospektif dengan pangsa pasar cukup tinggi. Upaya penggelondongan merupakan salah satu cara untuk mempersingkat waktu pemeliharaan di keramba jaring apung (KJA) agar kontinuitas produksi dapat terpenuhi sesuai permintaan pasar. Penelitian dilakukan selama 2 bulan dengan menggunakan 3 unit bak fiber (4m3), masing-masing diisi 200 ekor kerapu sunu yang berukuran awal rata-rata panjang total (TL) 16.6±0.5cm dan berat tubuh (BW) 72.2±7.6g. Pergantian air pemeliharaan merupakan perlakuan yaitu A (200%/hari), B (300%/hari) dan C (400%/hari). Pakan berupa pellet komersial diberikan dua kali sehari 3-5% dari bobot biomas. Hasil penelitian menunjukkan bahwa perbedaan persentase pergantian air memberikan pengaruh yang berbeda terhadap peningkatan pertumbuhan dan kelangsungan hidup ikan. Pergantian air 400%/hari (C) menghasilkan kelangsungan hidup (97%) dan pertumbuhan (TL 21.61±0,54cm; BW 156.84±1.05g) paling baik, sedangkan perlakuan B (300%/hari) dan A (200%/hari) menghasilkan kelangsungan hidup dan pertumbuhan lebih rendah dari pada C yaitu berturut-turut B (SR 95.5%; TL 19.51 ±0.52cm; BW 140.96±0.08g) dan A (SR 93%; TL 19.08±0.30cm; BW 132.2±2.65g). Kata Kunci: Ikan kerapu sunu, pertumbuhan, laju kelangsungan hidup, pergantian air 26 ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Aslianti I. PENDAHULUAN Ikan kerapu sunu (Plectropomus leopardus) yang dikenal dengan kerapu bintang termasuk satu diantara komoditas ekspor unggulan Indonesia dari budidaya laut (marine fin-fish culture). Warna merah pada kerapu sunu merupakan daya tarik tersendiri bagi beberapa negara importir seperti Hongkong dan China, yang sebagian besar masyarakatnya masih meyakini bahwa warna merah identik dengan keberuntungan, sehingga pangsa pasar kerapu sunu di kedua negara tersebut sangat tinggi dan merupakan negara tujuan ekspor yang potensial (Sidik, 2002; Nurjana, 2006). Kerapu sunu dalam budidayanya memiliki prospek pengembangan yang sangat baik, karena tehnik produksi benih secara masal telah dikuasai dan dapat diterapkan di hatchery skala rumah tangga (Aslianti et al., 2009a). Permintaan pasar terhadap kerapu sunu terutama dalam keadaan hidup sangat tinggi (Sutarmat et al., 2007) dan terus meningkat, sementara pemenuhan melalui penangkapan di alam sangat tidak disarankan, karena memiliki banyak resiko yang merugikan diantaranya adalah jumlah tidak kontinyu, ukuran tidak seragam, resiko cacat fisik saat penangkapan dan dampak tertinggi adalah berkurangnya populasi di alam. Hal ini memberikan peluang cukup besar bagi kegiatan budidaya sekaligus meningkatkan pasok yuwana yang kontinyu, namun faktor kualitas tetap menjadi prioritas yang perlu diperhatikan mengingat persaingan pasar yang semakin ketat (Nurjana, 2010). Dalam usaha pembesaran kerapu sunu di Keramba Jaring Apung (KJA) hingga mencapai ukuran konsumsi (300400gr) biasanya memerlukan waktu pemeliharaan yang cukup lama dan memiliki resiko kematian yang tinggi. Oleh karenanya upaya penggelondongan perlu dilakukan agar waktu pemeliharaan di KJA dapat dipersingkat dan diperoleh produksi yang tinggi dengan kualitas dan tampilan yang proporsional. Langkah awal yang harus dilakukan dalam mempersiapkan yuwana sebagai benih untuk pembesaran di KJA antara lain ukuran yang seragam, tidak cacat tubuh dan ikan sudah terbiasa mengkonsumsi pakan buatan (Sutarmat, 2005). Uji coba pendederan dalam wadah terkontrol dengan berbagai ukuran hingga pembesarannya di KJA telah dilakukan Sutarmat dan Ismi (2007), namun tingkat kelangsungan hidup yang dihasilkan belum stabil (Suwirya et al., 2006), dan masih perlu dilakukan perbaikan yang mengarah pada peningkatan produksi Dalam kegiatan budidaya, selain pakan yang memerlukan biaya ±60% dari total biaya operasional, faktor lingkungan juga merupakan sarana yang tidak kalah pentingnya, mengingat air sebagai media tempat ikan dipelihara juga merupakan media tumbuh bagi berbagai macam mikroorganisme baik yang pathogen maupun nonpathogen. Sedangkan di alam, golongan ikan kerapu umumnya hidup di perairan karang, bersifat menyendiri dan lebih menyukai habitat yang bersih dengan kondisi perairan yang stabil (Aslianti et al., 1998). Mengantisipasi sifat alami ikan kerapu tersebut, pergantian air merupakan faktor penting dalam menjaga kestabilan kualitas air selama pemeliharaan dalam wadah terkontrol. Oleh karenanya pengelolaan lingkungan pemeliharaan kerapu sunu melalui pergantian air yang kontinyu dengan prosentase yang optimal diharapkan dapat mengurangi kendalakendala ekstrim yang terjadi. Dengan melihat permasalahan yang cukup konkrit dan tuntutan peluang pasar yang memerlukan solusi secara cepat, maka perlu dilakukan penelitian penggelondongan kerapu sunu dalam wadah terkontrol melalui perbaikan Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 27 Pemeliharaan Gelondongan Kerapu Sunu (Plectropomus Leopardus)… lingkungan sampai mencapai ukuran siap tebar (± 150gr) sehingga masa pemeliharaan di KJA dapat dipersingkat. Penelitian bertujuan untuk mendapatkan informasi pemeliharaan gelondongan kerapu sunu dalam wadah terkontrol, serta kendala yang dihadapi selama pemeliharaan sehingga resiko yang mungkin terjadi pada budidaya pembesaran di KJA dapat diperkecil. II. BAHAN DAN METODE 2.1. Persiapan wadah Kegiatan penelitian diawali dengan pencucian wadah penelitian berupa tiga (3) unit bak fiber kapasitas 4 m3 berbentuk segi empat berukuran 2x2x1m, dengan cara menyikat dan menyemprot dengan air tawar, kemudian dikeringkan selama satu hari. Hal ini untuk menghindari kemungkinan kontaminasi parasit. Selanjutnya wadah diisi air laut bersalinitas 30-33 ppt sampai ketinggian air 75 cm. Pengaturan pipa dan batu aerasi disesuaikan dengan kebutuhan dan diatur sedemikian rupa sehingga pasok oksigen yang mengalir dalam bak diharapkan dapat memenuhi kebutuhan ikan. Demikian juga pengaturan debit air masuk (inlet) dan keluar (outlet) disesuaikan dengan perlakuan dengan cara terlebih dahulu memposisikan stop kran yang disesuaikan dengan jumlah air masuk per menit, sehingga dalam waktu 24 jam (sehari semalam) diharapkan air dapat terganti sesuai perlakuan. 2.2. Hewan uji. Hewan uji (kerapu sunu) yang telah melalui seleksi (keseragaman ukuran dan kondisi fisik) dimasukkan kedalam masing-masing bak dengan kepadatan 200 ekor. Rata-rata panjang total (TL) dan berat tubuh (BW) awal adalah 16.6±0.5cm dan 72.2±7.6g. Selama pemeliharaan, ikan diberi pakan komersial berbentuk pellet berdiameter ±5 mm dengan kandungan protein 42%, 28 lemak 10%, abu 13%, dan kadar air 10% (Aslianti et al., 2009b). Pakan diberikan dua kali sehari sebanyak 3-5% dari bobot biomasa (±500 gr) dengan cara menebar hingga ikan tidak merespon (ad-libitum). Sisa pakan ditimbang untuk mengetahui jumlah pakan yang terkonsumsi setiap hari serta dilakukan penghitugan pada akhir penelitian untuk mengetahui nilai rasio konversi pakannya. Penelitian dilakukan selama 2 bulan dengan menggunakan sistim air mengalir dan persentase pergantian air merupakan perlakuan yaitu A (200%/hari); B (300%/hari) dan C(400%/hari). Penyiponan dasar bak dilakukan setiap 2 hari untuk membersihkan sisa metabolisme berupa faeses ataupun sisa pakan yang tidak terkonsumsi oleh ikan. 2.3. Parameter yang diamati Pengamatan pertumbuhan dilakukan setiap bulan melalui pengukuran panjang total dan berat tubuh terhadap 20 ekor sampel (10% dari jumlah biomas) yang diambil secara acak dari masingmasing perlakuan. Untuk mengetahui efektifitas pakan terhadap pertumbuhan dilakukan dengan cara menghitung rasio konversi pakan (FCR), sedangkan kualitas air diamati secara kontinyu setiap minggu sebagai data pendukung meliputi oksigen terlarut (DO), ammonia (NH3-N), nitrit (NO2-N) dan pH. 2.4. Analisa data Semua data yang diperoleh dihimpun secara tabulasi dan dianalisis secara diskriptif serta diolah dengan menggunakan program microsoft Excel. III. HASIL DAN PEMBAHASAN Hasil pengamatan terhadap pertumbuhan panjang dan berat mutlak, kelangsungan hidup dan rasio konversi pakan gelondongan kerapu sunu dari masing-masing perlakuan tertera pada Tabel 1. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Aslianti Dari Tabel 1. diketahui bahwa persentase kenaikan pertumbuhan perlakuan C baik terhadap panjang total (30.18%) maupun berat tubuh (117.23%) terlihat lebih tinggi dari pada perlakuan A (14.94% dan 83.10%) maupun B (17.53% dan 95.24%). Demikian juga tingkat kelangsungan hidup yang dicapai perlakuan C lebih tinggi (97%) dari pada perlakuan A (93%) ataupun B (95,5%). Hal ini menunjukkan bahwa pergantian air sebesar 400% setiap hari terbukti sangat mendukung pertumbuhan maupun kelangsungan hidup kerapu sunu. Makin tinggi persentase pergantian air dalam penelitian ini mempunyai dampak yang positip terhadap kestabilan kualitas air pemeliharaan. Kondisi air yang kotor akibat terakumulasinya sisa metabolisme ataupun kotoran lain yang terdapat dalam bak, akan terganti secara cepat jika persentase pergantian air cukup tinggi, sehingga diprediksi dapat menghambat tumbuhnya parasit (jamur ataupun bakteri). Kondisi ini didukung dari hasil pengamatan kualitas air yang dilakukan setiap tiga hari yang menunjukkan bahwa kandungan nitrit dan ammonia cenderung menurun dengan meningkatnya persentase pergantian air (Tabel 2). Tabel 1. Pertumbuhan panjang dan berat mutlak, kelangsungan hidup dan rasio konfersi pakan kerapu sunu dari masing-masing perlakuan Parameter Panjang total awal/Initial length (cm) Panjang total akhir/Final length (cm Pertambahan panjang/TL gain (%) Berat tubuh awal/Initial weight (g) Berat tubuh akhir/Final weight (g) Pertambahan berat/BW gain (%) Kelangsungan hidup/SR (%) Rasio konfersi pakan/FCR A B C (200%/day) (300%/day) (400%/day) 16.60 16.60 16.60 19.08 19.51 21.61 14.94 17.53 30.18 72.20 72.20 72.20 132.20 140.96 156.84 83.10 95.24 117.23 93 95.5 97 1.05 1.21 1.34 Tabel 2. Hasil rata-rata pengamatan kualitas air yang dilakukan setiap tiga hari dari masing-masing perlakuan Parameter Suhu maximum/Temp. max (oC) Suhu minimum/Temp. min (oC) Salinitas/Salinity (ppt) pH DO/Disolve Oksigen (mg/L) NO2/Nitrite (mg/L) NH3/Amonium (mg/L) A B C (200%/day) (300%/day) (400%/day) 28.50 ± 0.16 28.28 ± 0.19 27.96 ± 0.23 26.64 ± 0.21 26.98 ± 0.45 27.30 ± 0.42 32.4 ± 1.61 32.4 ± 1.61 32.40 ± 1.61 7.78 ± 0.30 7.51 ± 0.23 7.05 ± 0.10 6.39 ± 0.13 6.72 ±0.31 7.10 ± 0.29 1.50 ± 0.25 1.26 ± 0.04 0.69 ± 0.13 1.83 ± 0.06 1.57 ± 0.08 0.98 ± 0.05 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 29 Pemeliharaan Gelondongan Kerapu Sunu (Plectropomus Leopardus)… Tingginya kandungan nitrit dan ammonia pada perlakuan A dan B diduga sebagai akibat adanya proses pembusukkan serta penguraian sisa metabolisme yang mengendap di dasar bak. Pengendapan terjadi akibat jumlah pergantian air yang kurang mencukupi sehingga badan air tidak cukup mampu mendorong sisa metabolisme ke-arah saluran pembuangan. Kondisi ini akan semakin buruk apabila air laut yang mengalir saat itu dalam keadaan keruh yang biasa terjadi akibat adanya hujan. Mekanisme toksisitas dari nitrit dapat berpengaruh terhadap transportasi oksigen dalam darah dan kerusakan jaringan tubuh ikan. Sesuai pendapat Boyd (1990) yang menyatakan bahwa kandungan ammonia dalam media pemeliharaan merupakan hasil metabolisme ikan, pembusukan senyawa organik dan bakteri. Amonia adalah hasil utama penguraian protein dan merupakan racun bagi ikan, karenanya kandungan ammonia dianjurkan tidak lebih dari 1 mg/L (Pescod, 1973 dalam Aslianti , et al. 1998). Tingginya kandungan nitrit dan ammonia pada perlakuan A dan B diduga berpengaruh terhadap kelangsungan hidup ikan sehingga lebih rendah dari pada C. Namun demikian kondisi suhu, salinitas, pH dan DO nampak masih berada dalam batas normal dan masih mendukung kehidupan ikan. Kualitas air yang baik dan stabil sangat berpengaruh terhadap kesehatan ikan yang pada gilirannya dapat meningkatkan kelangsungan hidupnya. Ditinjau dari hubungan partumbuhan panjang dan berat tubuh secara linier (Gambar 1) pada perlakuan C (pergantian air 400%/hari) diperoleh persamaan regresi Y=1.6972X+120.24 dengan R2 = 0.9354. Hal ini menunjukkan bahwa antara pertumbuhan panjang dan berat tubuh kerapu sunu pada perlakuan C terdapat korelasi yang positif, yang berarti bahwa pergantian air sebesar 400%/hari tidak saja berpengaruh terhadap pertumbuhan panjang tetapi juga berpengaruh terhadap pertumbuhan berat dengan tingkat korelasi sebesar 93.54%. Dari hasil pengamatan selama pemeliharaan gelondongan kerapu sunu dalam wadah terkontrol terdapat beberapa ekor ikan yang mengalami kematian. Hingga akhir penelitian kematian paling banyak terjadi pada perlakuan A (14 ekor), sedangkan B (9 ekor) dan C (6 ekor). Dari hasil deteksi laboratorium diketahui bahwa kematian umumnya disebabkan oleh penyakit infeksi bakteri gram negatif yang merupakan penyakit utama kerapu sunu. BERAT TUBUH (GR) 164.0 y = 1.6972x + 120.24 R2 = 0.9354 160.0 156.0 152.0 148.0 19.0 21.0 23.0 25.0 PANJANG TOTAL (CM) Gambar 1. Hubungan pertumbuhan panjang dan berat tubuh kerapu dengan pergantian air 400%/hari (linier) pada akhir penelitian 30 http://www.itk.fpik.ipb.ac.id/ej_itkt22 sunu Aslianti Gejala kematian ditandai dengan ikan tidak nafsu makan dan berenang lemah, cenderung sering berada di permukaan air, menyendiri, dan terdapat luka pada permukaan kulit (Gambar 2a). Infeksi bakteri diduga berasal dari lingkungan pemeliharaan (air) yang juga merupakan media hidup bagi semua mikroorganisme baik yang patogen maupun nonpatogen. Semakin tinggi persentase pergantian air terlihat semakin sedikit jumlah ikan yang mati karena dengan semakin cepat air media terganti maka kemungkinan ikan terinfeksi parasit yang ada dalam air media semakin kecil. Penanggulangan pertama yang dilakukan adalah dengan merendam ikan yang sakit dengan 100 ppm formalin selama 1 jam atau dalam larutan streptomysin 1 gram dalam 100 liter air laut selama 1 jam serta memisahkan ikan yang sakit dari yang lainnya (Sutarmat et al., 2007). Selain itu pada akhir penelitian juga ditemukan beberapa ekor ikan dengan performansi mulut yang abnormal (deformity) yaitu antara mulut bagian atas dan bawah tidak simetris (Gambar 2b). Kondisi ini dapat menyebabkan ikan tidak bisa merespon pakan dengan sempurna yang pada gilirannya dapat mengakibatkan kematian. Sesuai pendapat Kordi (2004), yang menyatakan bahwa ikan yang cacat akan mengalami kesulitan dalam mendapatkan a makanannya, sehingga ikan akan mengalami keterlambatan pertumbuhan (kerdil), memiliki daya tahan tubuh yang rendah dan akan sangat mudah diserang penyakit. Terjadinya performansi yang abnormal belum diketahui penyebabnya secara pasti, namun diduga adanya faktor genetis dari induk sebagai penghasil telur. Oleh karenanya dalam proses pembenihan, tahap seleksi telur sangat penting sebelum dilakukan penebaran dalam wadah pemeliharaan larva. Tingkat pembuahan telur yang optimal dengan daya tetas mencapai 90% umumnya menghasilkan benih yang normal. Namun demikian deformity pada mulut biasanya ditemukan justru setelah ikan bertumbuh menjadi gelondongan. Diduga selain faktor genetis, juga faktor nutrisi pakan selama pemeliharaan kurang mendukung (malnutrition), sehingga dapat memicu terjadinya ketidaknormalan pada mulut. Dengan demikian pemeliharaan gelondongan kerapu sunu dalam wadah terkontrol hingga mencapai ukuran ± 150 gram memberikan dampak positif terhadap tingkat selektifitas performansi ikan sebelum ditebar di KJA. Selain waktu pemeliharaan di KJA hingga mencapai ukuran konsumsi dapat dipersingkat, juga performansi yang dihasilkan sesuai dengan selera pasar. b Gambar 2. Penyakit bakteri gram negatif yang menimbulkan luka pada permukaan tubuh kerapu sunu (a), dan kerapu sunu yang mengalami deformity pada mulut unsimetris (b) Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 31 Pemeliharaan Gelondongan Kerapu Sunu (Plectropomus Leopardus)… IV. KESIMPULAN Pergantian air dalam pemeliharaan gelondongan kerapu sunu secara terkontrol merupakan faktor penting yang mendukung kelangsungan hidup maupun pertumbuhannya. Kelangsungan hidup dan pertumbuhan ikan yang dipelihara dengan pergantian air 400%/hari lebih tinggi (SR 97%; TL 21.61±0,54 cm, BW 156.84±1.05g) dari pada pergantian air 300% (SR 95.5%; TL 19.51±0.52 cm; BW 140.96±0.08 g) ataupun 200% (SR 93%; TL 19.08±0.30 cm; BW 132.2±2.65 g). Pergantian air hingga 400%/hari dapat menghindarkan ikan dari serangan parasit sehingga ikan bertumbuh normal dan memperkecil jumlah ikan yang cacat tubuh (deformity). DAFTAR PUSTAKA Aslianti, T., K.M. Setiawati dan Wardoyo. 1998. Pengaruh Peningkatan Pergantian Air terhdap Pertumbuhan dan Sintasan Larva Kerapu Bebek, Cromileptes altivelis. Prosiding Seminar Teknologi Perikanan Pantai. Denpasar-Bali, 6-7 Agustus 1998. hal 173-177. Aslianti, T., P.T. Imanto, dan M. Suastika. 2009a. Dampak Minyak Buah Merah, Pandanus conoideus Lam pada Performansi Yuwana Kerapu Sunu, Plectropomus leopardus. Jurnal Perikanan, XI(1):1-8. Aslianti T, Afifah dan M. Suastika. 2009b. Pemanfaatan Minyak Buah Merah, Pandanus conoideus Lam dan Carophyll Pink dalam Ransum Pakan Yuwana Ikan Kakap Merah, Lutjanus sebae. Jurnal Riset Akuakultur, 4(2):191-200. 32 Boyd, C.E. 1990. Water Quality in Ponds for Aquaculture. Alabama : Auburn University. 482p. Kordi, M.G.H. 2004. Penanggulangan Hama dan Penyakit Ikan. Jakarta. PT Rineka Cipta dan PT Bina Adikarsa. Hal 68. Nurjana, M.L. 2006. Indonesian Aquaculture Development. Innovative and Eco-friendly Technologies for the Production of Safe Aquaculture Food. Food & Fertilizer Technology Center For The Asian and Pacific Region (FFTC-ASPAC)/RCA. International Workshop. Den-pasar Bali, Indonesia. December 4-8, 2006. pp 81-100. Nurjana, M. L. 2010. Proyeksi produksi perikanan budidaya menurut komoditas utama 2009 s/d 2014. Materi presentasi pada acara Forum Inovasi dan Teknologi Akuakultur. Puriskan Budidaya. Bandar Lampung, 20-24 April 2010. Sidik. 2002. Alternatif Kebijakan Budidaya Ikan Kerapu Masya-rakat Nelayan dalam Pengem-bangan Industri Perikanan Kerapu. Majalah Ilmiah Analisis Sistem, 4(IX):104-109 Sutarmat, T. 2005. Analisis Finansial Produksi Yuwana Kerapu Macan (Epinephelus fuscoguttatus) dengan Pakan Pelet Komersial dan Ikan Rucah dalam Keramba Jaring Apung. Jurnal Perikanan, VII(2):144-150. Sutarmat, T. dan S. Ismi. 2007. Variasi ukuran tubuh benih pada pendederan ikan kerapu sunu (Plectropomus leopardus). Buku Pengembangan Teknologi Budidaya Perikanan. Balai Besar Riset Perikanan Budidaya Laut. Badan Riset Kelautan dan Perikanan. hal 59-63. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Aslianti Sutarmat, T., K. Suwirya, dan N. A. Giri. 2007. Penelitian pendahuluan pembesaran kerapu sunu (Plectropomus leopardus) dalam Keramba Jaring Apung. Buku Pengembangan Teknologi Budidaya Perikanan. Balai Besar Riset Perikanan Budidaya Laut. BRKP. Hal 438-445. Suwirya, K., A. Priyono, A. Hanafi, R. Andamari, R. Melianawati, M. Marzuqi, K. Sugama dan N.A. Giri. 2006. Pedoman Teknis Pembenihan Ikan Kerapu Sunu (Plectropomus leopardus). Pusat Riset Perikanan Budidaya. Badan Riset Kelautan dan Perikanan. 18 hal. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 33 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 34-40, Desember 2010 KARAKTERISTIK SENYAWA BIOAKTIF BAKTERI SIMBION MOLUSKA DENGAN GC-MS CHARACTERISTIC BIOACTIVE COMPOUND OF THE MOLLUSC SYMBIOTIC BACTERIA BY USING GC-MC Delianis Pringgenies Marine Science Departement, Faculty of Fisheries and Marine Science Diponegoro Univeristy, Semarang, e-mail: [email protected] ABSTRACT It has been discovered that mollusca produce a secondary metabolite and in the same time bear its important role in its ecosystems so became a strategic target for the development of noble bioactive substances for marine pharmacology. The current study of mollucs symbiotic bacteria showed that from species of Conus miles, Stramonita armigera, Cymbiola vespertilo and from based on screening of symbiotic bacteria in the Mollusc toward some bacteria , 3 isolates had been had good performance in inhibiting the grow of bacteria and to be the best candidates for a new antibiotic based on result of screening consistency. Size and character inhibiting zone resulted toward test bacteria were TCM, TCAand TOV. The research aims to current study of characteritic of symbiotic bacteria from mollucs that produce a new anti-pathogenic bacteria by using GC-MS method. GCMS result showed that fraction TCM-6.1 consist of some compounds, that are Nitrogen oxide (N2O) (CAS) Nitrous oxide; Acetic acid (CAS) Ethylic acid; Propanoic acid,2methyl-(CAS) Isobutyric acid and fraction TOV12.16 consist of compound such as Propanoic acid,2-methyl-(CAS)Isobutyric acid; Butanoic acid, 2-methyl-(CAS) 2Methylbutanoid acid then fraction TSA8.7 consist of 1,2-Propadiene (CAS) Allene. The research pointed towards the three active symbiotic bacteria seems to be promising since this three candidates potential result for the development of a new antibiotic. Keywords: Bacteria simbiont, mollusc, anti-bacteria, bioaktif compound ABSTRAK Keberadaan bakteri yang berasosiasi dengan moluska laut telah memungkinkan penggunaan organisme tersebut sebagai sumber utama bakteri yang baru dan sumber senyawa bioaktif termasuk senyawa antimikroba. Hasil penelitian sebelumnya dari isolasi bakteri simbion jenis Conus miles, Stramonita armigera, Cymbiola vespertilo dan berdasarkan konsistensi hasil skrining, besar kecilnya zona hambat yang dihasilkan dan sifat penghambatannya terhadap beberapa jenis bakteri uji melalui uji sensitivitas maka dihasilkan 3 isolat bakteri simbion yang memiliki senyawa bioaktif antibakteri yakni: TCM, TCA dan TOV. Tujuan penelitian adalah mengetahui karakteristik senyawa bioaktif bakteri yang berasosiasi dengan Moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo dengan metode GC-MS. Analisis GC-MS dilakukan menggunakan GCMS-QP2010S Shimadzu. Hasil analisis GC-MS menunjukkan bahwa terdapat beberapa senyawa yang terdeteksi dari fraksi isolat TCM6.1, yakni: Nitrogen oxide (N2O) (CAS) Nitrous oxide; Acetic acid (CAS) Ethylic acid; Propanoic acid,2-methyl-(CAS) Isobutyric acid dan fraksi isolat TOV12.16. : Propanoic acid,2-methyl-(CAS)Isobutyric acid; Butanoic acid, 2-methyl(CAS) 2-Methylbutanoid acid sedang fraksi isolat TSA8.7: 1,2-Propadiene (CAS) Allene. Hasil penelitian disimpulkan bahwa isolat bakteri aktif yang diperoleh dalam penelitian ini menunjukkan hasil yang sangat menjanjikan. Kata Kunci: Bakteri simbion, moluska, anti-bakteri, senyawa bioaktif 34 ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Pringgenies I. PENDAHULUAN Lautan merupakan sumber dari kelompok besar kimia bahan hayati laut dengan struktur yang unik, yang terutama terakumulasi pada hewan-hewan avertebrata yang banyak terdapat pada ekosistem terumbu karang, seperti, sponge, tunikata, bryozoa, karang lunak dan moluska. Beberapa metabolit sekunder yang dimiliki avertebrata laut tersebut menunjukkan adanya aktifitas farmakologi dan merupakan kandidatkandidat baru untuk bahan obat-obatan. Sejumlah senyawa bioaktif yang diperoleh dari hewan avertebrata diduga dihasilkan juga oleh mikroorganisme yang berasosiasi dengannya. Seperti dinyatakan oleh Watermann, 1999; Burgess et al. (2003) bahwa mikroorganisme yang berasosiasi dengan organisme laut akan mensistesa metabolit sekunder seperti organisme inangnya. Keberadaan bakteri yang berasosiasi dengan moluska laut telah memungkinkan penggunaan organisme tersebut sebagai sumber utama bakteri yang baru. Bukti-bukti ilmiah menunjukkan bahwa bakteri yang berasosiasi dengan avertebrata filum Moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo mempunyai peranan dalam produksi senyawa bioaktif, sehingga menjadi pemacu dalam pencarian senyawa antimikroba dari bakteri yang berasosiasi dengan Moluska (Pringgenies et al., 2008). Penanganan bakteri patogen di dalam bidang kesehatan serta pemanfaatan senyawa antibiotik yang ramah lingkungan yang dihasilkan oleh bakteri yang berasosiasi dengan Moluska telah menjadi pekerjaan rumah yang harus segera ditangani secara multidisiplin (Hunt and Vincent, 2006). Secara geografis perairan Ternate merupakan pertemuan lempeng antara benua Australia dan Indonesia dan biota di tempat tersebut memiliki potensi yang sangat spesifik terutama sebagai bahan farmasi bahari, maka sampel bakteri yang berasosiasi dengan Moluska dikoleksi dari perairan Ternate. Tujuan penelitian adalah mengetahui karakteristik senyawa bioaktif bakteri yang berasosiasi dengan Moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo dengan metode GCMS. II. METODE PENELITIAN Sampling Moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo dikoleksi dari perairan pulau Bastiong Kepulauan Ternate, Maluku. Selanjutnya dilakukan isolasi bakteri, skrining bakteri penghasil senyawa antiMDR, uji antibakteri moluska, isolasi bakteri patogen klinik (MDR) jenis Klebsiella, E. coli, Coagulase Negatif Staphylococcus (CNS), Enterobacter 5, Enterobacter 10 dan Pseudomonas, uji sensitifitas antibakteri, uji kepekaan terhadap kuman antimikroba. Hasil seleksi dari 12 isolat bakteri moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo yang sudah dilakukan sebelumnya memperlihatkan bahwa ada 3 isolat yang dianggap paling berpotensi untuk dijadikan sebagai sumber antibiotik baru berdasarkan konsistensi hasil skrining, besar kecilnya zona hambat yang dihasilkan dan sifat penghambatannya terhadap beberapa jenis bakteri uji yakni isolat bakteri TCM, TCA dan TOV digunakan untuk studi selanjutnya. Selanjutnya dilakukan ekstraksi DNA, amplifikasi DNA dengan metode PCR, Sekuensing DNA. Hasil sekuen 16S rDNA selanjutnya dianalisis dan diedit dengan menggunakan program GENETYX dan Analisis sekuen 16S rDNA. Amplifikasi DNA dari ke 3 isolat bakteri moluska yakni: TCM, TCA dan TOV yang sudah dilakukan sebelumnya Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 35 Karakteristik Senyawa Bioaktif Bakteri Simbion… menunjukkan bahwa semua isolat menghasilkan single band (pita tunggal) dengan ukuran sekitar 1500 bp sesuai dengan pembandingan menggunakan marker DNA. Analisis homologi menggunakan BLAST searching menunjukkan bahwa isolate TOV12.16 memiliki prosentase kesamaan tertinggi dengan genus Vibrio alginolyticus strain VM341 (96%). Sedangkan isolat TCM memiliki prosentase kesamaan tertinggi dengan genus Pseudoalteromonas sp. (99%), dan isolate TCA memiliki tertinggi dengan Vibrio sp. AC1 (99 %). Hasil identifikasi bakteri diketahui bahwa isolat TCM memiliki kekerabatan terdekat dengan Pseudoalteromonas sedangkan isolat TCA dan TOV memiliki kekerabatan yang dekat isolat yang sama-sama berada dalam genus Vibrio. Hasil penelitian ini dilajutkan untuk mengetahui karakteristik senyawa bioaktif Moluska sampel isolat TCM, TCA dan TOV dengan analisis Gas Chromatography-Mass Spectrometer (GC-MS). 2.1. Analisis Gas ChromatographyMass Spectrometer (GC-MS) Analisis GC-MS dilakukan menggunakan GCMS-QP2010S Shimadzu dengan kondisi analisis sebagai berikut : kolom Rtx-5MS 30 meter, diameter 0,25 mm, suhu terprogram dari 80 oC sampai 300 oC dengan kenaikan suhu 10 o C/menit, dan gas pembawa Helium, sedangkan untuk tekanannya sebesar 22 kPa. Jumlah senyawa yang terdapat dalam ekstrak ditunjukkan oleh jumlah puncak (peak) pada kromatogram, sedangkan nama/jenis senyawa yang ada diinterpretasikan berdasarkan data spektra dari setiap puncak tersebut dengan menggunakan metode pendekatan pustaka pada database GC/MS (Hendayana, 1994). 36 III. HASIL DAN PEMBAHASAN Skrining terhadap isolat bakteri simbion pada moluska jenis Conus miles, Stramonita armigera, Cymbiola vespertilo dari perairan pulau Bastiong Kepulauan Ternate, Maluku telah dilakukan. Hasil penelitian dapat disimpulkan bahwa isolat bakteri aktif yang diperoleh dalam penelitian ini menunjukkan hasil yang sangat menjanjikan karena ketiga isolat mampu menghambat bakteri MDR lebih dari satu jenis yang meliputi Klebsiella, Pseudomonas, Staphylococcus, E.coli dan Enterobacter. Hasil penelitian disimpulkan bahwa isolat TCM memiliki kekerabatan terdekat dengan Pseudoalteromonas sedangkan isolat TCA dan TOV memiliki kekerabatan yang dekat isolat yang sama-sama berada dalam genus Vibrio. Dari 3 isolat yang paling aktif dan menjanjikan yang berasosiasi dengan moluska dalam penelitian sebelumnya menunjukkan bahwa isolat-isolat tersebut tersebut termasuk genus Vibrio dan Pseudoaltermonas. Anggota dari Alteromonadales dan Vibrionales dalam Proteobacteria dikenal sebagai produser dominan antibiotic (Long and Azam, 2001; Grossat et al., 2004). Lebih lanjut Radjasa et al. (2007a), melaporkan aktivitas antibakteri dari bakteri Pseudoalteromonas luteoviolacea TAB 4.2 yang berasosiasi dengan karang keras Acropora sp. aktif menghambat pertumbuhan bakteri karang dan pathogen. Bakteri Pseudoalteromonas flavipulchra BSP5.1 yang merupakan simbion sponge Haliclona sp. yang diperoleh dari perairan Bandengan, Jepara mempunyai aktivitas antibakteri terhadap bakteri pathogen Alteromonas hydrophila dan Vibrio parahaemolyticus (Radjasa et al., 2007c). Penelitian lain (Radjasa et al., 2007d), melaporkan aktivitas antibakteri dari 3 isolat bakteri http://www.itk.fpik.ipb.ac.id/ej_itkt22 Pringgenies dari sponge Aaptos sp. termasuk Pseudoalteromonas luteoviolacea SPA21 yang mampu menghambat pertumbuhan bakteri MDR Escherichia coli dan Proteus sp. Bakteri genus Pseudoalteromonas diketahui memiliki fragmen gen Non-ribosomal peptide syntehase (NRPS) yang diketahui menghasilkan siderophore Alterobactin (Deng et al., 1995). Genus Vibrio diketahui sebagai sumber potensial antibiotika, Radjasa et al (2007c) menyatakan bahwa bakteri Vibrio BSP1.12 yang diisolasi dari sponge Haliclona sp. mampu menghambat pertumbuhan bakteri pathogen Alteromonas hydrophila yang merupakan causative agent pada penyakit Motile Aeromonas Septicemia (MAS) yang menyerang ikan mas. Bakteri Vibrio MJ.11 yang diisolasi dari karang lunak Porites lutea juga diketahui mampu menghambat bakteri pathogen Bacillus subtilis dan Staphylococcus sp. (Radjasa, tidak dipublikasikan). Hasil skrining menunjukkan bahwa bakteri simbion moluska sangat potensial dalam menghambat pertumbuhan bakteri MDR. Hal ini menjadi poin penting karena selama ini masalah suplai bahan baku menjadi kendala dalam pengembangan senyawa bioaktif dari avertebrata laut (Proksch et al., 2002), karena senyawa yang dihasilkan sangat terbatas sehingga dapat mengancam keberadaan avertebrata laut itu sendiri. Hasil skrining terhadap isolat bakteri simbion pada moluska yang diambil dari wilayah Maluku Utara menunjukkan bahwa bakteri simbion moluska sangat potensial dalam menghambat pertumbuhan bakteri MDR. Hal ini menjadi poin penting karena selama ini masalah suplai bahan baku menjadi kendala dalam pengembangan senyawa bioaktif dari invertebrate laut (Proksch et al., 2002), karena senyawa yang dihasilkan sangat terbatas sehingga dapat mengancam keberadaan avertebrata laut itu sendiri. Jadi dalam konteks pemanfaatan sumber daya laut yang berkelanjutan hasil penelitian ini sangat menjanjikan untuk ditindak lanjuti lebih lanjut. Analisis GC-MS dilakukan pada isolat aktif TCM6.1, TCA8.7 dan TOV12.sebagai fraksi yang memiliki aktivitas anti bakteri terbaik. Pendugaan senyawa dengan menggunakan Gas Chromatography menunjukkan terdapat beberapa senyawa yang terdeteksi dari fraksi TCM, yakni: Nitrogen oxide (N2O) (CAS) Nitrous oxide; 1,3Dioxolane, 2-(6-octynyl)-(CAS); Acetic acid (CAS) Ethylic acid; Propanoic acid,2-methyl-(CAS) Isobutyric acid; Iso-VALERIC ACID; Butanoic acid, 2methyl-(CAS) 2-Methylbutanoic acid; 1Pentadecanol (CAS) Pentadecanol; 1,2Benzenedicarboxylic acid, dioctyl ester (CAS) Dioctyl phthalate; Tetradecane,1iodo-; Cholastane, 3-thiocyanato, (3.aplpha,.5.alpha.)-(CAS) Thiocyanic acid,5.alp. Sedang fraksi TOV terdekteki senyawa Propanoic acid,2-methyl(CAS)Isobutyric acid; Butanoic acid, 2methyl-(CAS) 2-Methylbutanoid acid dan fraksi TSA terdekteksi senyawa: 1,2Propadiene (CAS) Allene seperti yang tertera pada Gambar 1, 2 dan 3. Hasil GC-MS pada sampel fraksi TCM memperlihatkan bahwa pada fraksi terdeteksi 10 puncak senyawa, tapi hanya 3 senyawa yang lebih dominan, yakni senyawa Nitrogen oxide (N2O) (CAS) Nitrous oxide; Acetic acid (CAS) Ethylic acid; Propanoic acid,2-methyl-(CAS) Isobutyric acid dan fraksi isolat TOV terdeteksi senyawa Propanoic acid,2-methyl-(CAS)Isobutyric acid; Butanoic acid, 2-methyl-(CAS) 2Methylbutanoid acid sedang fraksi isolat TSA terdeteksi senyawa 1,2-Propadiene (CAS) Allene. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 37 Karakteristik Senyawa Bioaktif Bakteri Simbion… Gambar 1. Kromatogram GC-MS TCM memperlihatkan bahwa senyawa yang dominan adalah Acetic acid (CAS) Ethylic acid Gambar 2. Kromatogram GC-MS TOV memperlihatkan bahwa Propanoic acid,2methyl-(CAS) Isobutyric acid 38 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Pringgenies Gambar 3. Kromatogram GC-MS TSA memperlihatkan bahwa Propadiene (CAS) Allene Senyawa acid memperlihatkan senyawa yang yang potensi sebagai anti bakteri dengan cara menghancurkan dinding sel dan menghambatan sintesis dinding sel (Mutchler, 1991). Diduga senyawa acid tersebutlah yang berperan memiliki kemampuan antibakteri (Kim et al., 2004). Seperti yang dinyatakan oleh Hillenga et al. (1995), bahwa Asam Benzenasetat digunakan dalam pembuatan Penicilin G. Asam Benzen-asetat mempunyai kemampuan untuk menembus membran plasma pada sel. Dapat diasumsikan bahwa mekanisme kerja dari TCM, TOV dan TSA dari bakteri simbion Moluska adalah melalui penghambatan sintesis dinding sel seperti mekanisme kerja penisilin melalui penghambatan sintesis dinding sel (Jawetz et al, 2001). Ditambahkan pula oleh Katzung (2004), bahwa antibiotik beta-laktam melintasi membran luar dan memasuki organismeorganisme gram negatif melalui saluran protein membran luar. Jadi dalam konteks pemanfaatan sumber daya laut yang berkelanjutan hasil penelitian ini sangat menjanjikan untuk ditindak lanjuti lebih lanjut. IV. KESIMPULAN Isolat bakteri aktif yang diperoleh dalam penelitian ini menunjukkan hasil yang sangat menjanjikan karena ketiga isolat mampu menghambat bakteri MDR. Hasil GC-MS pada sampel fraksi TCM memperlihatkan bahwa pada fraksi terdeteksi 10 puncak senyawa, senyawa yang lebih dominan yakni senyawa Acetic acid (CAS) Ethylic acid dan fraksi isolat TOV terdeteksi senyawa yang lebih dominan yakni Propanoic acid,2-methyl(CAS) sedang fraksi isolat TSA terdeteksi senyawa 1,2-Propadiene (CAS) Allene. Isolat fraksi TCM berpotensi memberikan kontribusi sebagai sumber alternatif baru metabolit sekunder dari bahan farmasi bahari dalam menghasilkan produk sebagai desinfektan. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 39 Karakteristik Senyawa Bioaktif Bakteri Simbion… UCAPAN TERIMA KASIH Penelitian ini dibiayai oleh Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan Nasional, Hibah Kompetensi 2010. Ucapan terima kasih disampaikan kepada Dr. Ocky K Radjasa dan Dr. Adus Sabdono MSc Jur. Ilmu Kelautan FPIK UNDIP Semarang. DAFTAR PUSTAKA Burgess J.G., K.G. Boyd, E. Amstrong, Z. Jiang, L. Yan, M. Berggren, U. May, T. Pisacane, A. Granmo, and D.R. Adams, 2003. Development of a marine natural product-based antifouling paint. Biofouling, 19:197- 205 Hunt, B., and A.C.J. Vincent. 2006. Scale and sustainability of marine bioprospecting for pharmaceuticals. Ambio, 35(2):57-64. Kim TK, MJ Garson., and J A Fuerst. 2005. Marine actinomycetes related to the Salinospora group from the Great Barrier Reef sponge Pseudoceratina clavata. Environ. Microbiol., 7:509-518. Long R, and F. Azam, 2001. Antagonistic interactions among marine pelagic bacteria. Appl Environ Microb., 67:4975-4983 Pringgenies. D, O.K. Radjasa, dan A. Sabdono. 2008. Bioprospeksi Moluska dan Bakteri Simbionnya Dalam Rangka Penanganan Strain MDR (Multi Drug Resistant. Program Intensif Riset Dasar 2007/2008.. LembagaPenelitian Universitas Diponegoro. November 2008. Laporan Penelitian. 40 Hal. Proksch, P., R.A. Edrada, and R. Ebel. 2002. Drugs from the sea-current status and microbiological implications. Appl. Microbiol. Biotechnol., 59:125-134. 40 Sukarmi dan O.K. Radjasa. 2007. Bioethical Consideration in the Search for Bioactive Compounds from Reef’s Invertebrates. J. Appl. Sci., 7(8):1235-1238 Rajasa, O.K. 2003. Marine invertebrataassociated bacteria in coral reef ecosystems as a new source of bioactive compounds. J. Coast. Dev., 7: 65-70 Radjasa, O.K. and A.Sabdono. 2003. Screening of secondary metabolite-producing bacteria associated with corals using 16S rDNA-based approach. J. Coast. Dev., 7:11-19. Radjasa, O.K., A. Sabdono, Junaidi, and E. Zocchi. 2007c. Richness of secondary metabolite- producing marine bacteria associated with sponge Haliclona sp. Int. J. Pharmacol., 3(3):275-279. Radjasa, OK, H Urakawa, K KitaTsukamoto, and K Ohwada. 2001. Characterization of psychrotro-phic bacteria in the surface and deep-sea waters from north-western Pacific Ocean based on 16S ribosomal DNA approach. Mar. Biotechnol., 3:454-462. Hunt, B., and A.C.J. Vincent. 2006. Scale and sustainability of marine bioprospecting for pharmaceuticals. Ambio, 35(2):57-64. Olivera, B.M. 2000. Conotoxin MVIIA: From marine snail venom to analgesic drug. In: Fusetani (ed.) Drugs from the sea. Karger, 74– 85pp. Watermann, B. 1999. Alternative antifoulant techniques present and future. Limno. Mar., 1(6):6-7. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 41-52, Desember 2010 AKUMULASI LOGAM BERAT PB, CU, DAN ZN DI HUTAN MANGROVE MUARA ANGKE, JAKARTA UTARA ACCUMULATION OF HEAVY METALS PB, CU, AND ZN IN THE MANGROVE FOREST OF MUARA ANGKE, NORTH JAKARTA Faisal Hamzah dan Agus Setiawan Balai Riset dan Observasi Kelautan, Kementerian Kelautan dan Perikanan [email protected], [email protected] ABSTRACT In this study, the concentrations of three kinds of heavy metals, namely Pb, Zn, and Cu from 3 species of mangrove that grow in Muara Angke were measured and analyzed. Our result showed that substrate of mangrove ecosystem in Muara Angke was dominated by clay (30.5% 62.4%), silt (21.7% -35.6%), and sand (2% -39.5%). The heavy metals accumulation in roots is higher than in sediment, water and leaves with concentration of Zn as the highest. Bioconcentration Factor (BCF; content ratio of heavy metal concentrations in roots or leaves and sediment) and Translocation Factor (TF; ratio of heavy metal concentrations in leaves and roots) of non-essential heavy metals (Pb) is higher in leaves than in roots, but for essential heavy metals (Zn and Cu), the BCF and TF was higher in roots than in leaves. TF values for heavy metals Pb, Cu, and Zn were 0.98-2.59, 0.17-0.51, and 0.52-0.86, respectively. The values of root BCF of those three heavy metals were 0.71-3.17, 0.27-0.74, and 0.95-1.53, while the values of leaf BCF were 1.84-3.45, 0.07-0.34, and 0.72-1.19, respectively. Furthermore, by calculating the phytoremediation (FTD), i.e. the difference between BCF and TF, it is obtained that Sonneratia caseolaris and Avicennia marina can be used in phytoremidiation, with leaves and roots FTD of 1.93 and 2.09, respectively for Sonneratia caseolaris and 1.93 and 1.98 for Avicennia marina. Keywords: heavy metals, mangroves, phytoremidiation, Muara Angke, bioconcentration factor, translocation factor ABSTRAK Dalam penelitian ini, konsentrasi tiga jenis logam berat, yaitu Pb, Zn, dan Cu dari 3 spesies mangrove yang tumbuh di Muara Angke diukur dan dianalisis. Hasil analisa menunjukkan bahwa substrat yang mendominasi ekosistem mangrove di Muara Angke adalah liat (30,5% 62,4%), debu (21,7%-35,6%), dan pasir (2%-39,5%). Akumulasi logam berat di akar lebih tinggi daripada di daun, dimana konsentrasi logam berat Zn adalah yang tertinggi. Nilai Bioconcentration Factor (BCF; rasio kandungan konsentrasi logam berat dalam akar atau daun dengan sedimen) dan Translocation Factor (TF; rasio konsentrasi logam berat dalam daun dan akar) logam berat non esensial (Pb) lebih tinggi di daun daripada di akar, namun untuk logam berat esensial (Zn dan Cu), BCF dan TF lebih tinggi di akar daripada di daun. Nilai TF untuk logam berat Pb, Cu, dan Zn berturut-turut adalah 0,98-2,59; 0,17-0,51 dan 0,52-0,86. Sementara itu, nilai BCF akar ketiga logam berat tersebut adalah 0,71-3,17; 0,27-0,74, dan 0,95-1,53. BCF daun logam Pb, Cu, dan Zn adalah 1,84-3,45, 0,07-0,34, dan 0,72-1,19. Selanjutnya, dengan menghitung fitoremediasi (FTD), yang merupakan selisih antara BCF dan TF, diperoleh bahwa Sonneratia caseolaris dan Avicennia marina merupakan spesies mangrove yang dapat digunakan dalam fitoremidiasi di Muara Angke, dengan FTD daun dan akar sebesar 1,93 dan 2,09 untuk Sonneratia caseolaris dan 1,93 dan 1,98 untuk Avicennia marina. Kata Kunci: logam berat, mangrove, fitoremidiasi, Muara Angke, bioconcentration faktor, translocation faktor ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 41 Akumulasi Logam Berat Pb, Cu, dan Zn… I. PENDAHULUAN Mangove merupakan salah satu ekosistem pesisir yang mempunyai peranan penting di daerah estuari. Ekosistem mangrove memiliki tingkat produktivitas paling tinggi dibandingkan dengan ekosistem pesisir lainnya. Mangrove juga merupakan tempat mencari makan, memijah dan berkembang biak bagi udang dan ikan serta kerang dan kepiting. Ekosistem mangrove bagi manusia juga bermanfaat baik secara langsung dan tidak langsung terhadap sosio-ekonomi penduduk sekitar. Selain itu, ekosistem mangrove juga berfungsi sebagai perangkap sedimen dan mencegah erosi serta penstabil bentuk daratan di daerah estuari (Harty, 1997). Disamping kegunaan mangrove yang begitu banyak, adapula usaha dan aktvitas lain yang menyebabkan luasan mangrove berkurang. Kegiatan ini seperti reklamasi pantai, pembukaan lahan untuk pertanian dan perikanan budidaya, industri serta pengembangan perumahan didaerah pesisir (Eong, 1995). Dampak dari aktivitas diatas dapat menyebabkan secara langsung masuknya limbah kedalam ekosistem estuari yang salah satunya adalah logam berat (MacFarlane, 2002). Peningkatan kadar logam berat pada ekosistem mangrove dapat juga berasal dari perkapalan, wisata, tumpahan minyak, pengolahan limbah tumbuhan serta peningkatan sampah dan aktivitas pertambangan (Peters et al., 1997). Masukan dari aktivitas pertanian seperti penggunaaan insektisida dan pupuk yang berlebihan juga meningkatkan konsentrasi logam berat di estuari (Alloway, 1994). Konsentrasi logam berat yang tinggi akan menyebabkan kerusakan lingkungan dan meningkatkan daya toksisitas, persistan dan bioakumulasi logam itu sendiri (Lindsey et al., 2004). 42 Secara umum, logam berat untuk pertumbuhan dan perkembangan tumbuhan dibagi menjadi dua yaitu logam esensial dan non esensial. Cu dan Zn merupakan logam yang termasuk esensial, sedangkan Pb merupakan logam non esensial bagi tumbuhan (Baker dan Walker, 1990 dalam MacFarlane and Burchett, 2002). Cu sangat berguna untuk pertumbuhan jaringan tumbuhan terutama jaringan daun dimana terdapat proses fotosintesis (Kamaruzzaman et al., 2008). Selain itu, Cu juga mempunyai fungsi sebagai salah satu mikronutrien yang diperlukan didalam mitokondria dan kloroplas, enzim yang berhubungan dengan transpor elektron II, proses sintesis dan metabolisme karbohidrat dan protein serta sebagai dinding sel lignin (Verkleij dan Schat, 1990 dalam MacFarlane and Burchett, 2003). Pb merupakan logam yang sangat rendah daya larutnya bersifat pasif, dan mempunyai daya translokasi yang rendah mulai dari akar sampai organ tumbuhan lainnya. Pb juga memiliki toksisitas yang tertinggi dan menyebabkan racun bagi beberapa spesies (Wozny dan Kzreslowka, 1993 dalam MacFarlane and Burchett, 2002). Zink sangat berguna dalam sistem enzim, enzim aktivator dalam proses respirasi dan hormon pertumbuhan. Mills (1995) telah melakukan observasi terhadap beberapa logam berat yang masuk ke daerah estuari, dan hasilnya adalah sebagian besar logam yang masuk kedalam estuari berasal dari aktivitasi industri antara lain Cu, Pb, dan Zn. Ketiga logam tersebut masuk kedaerah estuari dengan konsentrasi yang tinggi dengan peningkatan nilai konsentrasi sampai dengan 1000 µg Cu g-1, 1000 µg Pb g-1, dan 2000 µg Zn g-1 didalam sedimen yang terkontaminasi (Irvine dan Birch, 1998). Walaupun masukan sumber pencemar sangat banyak, mangrove memiliki toleransi yang tinggi terhadap logam http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hamzah dan Setiawan berat (Macfarlane dan Burchett, 2001). Hal ini menunjukkan bahwa mangrove secara aktif menghindari masukan logam berat yang berlebih dan berfungsi sebagai penyaring dan memiliki daya treatment khas secara alami melalui organ akar (Clark et al., 1998 dalam Kammaruzaman et al., 2008) Akumulasi logam berat terjadi pada akar dan dibawa ke jaringan lainnya dan proses ini bisa membatasi masuknya udara ke dalam jaringan tersebut (Silva et al., 1990; Chiu dan Chou, 1991 dalam MacFarlane et al., 2003). Hutan Lindung Angke Kapuk (HLAK) merupakan kawasan konservasi di daerah Muara Angke yang terletak di ujung Jakarta dengan luas hutan seluas 44,7 Ha. Luasan hutan ini termasuk kecil, namun mengingat keberadaan dan fungsinya kawasan ini mempunyai nilai yang sangat khusus. Sebagian besar ekosistem yang tumbuh didaerah HLAK yaitu mangrove jenis Avicennia marina, Rhizophora mucronata, dan Sonneratia caseolaris. Penelitian ini bertujuan untuk mengetahui akumulasi logam pada mangrove terutama pada jaringan utama seperti akar dan daun. Selain itu, diketahuinya perbandingan konsentrasi logam antara di mangrove dengan di lingkungan sehingga diketahui apakah vegetasi tersebut dapat dijadikan indikator pencemaran logam berat di kawasan Muara Angke. II. METODE PENELITIAN 2.1. Lokasi dan Jenis Sampel Survey pengambilan sampel dilaksanakan pada bulan Juni 2010 di Hutan Lindung Angke Kapuk (HLAK), Muara Angke, Jakarta. HLAK merupakan daerah konservasi hutan mangrove berada dibawah koordinasi Dinas Kelautan dan Pertanian DKI Jakarta. Sebanyak 3 titik diambil pada penelitian ini yaitu stasiun 2A, 2B dan 5 (Gambar 1). Pemilihan jenis mangrove berdasarkan spesies yang dominan yang ditemukan di daerah penelitian. Secara umum jenis mangrove yang dominan yang ditemuakan adalah Avicennia marina, Rhizophora mucronata, dan Sonneratia caseolaris. Sampel yang diambil meliputi parameter fisika kimia air dan sedimen yaitu suhu, pH, salinitas air dan tanah, kandungan oksigen terlarut (DO). Logam yang dianalisa meliputi logam esensial dan non esensial bagi mangrove yaitu Pb, Cu, dan Zn. Sampel untuk analisa logam berat berasal dari air, sedimen, akar dan daun mangrove. 2.2. Pengambilan Sampel dan Metode Ekstraksi Pengukuran insitu dilakukan terhadap parameter suhu, salinitas, kandungan oksigen terlarut (DO) dan pH. Suhu diukur dengan menggunakan termometer, salinitas air diukur dengan menggunakan refraktometer sedangkan salinitas sedimen diukur dengan SCT Meter. pH air diukur dengan menggunakan pH meter sedangkan pH sedimen diukur dengan mengunakan ekstrak H2O dan CaCl2 dimana pH yang diekstrak dengan air merupakan kemasaman aktif (aktual) dan ekstrak CaCl2 0.01 M merupakan kemasaman potensial (cadangan). Kandungan oksigen terlarut diukur dengan menggunakan metode titrasi winkler. Sampel air untuk logam diambil dengan menggunakan Van Dorn dan dimasukkan kedalam botol polietilen yang terlebih dahulu dibersihkan dengan menggunakan HCl 6 N dan dibilas dengan aquadest (Hutagalung et.al., 1997). Sampel sedimen diambil dengan menggunakan Ekman grab dan dimasukan kedalam botol polietilen. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 43 Akumulasi Logam Berat Pb, Cu, dan Zn… Gambar 1. Lokasi Penelitian di Muara Angke, Juni 2010 Kandungan logam berat pada air diukur dengan cara terlebih dahulu menghilangkan ion mayor seperti Na+, Ca2+, SO4-2, K+, dan Mg2+ dengan menambahkan metil iso butil keton, APDC, dan NaDDC sehingga memudahkan proses adsorbsi logam berat oleh AAS (Hutagalung et.al., 1997). Untuk logam berat pada sedimen juga dihilangkan ion mayor kemudian ditambahkan HF hingga suhu mencapai 1300C. Setelah dingin, sampel siap diukur dengan AAS menggunakan nyala udara asitilen. Fraksinasi sedimen juga dilakukan pada penelitian ini. Fraksinasi sedimen dilakukan dengan menganalisa tekstur dengan menggunanakan metode fraksional yang menggunakan alat saringan bertingkat. Sampel akar dan daun mangrove diambil dengan menggunakan gunting tanaman. Sampel akar yang diambil merupakan akar mangrove yang masuk kedalam sedimen, sedangkan daun mangrove yang diambil merupakan daun mangrove yang tidak terlalu tua dan tidak juga terlalu muda. Sebanyak ± 30 daun 44 diambil dari 5 jenis pohon 3 spesies. Pohon yang diambil mempunyai diamater berkisar antara 15-20 cm dengan tinggi 3-5 m. Sampel daun dan akar, terlebih dahulu di keringkan dengan menggunakan oven pada suhu 1050C selama 24 jam. Tambahkan HNO3 dan HClO4 (APHA, 2005), Kemudian dipanaskan dan ditambah HNO3, dan siap diukur dengan menggunakan nyala udara asitilen. 2.3. Analisa Data Untuk mengetahui terjadi akumulasi logam pada mangrove dilakukan dengan cara menghitung konsentrasi logam pada sedimen, akar dan daun. Perbandingan antara konsentrasi logam di akar/daun dengan konsentrasi di sedimen dikenal dengan bio-concentration factor (BCF). BCF pada daun dan akar dihitung untuk mengetahui seberapa besar konsentrasi logam pada daun dan akar yang berasal dari lingkungan (MacFarlane et al., 2007). Selain itu juga dihitung perbandingan antara konsentrasi logam http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hamzah dan Setiawan pada daun dan akar yang dikenal sebagai translocation factors (TF). Nilai TF dihitung untuk mengetahui perpindahan akumulasi logam dari akar ke tunas (MacFarlane et al., 2007). Selisih antara nilai BCF dan TF digunakan untuk menghitung fitoremediasi (Yoon et al., 2006). Pearson correlation digunakan untuk mengetahui hubungan antara konsentrasi logam pada sedimen, akar dan daun. III. HASIL DAN PEMBAHASAN 3.1. Paramater Fisika dan Kimia Parameter fisika dan kimia air yang diukur secara insitu adalah adalah pH, suhu, salinitas dan oksigen terlarut sedangkan untuk sedimen, paramater yang diukur adalah salinitas dan pH. Secara umum, pH hasil pengukuran diketiga lokasi hampir sama yaitu berkisar 7,53 - 7,68. Nilai tertinggi untuk pH ditemukan pada Stasiun 2B. Untuk suhu, secara umum juga hampir sama di semua lokasi penelitian yaitu berkisar antara 28.5-29 0C (Tabel 1). Nilai suhu dan pH berdasarkan baku mutu air laut untuk biota laut didaerah mangrove masih berada dibawah baku mutu (KepMen LH No 51, 2004). Nilai salinitas dan oksigen terlarut memiliki nilai yang beda antarstasiun. Stasiun 5 merupakan stasiun yang paling rendah salinitas dan DO-nya (3 ‰ & 1,77 mg/l), sedangkan stasiun 2B memiliki salinitas dan DO paling tinggi yaitu 19 ‰ dan 6,39 mg/l. Rendahnya salinitas pada stasiun 5 diduga bahwa Stasiun 5 dekat dengan Sungai Angke, sehingga mendapat pengaruh dari masukan air tawar. Kondisi substrat yang bersifat anaerob merupakan salah satu ciri daerah estuari. Hal ini mengindi-kasikan bahwa secara umum oksigen terlarut di daerah estuari memang sangat rendah dan bersifat anaerob. Oksigen terlarut dari air diserap ke sedimen dan digunakan untuk kegiatan respirasi oleh bakteri (Hogarth, 1999). Salinitas pada sedimen berkisar 1225 ‰, sedangkan pH sedimen yang diukur melalui pendekatan H2O berkisar 5,2-7,5 dan CaCl2 berkisar 5,1-7,4. Secara umum pH dengan menggunakan pendekatan ekstraksi H2O dan CaCl2 tidak jauh beda antarstasiun dan jika dibandingkan antara pH air dan sedimen, maka stasiun 2A dan 2B memiliki pH yang sama, tetapi untuk stasiun 5 memiliki perbedaaan nilai antara air dan sedimen. Pengukuran pH dengan menggunakan ekstraksi H2O akan mendapatkan hasil lebih besar dibandingkan dengan menggunakan ekstrak CaCl2 0.01 M karena ekstrak CaCl2 merupakan kemasaman cadangan sehingga nilainya selalu lebih kecil dari ekstrak air. Faktor lain yang mempengaruhi rendahnya nilai pH pada stasiun 5 adalah proses reaksi reduksi dan oksidasi yang terjadi pada sedimen diduga bisa mengurangi kandungan pH. Secara umum, kondisi ekosistem mangrove di Muara Angke berada didaerah tercemar (Teluk Jakarta). Masukan limbah yang berasal dari industri, pelabuhan, dan aktivitas rumah tangga menyebabkan tingginya pencemaran yang terjadi terutama logam berat. Sumber pencemar yang berasal dari aktivitas rumah tangga yang mengandung bahan organik akan mempengaruhi kondisi ekosistem mangrove terutama sedimen. Bahan organik yang ada pada ekosistem mangrove secara alami berasal dari serasah mangrove. Selain itu, bahan organik juga akan mempengaruhi derajat keasaman (Setyawan, 2008). Peningkatan logam didaerah estuari juga dipengaruhi oleh faktor sedimen yang dipengaruhi oleh pH, bahan organik, perpindahan kation, spesies mangrove, dan umur mangrove. Mobilitas logam dan ketersediaan dari logam berat di sedimen Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 45 Akumulasi Logam Berat Pb, Cu, Dan Zn… secara umum rendah. Kondisi ini ditentukan sekali oleh pH yang tinggi, sedimen berbentuk lempung dan bahan organik tinggi. (Jung dan Thomton, 1996; Rosselli et al., 2003 dalam Yoon et al., 2006). 3.2. Fraksinasi Sedimen Sedimen di daerah Muara Angke terdiri dari berbagai tipe substrat dengan ukuran yang berbeda, penentuan jenis dan komposisi sedimen dilakukan dengan mengidentifikasi fraksi-fraksi pemben- tuknya yakni pasir, debu dan liat. Gambar 2 menunjukkan bahwa, berdasarkan hasil analisa laboratorium, komposisi substrat didaerah Muara Angke sebagian besar didomininasi oleh liat (30,5 - 62,4%), sedangkan untuk fraksi lainnya yaitu debu (21,7%-35,6%), dan pasir (2%-39,5%). Stasiun 2B dan 2A merupakan stasiun tertinggi komposisi liatnya yaitu 62,4% dan 45,6%. Stasiun 5 lebih didominasi oleh pasir dengan jumlah persentase mencapai 39, 5%. Tabel 1. Parameter fisika kimia air dan sedimen di daerah Muara Angke No Parameter Satuan Stasiun 5 2A 2B 7,56 7,53 7,68 C 28,5 29 29 Air 1 pH (in-situ) o 2 Suhu (in-situ) 3 Salinitas (in-situ) ‰ 3 10 19 4 Oksigen Terlarut mg/L 1,77 5,68 6,39 ‰ 13 12 25 H2O - 5,2 7,5 7,5 CaCl2 - 5,1 7,4 7,4 Sedimen 5 Salinitas 6 pH Gambar 2. Komposisi tekstur sedimen di Muara Angke 46 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hamzah dan Setiawan Prosentase antara liat dan debu pada Stasiun 5 hampir sama yaitu 30,5% dan 30%. Kondisi substrat dengan fraksi lumpur akan berpengaruh terhadap konsentrasi logam (Hogarth, 1999). 3.3. Kandungan logam berat Pb, Cu dan Zn pada air dan sedimen Hasil pengukuran kandungan logam Cu dan Pb total pada air menunjukkan kurang dari 0,006 ppm, sedangkan untuk logam Zn total berkisar antara 0,044-0,062 ppm. Pada sedimen, kandungan logam berat Cu total berkisar antara 28,41-51,36 ppm, kandungan logam Zn total berkisar 56,58-69,3 ppm dan 18,64-29,57 ppm untuk logam Pb total. Kandungan rerata logam berat Pb, Cu, dan Zn pada air dan sedimen dapat dilihat pada Tabel 2. Secara umum kandungan logam baik Pb, Cu, dan Zn dalam air memiliki nilai yang lebih rendah dibandingkan dengan yang di sedimen. Dalam air, logam Zn memiliki konsentrasi tertinggi di dibandingkan logam lainnya. Pb dan Cu memiliki daya larut yang rendah dibandingkan dengan Zn, sehingga di perairan konsentrasinya rendah. Di sedimen, konsentrasi logam Zn memiliki nilai konsentrasi yang tinggi dibandingkan dengan Cu dan Pb. Tingginya konsentrasi logam Zn total baik di air dan sedimen dapat menunjukkan bahwa terdapat masukan dari aktivitas industri, pelabuhan, dan perumahan yang menuju HLAK. Jika dikaitkan dengan kondisi perairan yang asam, Zn akan berikatan dengan Cl- dan mudah terlarut sehingga bisa meningkat konsentrasi Zn. Tingginya kandungan Zn di sedimen dibandingkan dengan air dikarenakan ion Zn sangat mudah terserap kedalam sedimen. Dalam hal ini, konsentrasi Zn dalam air, tidak berupa ion melainkan berbentuk senyawa (Effendi, 2003). 3.4. Kandungan logam berat Pb, Cu, dan Zn pada daun dan akar Kandungan logam Cu pada akar berkisar antara 12,17-37,68 ppm dengan konsentrasi tertinggi terdapat pada species Avicennia marina ke-3 stasiun 2. Kandungan logam Zn total pada akar berkisar 55,38-99,88 ppm, sedangkan kandungan logam Pb berkisar 20,9868,78 ppm. Logam Zn tertinggi ditemukan di St. 2 pada species Avicennia marina ke-5 dan Pb teringgi ditemukan di stasiun 5 pada spesies Sonneratia caseolaris. Kandungan logam Cu pada daun berkisar 2,07 - 10,07 ppm dengan konsentrasi tertinggi terdapat pada spesies Avicennia marina stasiun 2 (Tabel 3). Tabel 2. Kandungan rerata logam berat Pb, Cu dan Zn pada air dan sedimen Stasiun No Parameter Satuan 5 2A 2B Air 1 Cu Total ppm <0,006 <0,006 <0,006 2 Pb Total ppm <0,006 <0,006 <0,006 3 Zn Total Sedimen ppm 0,062 0,048 0,044 4 Cu Total ppm 28,41 29,68 51,36 5 Zn Total ppm 56,58 65,28 69,3 6 Pb Total ppm 23,23 18,64 29,57 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 47 Akumulasi Logam Berat Pb, Cu, Dan Zn… Penyerapan tetap dilakukan, namun dalam jumlah yang terbatas dan terakumulasi di akar. Selain itu, terdapat sel endodermis pada akar yang menjadi penyaring dalam proses penyerapan logam berat. Dari akar, logam akan di translokasikan ke jaringan lainnya seperti batang dan daun serta mengalami proses kompleksasi dengan zat yang lain seperti fitokelatin. (Baker dan Walker, 1990 dalam MacFarlane et al., 2003). Logam Zn dan Pb berkisar 47,8667,45 ppm dan 54,31-85,48 ppm, dengan konsentrasi logam Zn dan Pb tertinggi terdapat pada stasiun 5, spesies Sonneratia caseolaris dan Rhizophora mucronata. Kandungan logam berat pada akar dan daun dapat dilihat pada Tabel 4. Jika dilihat perbandingan konsentrasi logam pada akar dan daun, maka kandungan logam berat pada akar lebih tinggi dibandingkan pada daun. Logam Cu dan Zn lebih tinggi pada akar dibandingkan pada daun, namun sebaliknya konsentrasi logam Pb lebih tinggi di daun dibandingkan pada akar. Tingginya konsentrasi logam Pb pada daun diduga tingkat mobilitas logam Pb yang tinggi, namun berbeda dengan apa yang dilakukan oleh MacFarlane et al., (2003). Kandungan logam berat Pb pada Avicennia marina pada kondisi terkontrol lebih tinggi di akar dibandingkan di daun. Baker (1981) dalam MacFarlane et al., (2003) menyatakan bahwa Avicennia marina merupakan spesies mangrove yang sangat ketat dalam menyerap logam Pb bahkan sampai tidak menyerap sama sekali. Berdasarkan mekanisme fisiologis, mangrove secara aktif mengurangi penyerapan logam berat ketika konsentrasi logam berat di sedimen tinggi. 3.5. Akumulasi dan translokasi logam Cu, Zn, dan Pb Berdasarkan data konsentrasi akar, daun dan sedimen, akumulasi logam bisa dilihat dengan cara membandingkan konsentrasi antar jaringan tumbuhan mangrove. Baker dan Brooks (1989) menyatakan bahwa, tumbuhan mampu mengakumulasi logam berat hingga > 1000 mg kg-1 dan dikenal sebagai hiperakumulator. Pada dasarnya, tumbuhan mempunyai daya toleransi dan mengakumulasi logam berat dan hal ini berkaitan dengan tujuan fitostabilisasi. Bioconcentration factors (BCF) dan Translocation factors (TF) bisa digunakan untuk menduga tumbuhan yang bisa dijadikan sebagai fitoremidiasi. Tabel 3. Kandungan rerata logam berat Pb, Cu dan Zn pada daun dan akar Lokasi dan Spesies mangrove No Parameter Satuan 5 - RM 3 5 - SC 4 2A - AM 3 2A - AM 4 2A - AM 5 Akar 1 Cu Total ppm 12,17 15,36 37,68 13,88 22,04 2 Zn Total ppm 55,38 83,05 95,61 66,18 99,88 3 Pb Total Daun ppm 53,89 68,78 20,98 57,52 59,16 4 5 Cu Total Zn Total ppm ppm 2,07 47,86 2,57 67,45 8,45 50,15 7,08 51,76 10,07 54,22 6 Pb Total ppm 85,48 67,71 54,31 61,93 64,32 Keterangan : 5 - RM 3 : Rhizophora mucronata St.5; 5 - SC 4: Sonneratia caseolaris St. 5; 2A - AM 3: Avicennia marina St. 2; 2A - AM 4: Avicennia marina St. 2A; 2A - AM 5: Avicennia marina St. 2A 48 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hamzah dan Setiawan Tabel 4. Akumulasi dan translokasi logam Cu, Zn dan Pb di daerah Muara Angke Spesies Stasiun Rhizophora mucronata Sonneratia caseolaris Avicennia marina-3 Avicennia marina-4 Avicennia marina-5 5 5 2B 2B 2A BCF Daun BCF Akar TF Cu Zn Pb Cu Zn Pb Cu Zn Pb 0,07 0,09 0,16 0,14 0,34 0,85 1,19 0,72 0,75 0,83 3,68 2,91 1,84 2,09 3,45 0,43 0,54 0,73 0,27 0,74 0,98 1,47 1,38 0,95 1,53 2,32 2,96 0,71 1,95 3,17 0,17 0,17 0,22 0,51 0,46 0,86 0,81 0,52 0,78 0,54 1,59 0,98 2,59 1,08 1,09 Berdasarkan Tabel 4, dapat dilihat bahwa dari 5 pohon mangrove untuk logam Pb, Rhizophora mucronata pada stasiun 5 memiliki nilai BCF daun paling tinggi yaitu 3,68 kemudian diikuti oleh Avicennia marina-5 pada stasiun 2A (3,45). Hal ini bisa dilihat bahwa konsentrasi logam Pb total pada daun memang memiliki nilai tertinggi dibandingkan dengan daun mangrove lainnya dan juga didukung oleh konsentrasi total logam Pb pada sedimen di stasiun 5 lebih tinggi dibandingkan pada stasiun 2A (23,23 ppm). Berbeda dengan logam Pb, nilai BCF daun untuk logam Zn tertinggi ditemukan pada spesies Sonneratia caseolaris (1,19) di stasiun 5 dan untuk BCF daun logam Cu tertinggi ditemukan di stasiun 2A spesies Avicennia marina-5 (0,34). Nilai BCF daun rata-rata logam Pb dan Zn (2,46 dan 0,76) spesies Avicennia marina lebih besar dibandingkan oleh yang dilakukan McFarlane et al., 2007 (BCF= 0,17 dan 0,45). Akumulasi logam dari sedimen menuju akar yang dilihat dari nilai BCF akar. Secara umum, akumulasi dan penyerapan logam Pb, Cu, dan Zn dari sedimen ke akar tertinggi ditemukan pada spesies Avicennia marina-5 di stasiun 2A dengan nilai BCF akar logam Pb, Cu, dan Zn berturut-turut adalah 3,17; 0,74 dan 1,53. Tingginya nilai BCF akar untuk semua logam didukung oleh tingginya konsentrasi semua logam pada akar dan rendah pada sedimen sehingga menghasilkan nilai BCF akar yang tinggi. Nilai BCF daun rata-rata logam Cu (0,58) spesies Avicennia marina lebih kecil dibandingkan oleh yang dilakukan McFarlane et al., (2007) (BCF= 0,92), sedangkan untuk BCF rata-rata akar logam Pb dan Zn jauh lebih tinggi (1,94 dan 1,62) dibandingkan dengan yang dilakukan oleh McFarlane et al., (2007) (BCF akar Pb=0,57; BCF akar Zn=0,69). Translokasi logam dihitung antara rasio konsentrasi logam di daun dan di akar. Berdasarkan hasil perhitungan, didapatkan bahwa nilai TF Cu, Zn, dan Pb berkisar 0,17-2,59. Nilai TF tertinggi ditemukan pada spesies Avicennia marina-3 stasiun 2B untuk logam Pb dan nilai TF terendah ditemukan pada logam Cu spesies Rhizophora mucronata dan Sonneratia caseolaris (0,17). Nilai TF daun rata-rata logam Pb dan Zn (1,58 dan 0,61) spesies Avicennia marina lebih besar dibandingkan oleh yang dilakukan McFarlane et al., 2007 (TF= 0,41 dan 0,51) namun, untuk logam Cu pada spesies yang sama, nilai TF rata-rata lebih kecil (TF= 0,39) dibandingkan yang dilakukan McFarlane et al. (2007) (TF=0,50). Jika dilihat kandungan total logam Pb pada akar dan daun Avicennia marina-3 pada Tabel 4, spesies ini merupakan spesies yang paling kecil memiliki kandungan logam dibandingkan dengan Rhizophora mucronata dan Sonneratia caseolaris. Avicennia marina merupakan salah satu jenis mangrove yang mensekresi garam melalui daun. Selain itu juga pada bagian akar terdapat pneumatopora sehingga dapat mentrans- Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 49 Akumulasi Logam Berat Pb, Cu, Dan Zn… lokasikan berbagai kandungan garam dan logam menuju daun (MacFarlane et al., 2007). Translokasi logam dari akar ke daun untuk logam esensial (Cu dan Zn) sangat rendah dibandingkan pada logam non esensial (Pb). Rendahnya nilai TF pada logam esensial menunjukkan bahwa mangrove menggunakan kedua logam tersebut untuk aktivitas metabolisme dan pertumbuhan. Sedangkan untuk logam non esensial, proses mobilitas logam dari akar ke daun sangat tinggi, hanya Sonneratia caseolaris yang nilai mempunyai nilai TF dibawah 1 (0,98). Pada daun, Pb bersifat racun terutama pada saat tumbuhan melakukan fotosintesis, sintesa khlorofil, dan sintesa enzim antioksidan (Kim et al., 2003 dalam Yoon et al., 2006). Terkadang akar juga mempunyai sistem penghentian transpor logam menuju daun terutama logam non esensial, sehingga ada penumpukkan logam di akar (Yoon et al., 2006). Analisa Pearson Correlation dilakukan untuk mengetahui hubungan antara kandungan logam pada daun dan sedimen, serta kandungan logam pada sedimen dan akar. Berdasarkan perhitungan didapatkan bahwa hubungan antara kandungan logam pada sedimen dan daun berkorelasi negatif terjadi pada logam Cu dengan nilai korelasi -0.296 dan berkorelasi positif pada logam Zn dan Pb dengan nilai korelasi masingmasing 0.192 dan 0.135. Hal ini menunjukkan bahwa, konsentrasi Cu pada daun secara tidak langsung ditranslokasikan dari sedimen menuju daun, tetapi diakumulasikan terlebih dahulu diakar sedangkan pada logam Pb dan Zn, sekitar 19 % dan 13 % kedua logam tersebut di transfer dari sedimen menuju daun. Hubungan antara kandungan logam berat di akar dan di sedimen untuk logam Pb, Cu dan Zn berturut-turut adalah -0.57, 0.57, dan 0.04. Korelasi antara kandungan logam Cu pada akar dan sedimen mempunyai hubungan yang positif yaitu sebesar 57%. Hal ini mengindikasikan bahwa sebanyak 57 % logam di sedimen diserap oleh organ akar. Sedangkan untuk logam Pb dan Zn mempunyai hubungan negatif. Ma et al. (2001) menyatakan bahwa fitoremidiasi merupakan salah satu solusi yang murah biaya, waktu yang lama, dan hemat tenaga didaerah terkontaminasi. Salah satu aplikasi dari fitoremidiasi yaitu fitostabilisasi. Fitostabilasasi merupakan usaha untuk mengurangi kandungan polutan dimana tumbuhan yang digunakan sebagai sarananya dengan tujuan mengurangi tingkat pergerakan logam pada tanah atau sedimen. Fitoremidiasi (FTD) merupakan selisih antara nilai BCF dan TF. FTD akan maksimal jika BCF tinggi dan TF rendah (Yoon et al., 2006). Berdasarkan Tabel 6, Avicennia marina-5 stasiun 2A merupakan jenis mangrove yang mempunyai nilai FTD paling besar dibandingkan dengan jenis mangrove lainnya (FTD akar dan daun; 1,93 dan 2,09), kemudian diikuti oleh Tabel 5. Fitoremidiasi pada akar dan daun di Muara Angke 50 Spesies Stasiun Rhizophora mucronata Sonneratia caseolaris Avicennia marina-3 Avicennia marina-4 Avicennia marina-5 5 5 2B 2B 2A FTD Daun Cu Zn -0,1 -0,02 -0,08 0,38 -0,06 0,2 -0,37 -0,04 -0,12 0,29 Pb 2,09 1,93 -0,8 1,02 2,36 Cu 0,26 0,37 0,51 -0,24 0,29 http://www.itk.fpik.ipb.ac.id/ej_itkt22 FTD Akar Zn Pb 0,11 0,73 0,66 1,98 0,86 -1,88 0,17 0,87 0,99 2,09 Hamzah dan Setiawan Sonneratia caseolaris stasiun 5 dengan nilai FTD daun dan akar masing-masing 1,93 dan 1,98. Untuk spesies Rhizophora mucronata stasiun 5, spesies ini mempunyai nilai FTD daun cukup tinggi (2,09), namun nilai FTD akar cukup kecil (0,73) dan juga nilai TF cukup besar (1,59). Dari uraian diatas dapat disimpulkan bahwa untuk didaerah Muara Angke, spesies mangrove yang dapat digunakan untuk tujuan fitoremidiasi dilingkungan tercemara adalah Avicennia marina-5 dan Sonneratia caseolaris. Fitoremidiasi dan fitostabilisasi bisa digunakan untuk mengurangi pergerakan polutan didalam tanah/sedimen. Proses ini menggunakan kemampuan akar tanaman (mangrove) untuk mengubah kondisi lingkungan tercemar berat menjadi sedang bahkan ringan. Mangrove bisa mengentikan atau mengurangi proses penyerapan dan akumulasi logam berat melalui akar. Proses ini akan mengurangi pergerakan logam dan mengencerkannya serta mengurangi logam masuk kedalam system rantai makanan di daerah estuari (Susarla et al., 2002). IV. KESIMPULAN Secara umum konsentrasi logam berat di Muara Angke pada akar mangrove lebih tinggi dibandingkan pada daun, sedimen dan air dengan konsentrasi: akar (12,17-99,88 ppm), daun (2,07-85,48 ppm), sedimen (18,6469,3 ppm), air (<0,006-0,0062 ppm). Konsentrasi Zn paling tinggi dibandingkan dengan Pb dan Cu. Akumulasi logam berat pada daun dan akar bisa dihitung melalui BCF. Nilai BCF daun tertinggi pada logam Pb species Avicennia marina (3,45) dan BCF akar juga tertinggi pada logam Pb pada Avicennia marina (3,17). Nilai TF tertinggi ditemukan pada spesies Rhizophora mucronata (1,59). Untuk tujuan fitoremidiasi, Sonneratia caseolaris dan Avicennia marina merupakan spesies mangrove yang dapat digunakan didaerah Muara Angke. DAFTAR PUSTAKA Alloway, B.J. 1994. Toxic metals in soil–plant systems. Chichester, UK: John Wiley and Sons. American Public Health Association, American Water Works Assosiation dan Water Pollution Control Federation. 2005. Standard Methods for the Examination of Water and Wastewater, AWWA, WPCF. 21st Eds. Hal 3-10. Baker, A.J.M. and R.R. Brooks. 1989. Terrestrial higher plants which hyperaccumulate metallic elements—a review of their distribution, ecology and phytochemistry. Biorecovery,1:81–126. Eong, O.J. 1995. The ecology of mangrove conservation and management. Hydrobiologia, 295: 343–351. Harty, C. 1997. Mangroves in New South Wales and Victoria. Vista Publications, Melbourne, 47 pp. Hutagalung, H.P, D. Setiapermana, dan S.H. Riyono. 1997. Metode Analisis Air laut, Sedimen dan Biota (Buku Kedua). P3O LIPI. Jakarta. Irvine, I. and G. F. Birch. 1998. Distribution of heavy metals in surficial sediments of Port Jackson, Sydney, New South Wales. Aust. J. Earth Sci., 45:297–304. Kamaruzzaman, B.Y., M.C. Ong., K.C.A., Jalal., S. Shahbudin., dan O.M. Nor. 2008. Accumulation of Lead and Copper in Rhizophora apiculata from Setiu Mangrove Forest, Terengganu, Malaysia. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 51 Akumulasi Logam Berat Pb, Cu, Dan Zn… Journal of Environ-mental Biology: 821-824 KepMen LH Nomor 51 Tahun 2004. Baku mutu Air Laut. Lindsey, H.D., M.M. James, and M.G. Hector. 2004. An Assessment of Metal Contamination in Mangrove Sediments and Leaves from Punta Mala Bay, Pacific Panama. Marine Pollution Bulletin., 50:, 547-552. Ma, L. Q., K.M. Komar, C. Tu, and W. A. Zhang. 2001. A fern that Hyperaccumulator arsenic. Nature 409:579. MacFarlane, G.R. and M.D. Burchett. 2001. Photosynthetic pigments and Peroxides activity as indicators of Heavy Metal stress in the Grey Mangrove Avicennia marina (Forsk.) Veirh. Marine Pollution Bulletin, 42: 233-240. MacFarlane, G.R. 2002. Leaf biochemical parameters in Avicennia marina (Forsk.) Vierh as potential biomarkers of heavy metal stress in estuarine ecosystems. Mar. Pollut. Bull, 44: 244–256. MacFarlane, G.R. and M.D. Burchett. 2002. Toxicity, growth and accumulation relationships of copper, lead and zinc in the Grey Mangrove Avicennia marina (Forsk.) Veirh. Marine Environmental Research, 54: 65–84. MacFarlane, G.R., Pulkownik, and M.D., Burchett. 2003. Accumulation and Distribution of Heavy Metals in grey Mangrove, Avicennia marina (Forsk) Vierh: Biological indication potential. Environmental Pollution, 123: 139-151. MacFarlane, G.R., E.C. Koller, and S.P. Blomberg. 2007. Accumula-tion and Patitioning of Heavy Metals in Mangrove: A Synthesis of Fieldbased Studies. Chemosphere. 14541464. 52 Mills, W.B. 1995. Water Quality Assessment: A Screening Procedure for Toxic and Conventional Pollutants in Surface and Ground Water – Part 1. US EPA, Georgia. Peters, E.C., N.J. Gassman, J.C. Firman, R.H. Richmond, and E.A. Power. 1997. Ecotoxicology of tropical marine ecosystems. Environmental Toxicology and Chemistry, 16:12– 40. Rao, N.S. 1994. Mikroorganisme Tanah dan Pertumbuhan Tanaman. Edisi kedua. Jakarta: Penerbit Universitas Indonesia. Setyawan, A.D. 2008. Biodiversitas Ekosistem Mangrove di Jawa; Tinjauan Pesisir Utara dan Selatan Jawa Tengah. Cetakan Pertama. Pusat Penelitian dan Pengembangan Bioteknologi dan Biodiversitas, LPPM Universitas Sebelas Maret. Surakarta: 71-79. Susarla S., V.F. Medina, and S.C. McCutcheon .2002. Phytoremediation, an ecological solution to organic contamination. Ecol Eng, 18:647–58. Yoon, J., C. Xinde, Z. Qixing , and L.Q. Ma. 2006. Accumulation of Pb, Cu, and Zn in Native Plants Growing on a Contaminated Florida Site. Science of the Total Environment: 456-464. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 53-61, Desember 2010 KAJIAN STABILITAS EMPAT TIPE KASKO KAPAL POLE AND LINE STABILITY ANALYSIS OF FOUR TYPES OF POLE AND LINER St. Aisyah Farhum Faculty of Marine and Fisheries Science, Hasanuddin University, Indonesia; email: [email protected] ABSTRACT This study was purposed to compare the stability of four types of pole and liner casco (round bottom, round flat bottom, round sharp bottom and u-v bottom),. The stability value was analyzed by calculating the stability on the curve with the heeling angle of 090°. The stability results of each casco type were then compared with the criteria of minimum standard value derived from IMO (International Maritime Organization). Results showed that the four casco types had greater stability values than IMO standard values. This study found that the stability value of the round –sharp bottom casco was better than the others. Keywords: Stability of fishing boat, Pole and liner, casco ABSTRAK Penelitian ini bertujuan untuk menganalisis perbandingan nilai stabilitas pada empat tipe kasko kapal pole and line, masing-masing round bottom, round flat bottom, round sharp bottom dan u-v bottom. Analisis nilai stabilitas dilakukan melalui perhitungan nilai stabilitas terhadap kurva stabilitas pada sudut kemiringan 0 – 90o. Hasil analisis stabilitas masing-masing bentuk kasko kemudian dibandingkan dengan standar minimum kriteria stabilitas kapal perikanan yang dikeluarkan International Maritim Organization. Hasil penelitian menunjukkan bahwa, keempat bentuk kasko kapal pole and line memiliki nilai stabilitas yang lebih besar dari kriteria stabilitas minimum IMO, dimana tipe kasko round-sharp bottom merupakan tipe kasko kapal yang memiliki nilai stabilitas yang lebih baik dibandingkan tipe kasko yang lain. Kata Kunci: Stabilitas, Kapal Pole and Line, Kasko I. PENDAHULUAN Desain merupakan hal yang penting dalam pembangunan kapal ikan (Fyson, 1985). Sesuai dengan perbedaan jenis kapal ikan, maka desain dan konstruksi kapal dibuat berbeda-beda dengan memperhatikan persyaratan teknis pengoperasian setiap jenis kapal berdasarkan alat tangkap yang dioperasikan. Desain dan konstruksi yang berbeda berdasarkan jenis kapal ikan ini, membuat bentuk badan kapal (kasko) menjadi berbeda-beda (Gillmer and Johnson, 1982 ; Fyson, 1985). Bentuk badan kapal bergantung pada ukuran utama, perbandingan ukuran utama dan koefisien bentuk kapal (Fyson, 1985). Ukuran utama kapal terdiri dari panjang kapal (L), lebar kapal (B), tinggi/dalam kapal (D) dan draft/sarat air kapal (d). Kesesuaian rasio dimensi sangat menentukan kemampuan suatu kapal ikan, karena akan mempengaruhi resistensi kapal (nilai L/B), kekuatan memanjang kapal (nilai L/D) dan stabilitas kapal (nilai B/D) (Fyson, 1985) Stabilitas kapal merupakan kemampuan sebuah kapal untuk kembali ke posisi semula setelah mengalami ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 53 Kajian Stabilitas Empat Tipe Kasko Kapal Pole and Line Penelitian yang menitikberatkan pada kajian stabilitas ini bertujuan untuk mengetahui tingkat kestabilan masingmasing jenis kasko kapal pole and line, sehingga dapat diperoleh informasi mengenai perbandingan nilai stabilitas pada empat tipe kasko kapal pole and line. keolengan. Stabilitas kapal terkait erat dengan distribusi muatan dan perhitungan nilai lengan penegak (GZ). Perbedaan distribusi muatan yang terjadi pada setiap kondisi pemuatan akan mengakibatkan terjadinya perubahan pada nilai KG, yaitu jarak vertikal antara titik K (keel) dan titik G (centre of gravity) yang selanjutnya akan mempengaruhi nilai lengan penegak (GZ) yang terbentuk (Hind, 1982; Derret, 1990). Stabilitas kapal bergantung pada beberapa faktor antara lain dimensi kapal, bentuk badan kapal yang berada di dalam air, distribusi benda-benda di atas kapal dan sudut kemiringan kapal terhadap bidang horizontal (Fyson, 1985). Pada kapal pole and line, keberhasilan pengoperasian sangat bergantung pada kondisi stabilitas kapal karena sifat pengoperasiannya yang oseanik. Kapal pole and line di Sulawesi Selatan, memiliki bentuk badan kapal yang beragam. Berdasarkan hasil penelitian Farhum (2006), ada empat bentuk badan kapal pole and line di Sulawesi Selatan, yaitu round bottom, round flat bottom, round sharp bottom dan u-v bottom. II. METODE PENELITIAN 2.1. Data yang Digunakan Kajian ini menggunakan data: empat karakteristik kapal pole and line dengan masing-masing bentuk kasko round-sharp botttom (K-A), round-flat bottom (K-B), U-V bottom (K-C) dan round bottom (K-D). Spesifikasi sampel kapal yang digunakan untuk keempat bentuk kapal pole and line diterakan pada Tabel 1. 2.2. Analisis Data Perhitungan stabilitas kapal pole and line sampel meliputi analisis terhadap perkiraan perubahan nilai KG pada empat kondisi distribusi muatan terhadap empat bentuk badan kapal, masing-masing adalah: 1) Kondisi kapal kosong; pada kondisi ini bahan bakar, umpan hidup dan muatan diasumsikan kosong (0%). Tabel 1. Spesifikasi Kapal Pole and Line Sampel Sampel K-A K-B K-C K-D LOA (m) 20.44 22.20 21.50 18.40 B (m) 4.06 5.04 3.84 3.52 D (m) 1.75 2.12 1.92 1.80 d (m) 1.40 1.70 1.20 1.44 Keterangan: LOA: lenght over all (panjang keseluruhan kapal) B: Breadth (lebar kapal) D: Depth (dalam/tinggi kapal) D: Draught (tinggi garis air) Cb: Coefficient of block GT: Gross tonnage 54 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Cb 0.46 0.54 0.57 0.43 GT 24.31 30.71 26.82 17.83 Farhum 2) Kondisi kapal berangkat; pada kondisi ini bahan bakar, umpan hidup diasumsikan penuh (100%) dan muatan kosong (0%). 3) Kondisi kapal beroperasi; pada kondisi ini bahan bakar diasumsikan setengah penuh (50%), umpan hidup seperempat penuh (25%) dan muatan tigaperempat penuh (75%). 4) Kondisi kapal pulang; pada kondisi ini bahan bakar diasumsikan seperempat penuh (25%), umpan hidup 10% dan muatan penuh 100%. Perubahan nilai KG dianalisis dengan membuat perkiraan perubahan jarak vertikal – horizontal pada setiap kondisi perubahan distribusi muatan. Nilai KG diperoleh dengan menggunakan formula berikut (Hind, 1982): moment of ∆ Z KG = ∆ dimana: ∆Z adalah momen vertikal Analisis stabilitas statis melalui kurva stabilitas statis GZ dilakukan dengan metode Attwood’s Formula (Hind, 1982). Metode ini menganalisis stabilitas statis kapal pada sudut keolengan 0O - 90O. Hasil perhitungan stabilitas kemudian dibandingkan dengan standar stabilitas kapal yang dikeluarkan oleh United Kingdom Regulations [The Fishing Vessels (Safety Provision) Rules,1975] (Hind, 1982) dan International Maritime Organization (IMO) pada Torremolinos International Convention for The Safety of Fishing Vessels–regulation 28 (1977) melalui kurva GZ. III. HASIL DAN PEMBAHASAN 3.1. Nilai KG Kapal Pole and Line Sampel pada Berbagai Kondisi Muatan Hasil perhitungan terhadap nilai KG kapal yang dibuat pada empat kondisi muatan disajikan pada Tabel 2. Analisis nilai KG dilakukan berdasarkan hasil perkiraan perubahan distribusi muatan pada empat kondisi pemuatan. Dari hasil analisis tersebut terlihat bahwa nilai KG kapal akan berubah jika terjadi perubahan distribusi muatan Jika nilai ton displacement bertambah maka nilai KG kapal akan semakin besar, tetapi nilai GM-nya menjadi lebih kecil. Umumnya nilai KG tertinggi pada keseluruhan kapal sampel berada pada kondisi kapal beroperasi yaitu pada kondisi bahan bakar diasumsikan setengah penuh (50%), umpan hidup satu per empat penuh (25%) dan muatan tiga per empat penuh (75%). Kapal sampel K-A dengan bentuk round sharp dan K-B dengan bentuk round flat memiliki nilai KG yang lebih Tabel 2. Hasil perhitungan perkiraan nilai KG, ton displacement pada empat kondisi distribusi muatan No Kondisi Kapal 1. Kapal Kosong 2. Kapal Berangkat 3. Kapal Beroperasi 4. Kapal Pulang ∆ K-A KG GM ∆ (∆) dan K-B KG GM ∆ K-C KG GM ∆ GM K-D KG GM 66.0 1.90 0.68 78.2 1.87 0.46 44.0 1.45 0.50 32.5 1.35 0.42 87.3 1.95 0.63 101.9 1.92 0.41 59.6 1.50 0.45 47.0 1.37 0.40 87.5 1.97 0.61 99.2 1.94 0.40 58.7 1.51 0.44 45.2 1.40 0.37 87.2 1.96 0.62 96.4 1.93 0.41 57.4 1.52 0.43 44.2 1.39 0.38 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 55 Kajian Stabilitas Empat Tipe Kasko Kapal Pole and Line tinggi dibandingkan kedua kapal sampel lainnya, karena nilai ton displacement kedua kapal tersebut juga lebih besar. Sebuah kapal dapat dikatakan stabil apabila kapal tersebut dapat kembali menjadi tegak setelah mengalami momen kemiringan. Stabilitas pada kapal ikan amat bergantung dari distribusi muatan yang ada pada kapal tersebut. Taylor (1977) dan Hind (1982) menyatakan bahwa stabilitas sebuah kapal dipengaruhi oleh letak ketiga titik konsentrasi gaya yang bekerja pada kapal tersebut. Ketiga titik tersebut adalah titik B (centre of bouyancy), titik G (centre of gravity) dan titik M (metacentre). Selanjutnya Hind (1982) mengemukakan bahwa posisi titik G bergantung dari distribusi muatan dan posisi titik B bergantung pada bentuk kapal yang terendam di dalam air. Saat kapal berangkat menuju daerah penangkapan, muatan pada kapal pole and line terdiri atas perbekalan, bahan bakar dan bak umpan hidup yang terisi penuh. Pada saat kembali, muatanmuatan tersebut akan berkurang tetapi palkah ikan akan terisi oleh hasil tangkapan. Hal ini menyebabkan perubahan titik berat pada kapal, sehingga letak titik G (centre of gravity) kapal akan berubah. Titik berat (G) pada sebuah kapal merupakan titik tangkap dari sebuah titik pusat seluruh gaya berat yang menekan ke bawah. Letak titik G dapat ditentukan dengan meninjau semua pembagian berat yang berada di atas kapal terhadap lunas kapal. Letak titik berat di atas lunas (KG) akan mempengaruhi besar kecilnya nilai lengan penengak GZ yang terbentuk pada saat kapal mengalami keolengan. Berdasarkan hasil perkiraan perubahan distribusi muatan pada keempat bentuk kapal (K-A, K-B, K-C dan K-D) yang diterakan pada Tabel 4.2 memperlihatkan bahwa nilai KG kapal akan berubah jika terjadi perubahan berat 56 dan distribusi muatan. Hal ini juga dijelaskan oleh Hind (1982) bahwa penambahan dan perpindahan muatan pada kapal dapat mengakibatkan perubahan nilai displacement, draft, posisi G, posisi B, posisi M dan trim fore dan aft. Dari Tabel 2 diketahui bahwa, perubahan nilai ton displacement berpengaruh terhadap nilai KG kapal, tetapi tidak menentukan peningkatan dan penurunan nilai tersebut. Peningkatan dan penurunan nilai KG bergantung kepada distribusi muatan yang ada di atas kapal. Hasil penelitian dari Iskandar (1997) juga menjelaskan bahwa tinggi rendah nilai KG tidak bergantung pada nilai ton displacement kapal, tetapi pada kondisi penempatan muatan di atasnya. Berdasarkan Tabel 2 juga diketahui bahwa nilai ton displacement berpengaruh terbalik terhadap nilai tinggi metacentre (GM) yang terbentuk, dimana semakin tinggi nilai ton displacement kapal, maka tinggi metacentre akan menurun. Hal ini dapat dijelaskan pada Gambar 1 bahwa, jika sebuah beban w (ton) meningkatkan draft kapal maka centre of gravity kapal akan meningkat, sehingga terjadi sebuah posisi GG1 yang baru, sehingga tinggi metacentre akan menurun. Kapal sampel K-A dengan bentuk round sharp bottom memiliki nilai KG yang lebih tinggi dibandingkan ketiga Gambar 1. Penambahan beban pada kapal (Sumber: Hind, 1982) http://www.itk.fpik.ipb.ac.id/ej_itkt22 Farhum statis kapal pole and line sampel pada berbagai kondisi distribusi muatan disajikan pada Gambar 2. Nilai lengan penegak GZ yang terbentuk pada kurva GZ berbanding terbalik dengan nilai KG. Pada kurva tersebut terlihat bahwa semakin tinggi nilai KG maka nilai GZ akan semakin rendah, demikian pula sebaliknya. Dari keempat bentuk kapal yang ada, kapal KA dengan bentuk badan round-sharp bottom memiliki nilai GZ yang lebih tinggi dibandingkan kapal sampel dengan bentuk badan kapal yang lain. Nilai lengan penegak GZ menunjukkan nilai stabilitas suatu kapal. Nilai ini memiliki standar yang ditetapkan oleh IMO. Hasil perhitungan stabilitas kapal pole and line sampel yang diterakan pada Tabel 3 sampai Tabel 6, menunjukkan bahwa seluruh nilai lengan penegak GZ kapal pole and line sampel memiliki nilai lebih besar dibandingkan nilai minimum yang ditetapkan oleh IMO. kapal sampel lainnya. Hal ini disebabkan karena bentuk seperti ini memungkinkan untuk mendistribusikan muatan yang lebih banyak pada bagian atas deck sehingga titik berat kapal bergerak ke atas. Dengan demikian nilai KG yang terbentuk menjadi lebih tinggi. Nilai KG tertinggi pada keseluruhan kapal sampel berada pada saat kondisi kapal beroperasi dan pulang, Tingginya nilai KG pada saat kondisi tersebut karena bertambahnya muatan di atas dek oleh hasil tangkapan sedangkan muatan di bawah dek seperti bahan bakar dan umpan hidup menjadi berkurang, sehingga titik G bergerak ke atas. 3.2. Nilai Lengan Penegak GZ Kapal Pole and Line Sampel Stabilitas statis keempat kapal sampel diukur dengan menghitung nilai lengan penegak (GZ) yang terbentuk pada kurva GZ. Pada kurva GZ ditunjukkan nilai GZ pada berbagai sudut keolengan (00 – 800). Kurva stabilitas GZ (m ) 0.35 K-A 0.30 0.25 0.20 0.15 0.10 0.05 0.00 0 10 20 30 40 50 60 70 80 Heel Angle (deg) KG-1 KG-2 KG-3 KG-4 Gambar 2. Kurva GZ Kapal Pole and Line Sampel Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 57 Kajian Stabilitas Empat Tipe Kasko Kapal Pole and Line GZ (m ) 0.35 K-B 0.30 0.25 0.20 0.15 0.10 0.05 0.00 0 10 20 30 40 50 60 70 80 Heel Angle (deg) KG-1 KG-2 KG-3 KG-4 Gambar 2. Kurva GZ Kapal Pole and Line Sampel (lanjutan) GZ (m ) 0.35 K-C 0.30 0.25 0.20 0.15 0.10 0.05 0.00 0 10 20 30 40 50 60 70 80 Heel Angle (deg) KG-1 KG-2 KG-3 KG-4 Gambar 2. Kurva GZ Kapal Pole and Line Sampel (lanjutan) GZ(m ) 0.35 K-D 0.30 0.25 0.20 0.15 0.10 0.05 0.00 0 10 KG-1 20 30 KG-2 40 KG-3 50 60 70 80 Heel Angle (deg) KG-4 Gambar 2. Kurva GZ Kapal Pole and Line Sampel (lanjutan) 58 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Farhum Tabel 3. Nilai kriteria stabilitas kapal pole and line K-A dan nilai standar IMO Nilai Pada Kurva GZ 0B 0 A (0-30 ) B (0-400) C (30-400) D (sudut GZmax) E (GZmin) F (GM) Kondisi Distribusi Muatan 2 3 Standar IMO (Nilai Minimum) 1 0.005 m-rad 0.090 m-rad 0.030 m-rad 30 deg 0.2 m 0.15 m 0.087 0.140 0.052 37 0.305 0.680 0.081 0.128 0.047 36 0.280 0.630 0.078 0.124 0.045 36 0.263 0.610 4 0.079 0.125 0.046 36 0.267 0.620 Tabel 4. Nilai kriteria stabilitas kapal pole and line K-B dan nilai standar IMO Nilai Pada Kurva GZ 2B A (0-300) B (0-400) C (30-400) D (sudut GZmax) E (GZmin) F (GM) Kondisi Distribusi Muatan 2 3 4 Standar IMO (Nilai Minimum) 1 0.005 m-rad 0.090 m-rad 0.030 m-rad 30 deg 0.2 m 0.15 m 0.061 0.095 0.035 37 0.200 0.460 3B 0.054 0.083 0.029 36 0.170 0.410 0.052 0.079 0.028 35 0.160 0.400 0.053 0.082 0.029 35 0.170 0.410 Tabel 5. Nilai kriteria stabilitas kapal pole and line K-C dan nilai standar IMO Nilai Pada Kurva GZ 4B A (0-300) B (0-400) C (30-400) D (sudut GZmax) E (GZmin) F (GM) Standar IMO (Nilai Minimum) 0.005 m-rad 0.090 m-rad 0.030 m-rad 30 deg 0.2 m 0.15 m Kondisi Distribusi Muatan 2 3 5B 1 0.064 0.105 0.041 37 0.240 0.500 0.057 0.093 0.036 35 0.210 0.450 0.056 0.091 0.035 35 0.200 0.440 4 0.055 0.088 0.033 35 0.200 0.430 Tabel 6. Nilai kriteria stabilitas kapal pole and line K-D dan nilai standar IMO Nilai Pada Kurva GZ 6B 0 A (0-30 ) B (0-400) C (30-400) D (sudut GZmax) E (GZmin) F (GM) Standar IMO (Nilai Minimum) 0.005 m-rad 0.090 m-rad 0.030 m-rad 30 deg 0.2 m 0.15 m Berdasarkan kurva stabilitas, diperoleh nilai stabilitas maksimum dan kisaran stabilitas yang diterakan pada Tabel 7. Stabilitas maksimum adalah nilai GZ maksimum yang dapat dicapai oleh kapal pada kondisi tertentu dan terjadi pada besar sudut tertentu. Kisaran Kondisi Distribusi Muatan 2 3 7B 1 0.049 0.073 0.023 30 0.140 0.420 0.047 0.069 0.022 30 0.130 0.400 0.043 0.061 0.019 30 0.120 0.370 4 0.044 0.064 0.019 30 0.120 0.380 stabilitas merupakan sudut terbesar kemiringan kapal tanpa terjadinya nilai GZ yang negatif. Besar sudut ini diketahui dari titik potong kurva GZ dengan sumbu X (axis), dimana nilai GZ sama dengan 0 dan disebut dengan angle of vanishing stability. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 59 Kajian Stabilitas Empat Tipe Kasko Kapal Pole and Line Tabel 7. Nilai maksimum dan kisaran stabilitas kapal pole and line sampel No. Kapal 1. K-A 2. K-B 3. K-C 4. K-A Kondisi Kapal Kosong Kapal Berangkat Kapal Beroperasi Kapal Pulang Kapal Kosong Kapal Berangkat Kapal Beroperasi Kapal Pulang Kapal Kosong Kapal Berangkat Kapal Beroperasi Kapal Pulang Kapal Kosong Kapal Berangkat Kapal Beroperasi Kapal Pulang Maksimum Stabilitas Sudut (º) GZ (m) 37 0.305 36 0.280 36 0.263 36 0.267 37 0.200 36 0.170 35 0.160 35 0.267 37 0.240 35 0.210 35 0.200 35 0.200 30 0.140 30 0.130 30 0.120 30 0.120 Ada dua gaya yang mengatur kestabilan kapal di laut, yaitu gaya berat (forces of gravity, G) yang selalu bergerak vertikal ke bawah dan gaya apung (forces of bouyancy, B) yang bergerak vertikal ke atas. Pada saat kapal dalam kondisi tenang, kedua gaya ini berada pada satu garis vertikal yang sama. Pada saat kapal mengalami keolengan, gaya berat dan gaya apung kapal akan bergerak ke arah yang berlawanan. Jarak perpendicular yang dibentuk oleh kedua garis gaya ini disebut lengan penegak (GZ). (Gillmer dan Johnson, 1982). Perhitungan stabilitas statis kapal pole and line sampel dilakukan dengan analisis nilai GZ kapal pada beberapa sudut keolengan yang sesuai dengan aturan yang dikeluarkan oleh IMO. Nilai lengan GZ kapal pole and line sampel yang disajikan pada Tabel 3 – 6 memiliki nilai yang lebih besar dibandingkan nilai minimum yang ditetapkan oleh IMO. Hal ini dapat dilihat dari nilai margin yang positif (Tabel 7). Kondisi tersebut menunjukkan bahwa pada keempat kondisi pemuatan, kapal dapat menghasilkan momen kopel yang positif untuk mengembalikan kapal 60 Sudut Kisaran Stabilitas (º) 0 – 80 0 – 77 0 – 74 0 – 75 0 – 72 0 – 67 0 – 65 0 – 67 0 – 80 0 – 75 0 – 74 0 – 72 0 – 70 0 – 67 0 – 62 0 – 64 ke posisi semula setelah terjadi oleng akibat gaya yang bekerja padanya. Nilai GZ akan menjadi negatif jika sudut keolengan lebih besar dari batas nilai maksimum kisaran stabilitas (Tabel 7), yang mengakibatkan kapal tidak lagi menghasilkan lengan GZ yang positif. Bila hal ini terjadi kapal akan terbalik karena saat terjadi keolengan pada sudut tersebut. Kapal dengan lengan GZ negatif akan meneruskan geraknya ke arah kemiringannya dan tidak kembali ke posisi semula. IV. KESIMPULAN Lengan penegak GZ yang terbentuk pada keempat kapal pole and line sampel pada setiap kondisi pemuatan bernilai positif dan berada di atas nilai standar minimum yang ditetapkan oleh IMO, yang berarti nilai lengan penegak GZ yang dihasilkan masih dapat mengembalikan kapal ke posisi semula setelah terjadi keolengan, sehingga dapat disimpulkan bahwa keempat kapal sampel tersebut memiliki stabilitas yang baik. Kapal sampel K-A dengan bentuk round-sharp bottom memiliki nilai kriteria stabilitas yang lebih tinggi http://www.itk.fpik.ipb.ac.id/ej_itkt22 Farhum dibandingkan lainnya. ketiga bentuk kapal DAFTAR PUSTAKA Derret, D.R. 1990. Ship Stability for Masters and Mates. Fourth Edition, Revised. Butler & Tanner Ltd, Frome and London. Farhum, S.A. 2006. Kajian Stabilitas dan Keselamatan Pengoperasian Kapal Pole and Line pada Kondisi Gelombang Beam Seas. Disertasi pada Program Pascasarjana IPB. Bogor. Farhum, S.A. 2007. Distribusi Statistik Karakteristik Teknis Kapal Pole and Line Sulawesi Selatan. Jurnal Torani, 3(17):246-253 Fyson, J. 1985. Design of Small Fishing Vessels. Fishing News (Books) Ltd. England. Gillmer, T.C and B. Johnson. 1982. Introduction to Naval Architecture. Naval Institute Press. Annapolis. Maryland. Hind, J.A. 1982. Stability and Trim of Fishing Vessels and Other Small Ships. Second Edition. Fishing News Books Ltd. Farnham, Surrey, England. International Maritime Organization (IMO). 1983. International Confrence on Safety Fishing Vessels 1977. IMO. London. Iskandar, B.H. 1997. Studi tentang Desain Kapal Kayu Mina Jaya BPPT 01. Tesis pada Program Pascasarjana IPB. Bogor. Kok, H.G.M, E.G.V. Lonkhyusen, and F.A.C. Nierich. 1983. Bangunan Kapal. Zundort. Netherland. Taylor, L.G. 1977. The Principles of Ship Stability. Brown, Son & Publisher, Ltd., Nautical Publisher, 52 Darnley Street. Glasgow. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 61 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 62-73, Desember 2010 STRUKTUR KOMUNITAS IKAN PADA PADANG LAMUN YANG BERBEDA DI PULAU BARRANG LOMPO FISH COMMUNITY STRUCTURE IN DIFFERENT SEAGRASS BEDS OF BARRANG LOMPO ISLAND Rohani Ambo Rappe Marine Science Department, Hasanuddin University, Makassar 90245 Email: [email protected] ABSTRACT The importance of seagrass meadows as a habitat for fishes, including several of economic importance, is widely acknowledged. The complexity of seagrass beds might offer a different condition of habitat for fishes. The physical nature of the seagrass canopy is thought to play a major role, potentially influencing available shelter, food, and protection from predators. Structural complexity of seagrass such as shoot and leaf density is also an important factor in determining ecological function of seagrass in the marine environment. The objective of the research is to assess the ecological function of different seagrass beds (in terms of spesies and density) in supporting fish community. The study found 28 species of fish originating from 14 families and Pomacentridae were dominantly found. Abundance of fish found to be higher in seagrass beds with high densities both composed by one species of seagrass (monospesific) or by more than one species of seagrass (multispesific), compared to the seagrass beds with low density and bare areas. Fish community diversity index was found higher in dense seagrass beds composed of many species of seagrass compared to the rare and consists of only one species of seagrass. The presence of epiphytes as nutrients for the fish that live in seagrass beds may contribute to the finding. Keywords: Seagrass, fish, Barrang Lompo Island ABSTRAK Peranan padang lamun terhadap keberadaan ikan terutama yang bernilai ekonomis penting, sudah sering dilaporkan. Hal ini terkait dengan kompleksitas dari padang lamun yang dapat menyediakan makanan dan perlindungan dari predator bagi ikan-ikan tersebut. Kompleksitas padang lamun dapat diukur dari kepadatan tegakan dan daun penyusunnya, penutupan daun, serta jenis-jenis lamun penyusun padang lamun tersebut. Tujuan dari penelitian ini adalah untuk mengevaluasi fungsi ekologis dari padang lamun yang berbeda (dalam hal perbedaan spesies lamun penyusun dan kerapatan lamun) dalam mendukung keberadaan komunitas ikan. Penelitian ini menemukan 28 spesies ikan yang berasal dari 14 famili dan Pomacentridae adalah famili yang dominan ditemukan. Kelimpahan ikan ditemukan lebih tinggi pada padang lamun dengan kerapatan yang tinggi baik itu tersusun oleh satu spesies lamun (monospesifik) maupun oleh lebih dari satu spesies lamun (multispesific), dibandingkan pada padang lamun dengan kerapatan rendah dan pada daerah tidak bervegetasi. Nilai indeks keanekaragaman komunitas ikan ditemukan lebih tinggi pada padang lamun yang rapat dan tersusun oleh banyak spesies lamun dibandingkan pada padang lamun jarang dan hanya terdiri dari satu spesies lamun. Keberadaan epifit sebagai nutrisi bagi ikan yang hidup di padang lamun dapat berkontribusi terhadap hasil yang dicapai. Kata Kunci: Padang lamun, ikan, Pulau Barrang Lompo 62 ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Rappe I. PENDAHULUAN Lamun (seagrass) adalah satusatunya tumbuh-tumbuhan berbunga yang terdapat di lingkungan laut. Seperti halnya rumput di darat, mereka mempunyai tunas berdaun yang tegak dan tangkai-tangkai yang merayap efektif untuk berkembang-biak dan mempunyai akar dan sistem internal untuk mengangkut gas dan zat-zat hara (Romimohtarto dan Juwana, 2001) Lamun juga merupakan tumbuhan yang telah menyesuaikan diri hidup terbenam di laut dangkal. Lamun mempunyai akar dan rimpang (rhizome) yang mencengkeram dasar laut sehingga dapat membantu pertahanan pantai dari gerusan ombak dan gelombang. Padang lamun dapat terdiri dari vegetasi lamun jenis tunggal ataupun jenis campuran (Hemminga and Duarte, 2000). Padang lamun memiliki produktivitas sekunder dan dukungan yang besar terhadap kelimpahan dan keragaman ikan (Gilanders, 2006). Padang lamun merupakan tempat berbagai jenis ikan berlindung, mencari makan, bertelur, dan membesarkan anaknya. Ikan baronang, misalnya, adalah salah satu jenis ikan yang hidup di padang lamun. Bell dan Pollard (1989) mengidentifikasi 7 karakteristik utama kumpulan ikan yang berasosiasi dengan lamun yaitu: (1) Keanekaragaman dan kelimpahan ikan di padang lamun biasanya lebih tinggi daripada yang berdekatan dengan substrat kosong, (2) Lamanya asosiasi ikan-lamun berbedabeda diantara spesies dan tingkatan siklus hidup, (3) Sebagian besar asosiasi ikan dengan padang lamun didapatkan dari plankton, jadi padang lamun adalah daerah asuhan untuk banyak spesies yang mempunyai nilai ekonomi penting, (4) Zooplankton dan epifauna krustasean adalah makanan utama ikan yang berasosiasi dengan lamun, dengan tumbuhan, pengurai dan komponen infauna dari jaring-jaring makanan di lamun yang dimanfaatkan oleh ikan, (5) Perbedaan yang jelas (pembagian sumberdaya) pada komposisi spesies terjadi di banyak padang lamun, (6) Hubungan yang kuat terjadi antara padang lamun dan habitat yang berbatasan, kelimpahan relatif dan komposisi spesies ikan di padang lamun menjadi tergantung pada tipe (terumbu karang, estuaria, mangrove) dan jarak dari habitat yang terdekat, (7) Kumpulan ikan dari padang lamun yang berbeda seringkali berbeda juga, walaupun dua habitat itu berdekatan. Hasil penelitian Radjab et al. (1992) menemukan 1.588 jumlah individu ikan yang terdiri dari 61 spesies yang mewakili 10 suku di areal padang lamun Teluk Baguala, khususnya di perairan Passo. Sedangkan hasil penelitian Rani et al. (2010) pada areal lamun buatan menemukan bahwa ikan memilih padang lamun dengan struktur yang lebih kompleks dibandingkan struktur yang sederhana. Oleh karena itu peneliti marasa perlu untuk membuktikan pengaruh keberadaan padang lamun dengan tingkat kompleksitas yang berbeda terhadap kelimpahan dan keragaman jenis ikan. Adapun tujuan penelitian ini adalah untuk mengetahui keanekaragaman jenis dan kelimpahan ikan pada padang lamun yang berbeda kerapatan dan komposisi jenisnya. II. METODE PENELITIAN 2.1. Lokasi dan waktu pengamatan Penelitian ini dilakukan pada bulan April 2010 pada daerah padang lamun perairan Pulau Barrang Lompo, Kota Makassar (Gambar 1). Terdapat 5 stasiun pengamatan yang ditetapkan berdasarkan tingkat kompleksitas yang berbeda berdasarkan kerapatan dan jenis lamun penyusunnya, yaitu: (1) LPU; lamun Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 63 Struktur Komunitas Ikan Pada Padang Lamun… padat multispesifik, (2) LPO; lamun padat monospesifik, (3) LJU; lamun jarang multispesifik; (4) LJO; lamun jarang monospesifik, dan (5) LNV; daerah tidak bervegetasi. Terdapat 3 ulangan untuk setiap stasiun. Masing-masing stasiun dibatasi menggunakan tali untuk membuat area pengamatan seluas 10m x 10m. Pengambilan data ikan meliputi pengamatan jenis dan jumlah ikan dilakukan dalam area pengamatan (10m x 10m) pada setiap stasiun. Pengamatan ini dilakukan pada saat air pasang dengan metode sensus visual dengan bantuan kamera bawah air mengikuti Edgar et al. (2001). Metode ini dapat dilakukan untuk mengambil data ikan yang berukuran cukup besar pada daerah padang lamun di perairan dangkal dengan kecerahan air yang tinggi. Pengukuran parameter lingkungan meliputi pengukuran suhu, salinitas, pH, kedalaman dan kecerahan perairan dilakukan secara in situ pada setiap stasiun pengamatan. Adapun biomassa epifit diukur dengan mengambil secara acak 3 sampel daun lamun pada setiap stasiun pengamatan menggunakan kuadrat 20cm x 20cm. Epifit diserut dari permukaan daun lamun kemudian dikeringkan dalam oven dengan suhu 60oC selama 48 jam, kemudian ditimbang. Metode pengukuran biomassa epifit ini mengikuti Sidik et al. (2001). 2.2. Pengolahan data Parameter yang diamati untuk data ikan adalah kelimpahan, komposisi jenis (KJ), indeks keanekaragaman (H’), Indeks keseragaman (E), dan Indeks dominansi (C). Komposisi jenis adalah perbandingan antara jumlah jenis tiap suku dengan jumlah seluruh jenis yang Gambar 1. Peta Lokasi Penelitian 64 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Rappe ditemukan berikut: dengan KJ = formula sebagai ni x 100% N dimana: KJ = Komposisi jenis (%) Ni = Jumlah individu setiap jenis N = Jumlah individu seluruh jenis Indeks keanekaragaman adalah nilai yang dapat menunjukkan keseimbangan keanekaragaman dalam suatu pembagian jumlah individu tiap jenis. Sedikit atau banyaknya keanekaragaman spesies dapat dilihat dengan menggunakan indeks keanekaragaman (H’). Keanekaragaman (H') mempunyai nilai terbesar jika semua individu berasal dari genus atau spesies yang berbeda-beda. Sedangkan nilai terkecil didapat jika semua individu berasal dari satu genus atau satu spesies saja (Odum, 1983). Adapun kategori Indeks Keanekaragaman dapat dilihat pada Tabel 1. Adapun indeks keanekaragaman Shannon (H’) menurut Shannon and Weaver (1949) dalam Odum (1983) dihitung menggunakan formula sebagai berikut: H’ = -∑ (ni/N)ln(ni/N) dimana: ni = Jumlah individu setiap jenis N = Jumlah individu seluruh jenis Pengujian juga dilakukan dengan pendugaan indeks keseragaman (E), dimana semakin besar nilai E menunjukkan kelimpahan yang hampir seragam dan merata antar jenis (Odum, 1983). Adapun kriteria komunitas lingkungan berdasarkan nilai indeks keseragaman disajikan pada Tabel 2. Rumus dari indeks keseragaman Pielou (E) menurut Pielou (1966) dalam Odum (1983) yaitu: H' E= ln S dimana: E = Indeks keseragaman H’ = Indeks keanekaragaman S = Jumlah jenis Nilai dari indeks dominansi Simpson memberikan gambaran tentang dominansi organisme dalam suatu komunitas ekologi. Indeks ini dapat menerangkan bilamana suatu jenis lebih banyak terdapat selama pengambilan data. Adapun kategori penilaiannya disajikan pada Tabel 3. Tabel 1. Kategori Indeks Keanekaragaman Nilai Keanekaragaman (H’) H’ ≤ 2,0 2,0 < H’ ≤ 3,0 H’ ≥ 3,0 Kategori Rendah Sedang Tinggi Tabel 2. Kriteria Komunitas Lingkungan Berdasarkan Nilai Indeks Keseragaman Nilai Indeks Keseragaman (E) 0,00 < E ≤ 0,50 0,50 < E ≤ 0,75 0,75 < E ≤ 1,00 Kondisi Komunitas Komunitas berada pada kondisi tertekan Komunitas berada pada kondisi labil Komunitas berada pada kondisi stabil Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 65 Struktur Komunitas Ikan Pada Padang Lamun… Tabel 3. Kategori Indeks Dominansi Dominansi (C) 0,00 < C ≤ 0,50 0,50 < C ≤ 0,75 0,75 < C ≤ 1,00 Kategori Rendah Sedang Tinggi Rumus indeks dominansi Simpson (C) menurut Margalef (1958) dalam Odum (1983) yaitu: C = ∑(ni/N)2 dimana: C = Indeks dominansi Simpson ni = Jumlah individu spesies ke-i N = Jumlah individu seluruh spesies 2.3. Analisis data Perbedaan kelimpahan ikan antara stasiun pengamatan dianalisis menggunakan uji statistik One-way ANOVA dengan bantuan paket program SPSS (Statistical Product and Service Solutions). III. HASIL DAN PEMBAHASAN 3.1. Kelimpahan ikan pada daerah padang lamun Kelimpahan ikan ditemukan berbeda antar stasiun pengamatan (p<0,05). Hasil uji lanjut menunjukkan Kelimpahan Ikan (ekor/100 m²) 90 80 a 70 bahwa ikan lebih melimpah pada daerah padang lamun dengan kerapatan tinggi baik monospesifik (hanya tersusun oleh satu jenis lamun; LPO) maupun multispesifik (tersusun oleh lebih dari satu jenis lamun; LPU) dibandingkan pada padang lamun jarang terutama monospesifik (LJO) maupun daerah yang tidak bervegetasi (LNV) (Gambar 2). Menurut Hemminga and Duarte (2000), padang lamun terutama dengan kerapatan yang tinggi menyediakan perlindungan bagi ikan dari serangan predator, selain itu kerapatan lamun yang tinggi tentunya meningkatkan luas permukaan bagi perlekatan hewan-hewan maupun tumbuhan renik yang merupakan makanan utama bagi ikan-ikan di padang lamun. 3.2. Komposisi jenis Hasil penelitian pada ekosistem padang lamun di perairan Pulau Barrang Lompo secara keseluruhan ditemukan 28 spesies ikan yang berasal dari 14 famili yaitu 1 spesies dari famili Gerreidae, 3 spesies dari Siganidae, 2 spesies dari Labridae, 8 spesies dari Pomacentridae, 3 spesies dari Nemipteridae, 2 spesies dari Gobiidae, 2 spesies dari Apogonidae, dan masing-masing 1 spesies dari Sphyraenidae, Muraenidae, Monachanti dae, Tetraodontidae, Hemiramphidae, a 60 ab 50 40 b 30 b 20 10 0 LPU LPO LJU LJO LNV Gambar 2. Kelimpahan ikan (mean±SE, n=3) pada setiap stasiun pengamatan 66 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Rappe Serranidae, dan Acanthuridae. Ketersediaan pangan dan tempat perlindungan dari predator juga menjadikan sejumlah besar organisme termasuk ikan hidup pada padang lamun (Gilanders, 2006). Adapun jenis yang ditemukan maupun tidak ditemukan pada setiap stasiun pengamatan disajikan pada Tabel 4. Jumlah jenis ikan yang ditemukan pada penelitian ini lebih tinggi jika dibandingkan hasil yang didapatkan peneliti sebelumnya di lokasi yang sama yaitu Erftemeijer and Allen (1993) dan Supriadi et al. (2004) yang menemukan berturut-turut 27 dan 19 spesies. Jika dibandingkan dengan hasil penelitian di tempat lain, jumlah jenis ikan yang ditemukan di daerah padang lamun Pulau Barrang Lompo ini masih lebih rendah dibandingkan dengan yang ditemukan di daerah padang lamun Pulau Wakatobi Marine National Park sebanyak 81 jenis (Unsworth et al., 2007). Hal ini dikarenakan areal pengamatan di Wakatobi yang lebih luas dan terutama teknik pengambilan data ikan yang berbeda, dimana teknik “beach seining” digunakan di Wakatobi dan teknik “visual sensus” digunakan pada penelitian ini. Beach seine dapat menjaring ikan yang jauh lebih banyak termasuk ikan-ikan yang bersembunyi di antara rhizoma dan daun lamun, yang kemungkinan tidak terdata pada saat teknik visual sensus diterapkan. Dari hasil pengamatan, ikan yang dominan ditemukan di daerah padang lamun Pulau Barrang Lompo ini juga banyak ditemukan pada daerah terumbu karang (Kuiter and Tonozuka, 1992; Azis, 2002). Hal yang sama ditemukan sebelumnya oleh Erftemeijer and Allen (1993) dan Supriadi et al. (2004). Pola yang serupa juga ditemukan oleh Unsworth et al. (2007) di Taman Laut Nasional Wakatobi. Menurut Kikuchi dan Peres (1977), padang lamun (seagrass beds) dapat juga sebagai daerah asuhan, padang penggembalaan dan mencari makan bagi berbagai jenis ikan herbivora dan ikan-ikan karang (coral fishes). Hal ini didukung pula oleh karena daerah padang lamun perairan Pulau Barrang Lompo merupakan areal yang bersambungan langsung dengan daerah terumbu karang (seagrass associated reef system). Stasiun pengamatan LPU (lamun padat multispesifik) memiliki jumlah jenis ikan yang tertinggi yaitu 21 jenis dibanding stasiun lain, dan yang terendah adalah pada stasiun LNV (daerah tidak bervegetasi) yaitu hanya ditemukan 4 jenis ikan (Tabel 4). Hasil tersebut menunjukkan bahwa secara umum ikan memilih berada pada daerah padang lamun dibandingkan pada daerah kosong yang tidak bervegetasi kemungkinan berkaitan dengan tersedianya perlindungan dan makanan pada daerah padang lamun untuk ikanikan tersebut. Lebih spesifik untuk daerah padang lamun yang berbeda, ikanikan memilih padang lamun dengan susunan yang lebih kompleks (yaitu kerapatan tinggi dan terdiri dari banyak spesies lamun) seperti pada stasiun LPU. Hal ini sejalan dengan hasil penelitian Rani et al. (2010) pada lamun buatan yang mendapatkan bahwa lamun buatan dengan struktur yang lebih kompleks menarik lebih banyak jenis ikan dibandingkan lamun buatan dengan struktur yang lebih sederhana. Padang lamun multispesifik di Pulau Barrang Lompo tersusun atas enam spesies lamun yaitu Enhalus acoroides, Thalassia hemprichii, Halophila ovalis, Syringodium isotifolium, Cymodocea rotundata, dan Halodule pinifolia (Amran and Ambo Rappe, 2009). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 67 Struktur Komunitas Ikan Pada Padang Lamun… Tabel 4. Jenis ikan yang ditemukan pada setiap stasiun pengamatan Famili Spesies Gerreidae Siganidae Labridae Pomacentridae LPU LPO LJU LJO LNV Gerres oyena + + + - + Siganus margaritiferus + + - + - Siganus canaliculatus + + + + - Siganus virgatus - + - - Halichoeres chloropterus + + + + - Novaculichthys sp + - + + - Pomacentrus simsiang + + + - Pomacentrus saksoni + + + - + Pomacentrus sp. - - + - - Dischistodus perspicillatus + + - + + Dischistodus fasciatus + + - - - Dischistodus sp. - - - + - Abudefduf vaigiensis + + + - - Amphiprion ocellaris + - - - - Pentapodus trivittatus + + - + - Pentapodus bifasciatus + + + + - Scolopsis sp. + + - + - Cryptocentrus cinctus + + + + - Cryptocentrus sp. + + + + + Apogon cyanosoma - - + - - Apogon males - + - + - Sphyraenidae Sphyraena barracuda + - - - - Muraenidae Gymnothorax sp. + - - - - Monachantidae Acreichthys tomentosus + - - - - Tetraodontidae Arothron manilensis - - - + - Hemiramphidae Tylosurus sp. + + + + - Serranidae Ephinephelus ongus - + - - - Acanthuridae Acanthurus auranticafus + - - - - Jumlah Jenis 21 16 14 15 4 Nemipteridae Gobiidae Apogonidae Ket : + = Ditemukan, - = Tidak Ditemukan LPU = lamun padat multispesifik, LPO = lamun padat monospesifik, LJU = lamun jarang multispesifik, LJO = lamun jarang monospesifik, LNV = daerah tidak bervegetasi Keenam jenis lamun ini mempunyai bentuk morfologi yang berbeda-beda dalam hal bentuk dan ukuran daun. Hal inilah yang meningkatkan kompleksitas padang lamun multispesifik sehingga dapat menawarkan perlindungan dan penyediaan makanan yang lebih optimal bagi ikan-ikan di padang lamun tersebut. 68 Komposisi jenis ikan pada stasiun LPU didominasi oleh 4 jenis yaitu Siganus margaritiferus (20%), Gerres oyena (19%), Pentapodus bifasciatus (14%), dan Tylosurus sp (12%). Sedangkan pada stasiun LPO, komposisi jenis ikan tertinggi ada pada Siganus canalicatus (46%) yang disusul Siganus margaritiferus (27%) (Gambar 3). http://www.itk.fpik.ipb.ac.id/ej_itkt22 Rappe Tingginya persentasi komposisi jenis Siganus canalicatus dan Siganus margaritiferus pada stasiun LPO diduga disebabkan antara lain karena ikan tersebut memiliki kebiasaan hidup bergerombol di daerah padang lamun, terutama lamun monospesifik yang hanya disusun oleh jenis Enhalus acoroides. Sesuai dengan pernyataan Darsono dan Prapto (1993) sebagian besar jenis Siganus (Siganidae) hidup menggerombol (schooling). Komposisi jenis tertinggi pada stasiun LJU adalah Siganus canalicatus (27%) namun tidak berbeda jauh dengan spesies lain. Sedangkan pada stasiun LJO, komposisi jenis dengan persentase tertinggi terdapat spesies Cryptocentrus sp. Walaupun stasiun LJU dan LJO memiliki kepadatan lamun yang jarang namun masih mendukung keberadaan berbagai jenis ikan, yaitu 14 jenis pada LJU dan 15 jenis pada LJO. Adapun komposisi jenis ikan pada stasiun LNV sangat kurang yaitu hanya ditemukan 4 spesies (Cryptocentrus sp, Dischistodus perspicillatus, Gerres oyena, dan Pomacentrus saksoni) (Gambar 3). Rendahnya jumlah spesies disebabkan karena stasiun ini tidak bervegetasi, hal ini sesuai dengan perrnyataan Hemminga and Duarte (2000) bahwa keanekaragaman dan kelimpahan ikan di padang lamun biasanya lebih tinggi daripada yang berdekatan dengan substrat kosong. Meskipun didominasi oleh ikan yang berasal dari terumbu karang, pada penelitian ini teridentifikasi 2 spesies yang khas ditemukan pada daerah padang lamun Pulau Barrang Lompo, yaitu Acreichthys tomentosus dan Novaculichthys sp, sesuai dengan Erftemeijer and Allen (1993). Pada penelitian ini ditemukan pula 2 spesies ikan bernilai ekonomis penting yang menghuni daerah padang lamun Pulau Barrang Lompo yaitu Siganus canaliculatus dan Sphyraena barracuda. 3.3. Indeks keanekaragaman, keseragaman, dan dominansi Indeks keanekaragaman, keseragaman, dan dominansi menunjukkan keseimbangan dalam pembagian jumlah individu setiap jenis dan juga menunjukkan kekayaan jenis (Odum, 1983). Hasil analisa data untuk indeks keanekaragaman (H’), indeks keseragaman (E) dan indeks dominansi (C) ikan yang ditemukan selama penelitian dapat dilihat pada Tabel 5. Tabel 5. Nilai indeks keanekaragaman, keseragaman, dan dominansi komunitas ikan pada daerah padang lamun Pulau Barrang Lompo Stasiun Indeks Keanekaragaman (H') Indeks Keseragaman (E) Indeks Dominansi (C) LPU 2,44 0,80 0,12 LPO 1,65 0,60 0,29 LJU 2,24 0,85 0,14 LJO 2,19 0,81 0,16 LNV 1,10 0,79 0,41 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 69 Struktur Komunitas Ikan Pada Padang Lamun… LPU Abudefduf vaigiensis 2% Acanthurus auranticafus 2% Pentapodus trivittatus 4% Amphiprion ocellaris 1% Scolopsis sp. 2% LPO Halichoeres chloropterus 7% Tylosurus sp. 12% Pomacentrus saksoni 1% Gerres oyena 7% Siganus margaritiferus 20% Pentapodus bifasciatus 14% Apogon males 1% Scolopsis sp. 1% Tylosurus sp. 1% Cryptocentrus sp. 4% Siganus canalicatus 1% Siganus canaliculatus 46% Cryptocentrus cinctus 2% Pentapodus bifasciatus 3% Pentapodus trivittatus 1% Gerres oyena 19% Sphyraena Acreichthys Cryptocentrus barracuda tomentosus cinctus 1% 1% 3% Dischistodus Gymnothorax sp. fasciatus 1% Pomacentrus 1% Dischistodus Pomacentrus simsiang Halichoeres Cryptocentrus sp. perspicillatus Novaculichthys sp saksoni 4% chloropterus 4% 1% 1% 7% 4% Siganus margaritiferus 27% LJU Novaculichthys sp 2% Pomacentrus sp. 1% Pomacentrus saksoni 3% LJO Tylosurus sp. 2% Apogon cyanosoma 12% Apogon males 1% Pentapodus bifasciatus 12% Novaculichthys sp 6% Cryptocentrus cinctus 5% Cryptocentrus sp. 12% Pomacentrus simsiang 1% Siganus virgatus 3% Dischistodus perspicillatus Dischistodus 1% fasciatus Abudefduf 1% vaigiensis Ephinephelus ongus 1% 1% Siganus canaliculatus 27% Siganus margaritiferus 1% Gerres oyena 3% Abudefduf vaigiensis Halichoeres 7% Scolopsis sp. 10% Siganus canalicatus 17% Pentapodus trivittatus 12% Arothron Pentapodus manilensis Halichoeres Dischistodus sp. bifasciatus chloropterus 1% 3% 4% 3% chloropterus 10% Cryptocentrus cinctus 6% Cryptocentrus sp. 30% Tylosurus sp. 2% Pomacentrus simsiang 2% Dischistodus perspicillatus 2% LNV Gerres oyena 13% Cryptocentrus sp. 59% Dischistodus perspicillatus 20% Pomacentrus saksoni 8% Gambar 3. Komposisi jenis ikan pada setiap stasiun pengamatan Nilai indeks keanekaragaman ikan pada semua stasiun berkisar antara 1,10 – 2,44. Berdasarkan kriteria indeks keanekaragaman, pada stasiun pengamatan LPO dan LNV masih tergolong rendah sedangkan LPU, LJU, LJO tergolong sedang. 70 Rendahnya keanekaragaman pada stasiun LNV disebabkan oleh sedikitnya jumlah spesies ikan yang ditemukan, yaitu hanya ditemukan 4 spesies, dan kecenderungan indeks dominansi yang cukup besar. Hal ini disebabkan stasiun pengamatan tidak bervegetasi sehingga http://www.itk.fpik.ipb.ac.id/ej_itkt22 Rappe tidak ditemukan banyak spesies ikan serta adanya kemungkinan dominansi oleh spesies tertentu yaitu Cryptocentrus sp (lihat Gambar 3). Sedangkan pada stasiun LPO didapatkan nilai indeks keanekaragaman yang rendah meskipun jumlah jenis cukup banyak. Hal ini disebabkan indeks keseragaman pada stasiun LPO termasuk dalam kategori komunitas yang labil, hal ini menunjukkan kemerataan jumlah individu untuk setiap jenis ikan di stasiun LPO rendah. 3.4. Keberadaan ikan, epifit, dan faktor lingkungan di daerah padang lamun Adapun data parameter lingkungan yang diukur seperti suhu (28,8 – 32,0oC), salinitas (29 – 31 ppt), pH (7,80 – 8,14), kedalaman (83 – 283 cm), dan kecerahan perairan pada semua stasiun mencapai 100%. Parameter lingkungan tersebut tidak berpengaruh terhadap hasil yang dicapai. Tingginya kelimpahan dan jumlah jenis ikan yang didapatkan pada stasiun pengamatan LPU, lamun padat multispesifik, lebih berkaitan dengan ketersediaan makanan yang tinggi di daerah tersebut. Hal ini didukung oleh data biomassa epifit yang diperoleh, dimana biomassa epifit tertinggi didapatkan pada stasiun pengamatan LPU, padang lamun dengan kerapatan tinggi dan tersusun atas kurang lebih 6 spesies lamun (Gambar 4). Epifit pada lamun adalah seluruh organisme autotrofik (yaitu, produsen primer) yang menempel pada rhizoma, batang dan daun lamun. Epifit merupakan produsen primer yang penting dalam ekosistem lamun dan memberikan konstribusi yang signifikan dalam rantai makanan. Konstribusi epifit bisa mencapai lebih dari 50% dalam rantai makanan di padang lamun. (Borowitzka et al., 2006). IV. KESIMPULAN Padang lamun dengan tingkat kompleksitas yang berbeda (dapat diukur dari tingkat kerapatan dan banyaknya jenis lamun penyusun) berpengaruh terhadap keberadaan ikan di daerah tersebut. Kelimpahan ikan ditemukan lebih tinggi pada padang lamun dengan kerapatan yang tinggi baik itu tersusun oleh satu spesies lamun (monospesifik) maupun oleh lebih dari satu spesies 0,7000 BIOMASSA EPIFIT (GRAM) 0,6000 0,5000 0,4000 0,3000 0,2000 0,1000 0,0000 Gambar 4. Biomassa epifit (mean±SE, n=3) pada setiap stasiun pengamatan Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 71 Struktur Komunitas Ikan Pada Padang Lamun… lamun (multispesific), dibandingkan pada padang lamun dengan kerapatan rendah dan pada daerah tidak bervegetasi. Nilai indeks keanekaragaman dan keseragaman komunitas ikan yang lebih tinggi dengan indeks dominansi yang rendah ditemukan pada padang lamun yang rapat dan tersusun oleh banyak spesies lamun. DAFTAR PUSTAKA Amran, M.A. and R.A. Rappe. 2009. Estimation of Seagrass Coverage by Depth Invariant Indices on Quickbird Imagery. Research Report DIPA Biotrop 2009. Aziz, A.W. 2002. Studi Kelimpahan dan Keanekaragaman Ikan Karang Famili Pomacentridae dan Labridae pada Daerah Rataan Terumbu (Reef Flat) di Perairan Pulau Barrang Lompo. Skripsi. Program Studi Ilmu Kelautan. Jurusan Ilmu Kelautan. Fakultas Ilmu Kelautan dan Perikanan. Universitas Hasanuddin, Makassar. Bell, J.D. and D.A. Pollard. 1989. Ecology of Fish Assemblages and Fisheries Associated with Seagrasses. In: Larkum, A.W.D., McComb, A.J., and Shepherd, S.A. (Eds.), Biology of Seagrasses: A Treatise on the Biology of Seagrasses with Special Reference to the Australasian Region. Elsevier, Amsterdam, 565– 609pp. Borowitzka, A.M., S.P. Lavery, and V.M. Keulen. 2006. Epiphytes of Seagrasses. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.), Seagrasses: Biology, Ecology, and Conservation. Springer, The Netherland, 441-461pp. Darsono dan Prapto. 1993. Culture Potential Of Rabbitfishes, Siganus (Siganidae) . Bidang Sumberdaya Laut, P2O-LIPI. 72 Edgar, G.J., H. Mukai, and R.J. Orth. 2001. Fish, Crabs, Shrimps and Other Large Mobile Epibenthos: Measurement Methods for Their Biomass and Abundance in Seagrass. In: Short, F.T., Coles, R.G. and Short, C.A. (Eds.), Global Seagrass Research Methods. Elsevier, New York, 255-270pp. Erftemeijer, P.L.A. and G.R. Allen. 1993. Fish fauna of seagrass beds in South Sulawesi, Indonesia. Rec. West. Aust. Mus., 16(2):269-277. Gilanders, B.M. 2006. Seagrasses, Fish, and Fisheries. In: Larkum, A.W.D., Orth, R.J., Duarte, C.M. (Eds.), Seagrasses: Biology, Ecology, and Conservation. Springer, The Netherland, 503-536pp. Hemminga, M.A. and C.M. Duarte. 2000. Seagrass Ecology. Cambridge University Press, Cambridge, UK. Hind, J.A. 1982. Stability and Trim of Fishing Vessels and Other Small Ships. Second Edition. Fishing News Books Ltd. Farnham, Surrey, England. International Maritime Organization (IMO). 1983. International Confrence on Safety Fishing Vessels 1977. IMO. London. Iskandar, B.H. 1997. Studi tentang Desain Kapal Kayu Mina Jaya BPPT 01. Tesis pada Program Pascasarjana IPB. Bogor. Kok, H.G.M, E.G.V. Lonkhyusen, and F.A.C. Nierich. 1983. Bangunan Kapal. Zundort. Netherland. Kikuchi, T., J.M. Peres. 1977. Consumer Ecology of Seagrass Beds. In: McRoy, C.P., Helffrich, C. (Eds.), Seagrass Ecosystems: A Scientific Perspective. Marcel Dekker, Inc., New York, 147-193pp. Kuiter, R.H. and T. Tonozuka. 1992. Tropical Reef of The Western Pacific, Indonesia and Adjacent http://www.itk.fpik.ipb.ac.id/ej_itkt22 Rappe Waters. PT Gramedia Pustaka Major, Jakarta. Odum, E.P. 1983. Basic Ecology. Saunders College Publishing, New York. Radjab, W. A. S. Dody, dan F.D. Hukom. 1992. Komunitas Ikan di Padang Lamun Perairan Passo Teluk Baguala. Balai penelitian dan Pengembangan Sumberdaya Laut, P2O-LIPI, Ambon. Rani, C., Budimawan, dan Rohani. 2010. Kajian keberhasilan ekologi dari penciptaan habitat dengan lamun buatan: penilaian terhadap komunitas ikan. Ilmu Kelautan. Indonesian Journal of Marine Sciences, 2(Edisi Khusus):244-255. Romimohtarto, K. dan S. Juwana. 2001. Biologi Laut. Ilmu Pengetahuan Tentang Biota Laut. Penerbit Djambatan. Jakarta. Sidik, B.J., S.O. Bandeira, and N.A. Milchakova. 2001. Methods to Measure Macroalgal Biomass and Abundance in Seagrass Meadows. In: Short, F.T., Coles, R.G. and Short, C.A. (Eds.), Global Seagrass Research Methods. Elsevier, New York, 223-235pp. Supriadi, Y.A. La Nafie, dan A.I. Burhanuddin. 2004. Inventarisasi jenis, kelimpahan, dan biomassa ikan di padang lamun Pulau Barrang Lompo Makassar. Torani, 14(5): 288-295. Taylor, L.G. 1977. The Principles of Ship Stability. Brown, Son & Publisher, Ltd., Nautical Publisher, 52 Darnley Street. Glasgow. Unsworth, R.K.F., E. Wylie, D.J. Smith, and J.J. Bell. 2007. Diel trophic structuring of seagrass bed fish assemblages in the Wakatobi Marine National Park, Indonesia. Estuarine, Coastal and Shelf Science, 72:81-88. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 73 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 74-82, Desember 2010 DISTRIBUSI FORAMINIFERA BENTIK RESEN DI LAUT ARAFURA THE DISTRIBUTION OF RECENT BENTHIC FORAMINIFERA IN THE ARAFURA SEA Suhartati M. Natsir dan Rubiman Pusat Penelitian Oseanografi-LIPI Email: [email protected] ABSTRACT Arafura Sea consists of shallow waters and located in the Southern of Papua to the north coast of Australia. The waters is vegetated by shallow-water ecosytems such as mangrove, seagrass bed, and coral reefs. The Arafura continental shelf is predominated by sediment from late Paleozoic, Mesozoic to Cenozoic and underlain by granitic basement. Foraminifera is a single cell microorgainsm, has pseudopodia with high level of diversity. Foraminifera dwells in every level of sea depth, from estuary to the deep sea. However, a certain species commonly dwells in the specific profundity. The aim of the study was to recognize the distribution of benthic foraminifera in the waters of Arafura Sea and it relation with the environmental characteristics. As many as 11 sediment samples was collected in May 2010 from the water of Arafura Sea using a box core with capcity of 0,3 m3. Laboratory analyses on the colleted samples were performed to determine the type of sediments and identify the benthic foraminifera, and to determine the abundance of each samples. The number of species found from the collected sediments were 37 species consisting of 29 genera of which most of them were member of Suborder Rotaliina and many of them belong to Suborder Miliolina and Textulariina. The most common species of the sampling sites were Ammonia beccarii and Pseudorotalia schroeteriana. The Arafura Sea commonly recognized as shallow waters, open seas, with current speed of midium to high. The predominant sediment type of the waters is sandy mud and little of clay. Keywords: distribution, benthic foraminifera, sediment and Arafura ABSTRAK Laut Arafura merupakan perairan dangkal yang terletak di wilayah Papua bagian Selatan sampai bagian utara pantai Australia. Ekosistem yang terdapat pada perairan tersebut merupakan ekosistem penciri perairan dangkal seperti hutan mangrove, padang lamun dan terumbu karang. Sedimen yang mendominasi landas kontinen perairan Arafura berasal dari masa Paleozoikum akhir, Mesozoikum sampai Kenozoikum yang dilandasi oleh lapisan granit pada bagian bawah. Foraminifera merupakan mikroorganisme bersel tunggal dan berkaki semu yang mempunyai keragaman sangat tinggi. Habitat foraminifera terdiri dari semua kedalaman laut dari tepi pantai sampai pada laut dalam. Secara umum, suatu spesies bentik hidup pada kedalaman tertentu. Penelitian ini dilakukan untuk mengetahui distribusi foraminifera bentik yang terdapat pada sedimen di perairan Laut Arafura dan kaitannya dengan karakteristik perairan tersebut. Pengambilan sampel dilakukan pada bulan Mei 2010 di Peraiaran Laut Arafura. Sebanyak 11 sampel sedimen diambil dari dasar perairan menggunakan box core. Kemudian sampel yang diperoleh dianalisis jenis sedimennya dan kandungan foraminifera bentik didalamnya. Jumlah spesies yang ditemukan mencapai 37 spesies yang termasuk dalam 29 genus yang sebagian besar merupakan anggota dari subordo Rotaliina dan beberapa spesies merupakan anggota Miliolina dan Textulariina. Spesies yang ditemukan merata hampir di semua stasiun adalah Ammonia beccarii dan Pseudorotalia schroeteriana. Karakeristik sebagian besar perairan Laut Arafura merupakan perairan dangkal, terbuka dengan tingkat energi arus menengah sampai kuat. Jenis sedimen yang mendominasi perairan Laut Arafura adalah Lumpur pasiran dengan sedikit lempung. Kata Kunci: distribusi, foraminifera bentik, Sedimen, Arafura 74 ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Natsir dan Rubiman I. PENDAHULUAN Laut Arafura merupakan perairan yang meliputi landas kontinen Arafura – Sahul dan terletak di wilayah Papua bagian Selatan sampai perbatasan Benua Australia. Batas bagian Utara perairan tersebut merupakan Laut Seram dan Pulau Irian Jaya (Papua), sedangkan Pantai Utara Australia dari Semenanjung York sampai Semenanjung Don merupakan batas di bagian Selatan. Di bagian Barat, perairan tersebut dibatasi oleh Laut Banda dan Laut Timor yang melewati Kepulauan Aru dan Tanimbar. Sedangkan di bagian Timur terdapat Pulau Dolak dan Semenajung Don yang membatasi perairan tersebut. Brdasarkan tingkat kedalamannya, Laut Arafura termasuk perairan dangkal dengan kisaran kedalaman antara 30-90 m. Ekosistem yang terdapat pada perairan tersebut merupakan ekosistem penciri perairan dangkal seperti hutan mangrove, padang lamun dan terumbu karang (Wagey dan Arifin, 2008). Menurut Katili (1986), sedimen yang mendominasi landas kontinen perairan Arafura berasal dari masa Paleozoikum akhir, Mesozoikum sampai Kenozoikum yang dilandasi oleh lapisan granit pada bagian bawah. Foraminifera merupakan mikroorganisme bersel tunggal dan berkaki semu yang mempunyai keragaman sangat tinggi dan menempati hampir 2,5% dari seluruh hewan yang dikenal sejak zaman kambrium hingga resen. Sebanyak 38.000 spesies berupa fosil dan 10.000 – 12.000 spesies foraminifera resen ditemukan di seluruh lauatan (Boltovskoy and Wright, 1976). Menurut Murray (1973), distribusi dan kelimpahan spesies mendapat perhatian yang cukup besar, baik spesies yang masih hidup maupun yang sudah mati. Foraminifera merupakan kelompok hewan yang sebagian besar hidup di laut. Program pemantauan lingkungan perairan dapat dilakukan berdasarkan distribusi foraminifera karena beberapa keunggulannya antara lain ukurannya yang relatif kecil, hidup pada lingkungan tertentu, jumlahnya melimpah, mudah dikoleksi, ekonomis dan secara signifikan dapat diolah secara statistik. Habitat foraminifera terdiri dari semua kedalaman laut dari tepi pantai sampai pada laut dalam. Secara umum, suatu spesies bentik hidup pada kedalaman tertentu. Kedalaman merupakan faktor ekologi yang mempengaruhi distribusinya (Boltovskoy and Wright, 1976). Tujuan dari Penelitian ini adalah untuk mengetahui distribusi foraminifera bentik yang terdapat pada sedimen di perairan Laut Arafura dan kaitannya dengan karakteristik perairan tersebut. II. METODE PENELITIAN Secara umum, metode yang digunakan dalam penelitian dilapangan adalah metode survey, sedangkan observasi dan analisis dilakukan di dalam laboratorium. Pengambilan sampel dilakukan pada bulan Mei 2010 di Laut Arafura dari bagian tenggara Kepulauan Tanimbar ke arah bagian selatan dan timur Kepulauan Aru sampai sekitar Pulau Dolak dan Pulau Irian Jaya (Papua) (Gambar 1). Sedimen dasar laut diambil dengan menggunakan box core yang berkapasitas 0,3 m3 untuk memperoleh sampel foraminifera bentik dari 11 lokasi yang telah ditentukan. Sampel yang diperoleh dimasukkan ke dalam kantong plastik yang telah diberi label untuk dianalisa lebih lanjut di laboratorium. Proses preparasi, observasi dan analisis terhadap sampel dilakukan di laboratorium Geologi Laut, Pusat Penelitian Oseanografi LIPI, Jakarta. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 75 Distribusi Foraminifera Bentik Resen di Laut Arafura Gambar 1. Lokasi pengambilan sampel di perairan Laut Arafura Sampel yang diperoleh merupakan material dari dasar laut secara keseluruhan yang meliputi material sedimen, serasah dan organisme termasuk foraminifera bentik. Tahap preparasi diperlukan untuk memisahkan foraminifera bentik yang terdapat pada sampel tersebut dari bahan-bahan dan organisme lain sehingga dapat diidentifikasi dengan mudah. Preparasi sampel dilakukan dengan beberapa tahap, antara lain pencucian sampel, picking, deskripsi dan identifikasi serta sticking dan dokumentasi. Pencucian sampel dilakukan dengan menggunakan air mengalir diatas saringan dengan diameter berturut-turut 1.0, 0.5, 0.250, 0.125, 0.063 mm. Setelah pencucian, sampel tersebut dikeringkan menggunakan oven pada suhu 30°C sampai kering (selama ± 30 menit). Sampel yang telah kering dimasukkan ke dalam kantong plastik yang telah diberi label untuk analisis lebih lanjut. Setelah 76 pencucian dan pengeringan, saringan harus direndam dalam larutan methiline blue untuk mencegah kontaminasi oleh sampel berikutnya dan dicuci. Tahap selanjutnya adalah picking yang dilakukan dengan menyebarkan sampel yang telah dicuci pada extraction tray dibawah mikroskop secara merata. Foraminifera yang terdapat dalam sampel tersebut diambil dan disimpan pada foraminiferal slide. Kemudian dilakukan proses deskripsi dan identifikasi terhadap spesimen yang didapatkan. Spesimen yang telah dipisahkan diklasifikasikan berdasarkan morfologinya seperti bentuk cangkang, bentuk kamar, formasi kamar, jumlah kamar, ornamentasi cangkang, kemiringan apertura, posisi apertura dan kamar tambahan. Sedangkan proses identifikasi dilakukan berdasarkan berbagai referensi tentang foraminifera bentik. Tahap selanjutnya merupakan kajian sistemik dan analisis kuantitatif http://www.itk.fpik.ipb.ac.id/ej_itkt22 Natsir dan Rubiman untuk mendapatkan data kelimpahan. Proses sticking dan dokumentasi dilakukan dengan meletakkan spesimen yang terpilih pada foraminiferal slide dengan posisi tampak apertura, tampak dorsal, tampak ventral dan tampak samping yang kemudian didokumentasikan dibawah mikroskop. Pengelompokan kelimpahan foraminifera bentik yang ditemukan berdasarkan jumlah spesimen yang ditemukan. kelimpahan foraminifera bentik dikelompokkan kedalam 3 kategori yaitu tinggi (melimpah), sedang dan rendah (jarang). Spesies yang tergolong dalam kelimpahan tinggi merupakan spesies yang ditemukan sebanyak lebih dari 50 spesimen, sedangkan kelimpahan sedang dan rendah masing-masing diwakili oleh jumlah spsies yang ditemukan sebanyak 11 – 50 spesimen dan kurang dari 11 spesimen. Penentuan jenis sedimen dari sampel yang diambil dilakukan dengan analisis granulometri menggunakan ayakan berukuran 0,063 – 4 mm. Pengelompokan butir sedimen dilakukan berdasarkan skala Wenworth (1922) dan penamaannya berdasarkan klasifikasi Shepard (1960). III. HASIL DAN PEMBAHASAN Laut Arafura terletak di wilayah Papua bagian Selatan sampai perbatasan Benua Australia. Menurut Wagey dan Arifin (2008), perairan Laut Arafura merupakan perairan dangkal dengan kisaran kedalaman antara 30-90 m dengan Ekosistem yang terdapat pada perairan tersebut merupakan ekosistem penciri perairan dangkal seperti hutan mangrove, padang lamun dan terumbu karang. Namun, pada penelitian ini ditemukan bahwa terdapat perairan yang mempunyai kedalaman lebih dari 300 m. Stasiun 13 yang terletak di bagian Selatan Kepulauan Tanimbar tercatat mempunyai kedalaman yang mencapai 341 m. Lokasi tersebut diduga merupakan titik pertemuan antara Busur Banda dan lempeng Benua Australia seperti yang dinyatakan oleh Katili (1986) bahwa terdapat lengkungan kebawah pada sedimen di perairan Arafura yang berbatasan dengan Busur Banda. Pola tektonik dari deformasi tersebut terjadi karena dorongan Busur Banda ke arah Benua Australia dan semakin meningkat ke arah Utara. Sedangkan kedalaman di stasiun lainnya tercatat tidak lebih dari 60 m dan perairan paling dangkal ditemukan di dekat Pulau Dolak (stasiun 14). Secara keseluruhan, hasil analisis terhadap sampel sedimen yang diperoleh dari 10 lokasi di perairan Laut Arafura diperoleh foraminifera bentik resen sebanyak 1593 individu. Jumlah tersebut terdiri dari 37 spesies yang termasuk dalam 29 genus (Tabel 1). Sebagian besar spesies yang ditemukan merupakan anggota dari subordo Rotaliina, namun juga ditemukan beberapa spesies yang merupakan anggota Milioliina dan Textulariina. Kelimpahan dan jenis foraminifera bentik yang ditemukan pada masing-masing stasiun berbeda-beda seiring dengan komposisi atau jenis sedimennya. Setiap stasiun mempunyai komposisi kelimpahan foraminifera bentik yang berbeda. Jumlah foraminifera terbanyak diperoleh dari stasiun 22 dengan kedalaman 38 m yang terletak di sebelah tenggara Kepulauan Aru. Sedimen yang mendominasi stasiun tersebut adalah jenis lumpur pasiran dan sedikit lempung (Tabel 2). Spesies yang ditemukan melimpah pada stasiun tersebut adalah Ammonia beccarii dan Pseudorotalia schroeteriana yang masing-masing mencapai 111 dan 64 individu (Tabel 1). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 77 Distribusi Foraminifera Bentik Resen di Laut Arafura Tabel 1. Jumlah foraminifera bentik yang ditemukan pada sampel yang berasal dari perairan Laut Arafura Sampel Foraminifera Benthic 13 14 15 16 17 18 19 20 21 22 23 Ammonia beccarii (Linnaeus) - 104 21 46 121 69 14 116 131 111 14 Amphistegina lessonii - - - - - - - - - - 1 Anomalina rostrata (Bradyi) 21 - - - - - - - - - - Asterorotalia trispinosa - - - - - - - - - 1 - Astocolus reniformis (d'Orbigny) 6 - - - - - - - - - - Bolivina earlandi (Parr) 82 - - - 2 - - - - - - Bolivina spathulata (Williamson) 26 16 - - 41 4 - - - - 4 Bolivina subspinecens (Cushman) 12 - - - - - - - - - - Cancris oblongus (Cushman) 40 - 48 - - - - - - 18 - Cibicides berthelotianus (d'Orbigny) - - - - - - 2 - - - - Cibicides molis - - - - - - - - - 14 - Discorbinella biconcavus (Parker & Jones) - - - - - - - - - - 2 Elphidium craticulatum - - - - - - - 6 9 41 - Elphidium crispum - - 7 - - 8 4 - - 29 - Eponides berthelotianus (d'Orbigny) - 12 - - - - - - - - - Fissurina exsculpra (Brady) - - - - 1 - - - - - - Guttulina dawsoni (Chusman and Ozawa) - - - - - - 2 - - - - Gyroidina neosoldanii - - - - - - 2 2 - - 2 Hoglundina elegans (d'Orbigny) - - - - 6 - 2 4 - - - Lagena gracillisima (Sguenza) 1 - - - - - - - - - - Nonion sp. 8 12 - - - - 24 2 - - 2 Oolina apiculata (Reuss) - - - - - 2 - - - - - Operculina ammonoides - - - - - - - 12 12 - 1 Planispinoides bucculantus (Brady) - - - - - - - - 6 - - Planorbulina sp. (d'Orbigny) - - - - - - - - - - 3 Pseudopolymorphina ligua (Rosmer) - - - - - - 2 - - - - Pseudorotalia schroeteriana - 40 - 28 30 24 3 21 6 64 2 Quinqueloculina cultrate - - - - 2 - - - - - - Quinqueloculina granulocostata - - - - - 4 - - - 3 - Quinqueloculina parkery - 2 - - 1 3 - - - 8 1 Quinqueloculina seminulum - - - - - 2 - - - 8 2 Quinqueloculina sp. Rosalina sp. Spiroloculina communis - - - - 1 - 2 - - - 4 - 2 3 Textularia pseudogramen - - - - - - - - - - 2 Triloculina tricarinata - - - - - - - - - - 8 Young miliolidae 4 - - - - 2 - - - - - 78 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Natsir dan Rubiman Tabel 2. Kedalaman dan jenis sedimen pada masing-masing stasiun pengambilan sampel di perairan Laut Arafura Stasiun Kedalaman (m) Jenis sedimen 13 341 Lempung 14 19 Lempung 15 35 lumpur pasiran - pasir sedang 16 29 lanau – lempung 17 35 Lumpur - pasir sedang – lempung 18 38 Lumpur pasiran – lempung 19 48 Lumpur pasiran – lempung 20 60 Lumpur pasiran 21 35 Lumpur pasiran – lempung 22 38 Lumpur pasiran – Lempung 23 59 Pasir (sedang-kasar) lumpuran Boltovskoy and Wright (1976) menyatakan bahwa Asterorotalia trispinosa dan Ammonia beccarii banyak dijumpai pada sedimen pasir dan lumpur pasiran. Namun A. Trispinosa hanya ditemukan dalam jumlah yang sangat sedikit. Spesies yang ditemukan dengan tingkat kelimpahan sedang pada stasiun tersebut adalah Cancris oblongus, Cibicides molis dan dua spesies dari genus Elphidium. Spesies yang terdapat melimpah dan sedang tersebut merupakan anggota dari Subordo Rotaliina. Selain itu, pada stasiun 22 juga ditemukan beberapa spesies yang termasuk dalam Subordo Miliolina, namun dalam jumlah yang sedikit atau termasuk dalam kelimpahan rendah. Spesies-spesies tersebut diwakili oleh merupakan anggota dari genus Quinqueloculina yang diwakili oleh Quinqueloculina sp., Q. granulocostata, Q. parkery dan Q. seminulum. Spesiesspesies yang bercangkang hialin tersebut masing-masing ditemukan tidak lebih dari 10 individu. Spesies yang bersimbiosis dengan terumbu karang, berdasarkan klasifikasi yang dilakukan oleh Hallock et al. (2003) hanya ditemukan pada stasiun 23 dengan jumlah yang sangat sedikit. Hal ini dapat mengindikasikan bahwa perairan tersebut bukan termasuk lingkungan yang kondusif untuk pertumbuhan terumbu karang. Spesies tersebut adalah Amphistegina lessonii yang ditemukan dengan kondisi cangkang yang sudah rusak. Demikian pula dengan beberapa spesies yang ditemukan pada stasiun 17 dan 19 juga ditemukan dengan kondisi cangkang yang rusak. Hal tersebut dapat dimungkinkan akibat hempasan arus sehingga dapat menghancurkan cangkang foraminifera bentik yang terdapat di perairan tersebut. Beberapa spesies yang ditemukan di lokasi ini merupakan penciri perairan dangkal dan terbuka dengan kecepatan arus menengah sampai tinggi. Menurut Gustiantini dan Usman (2008), beberapa spesies dari genus Elphidium merupakan penciri perairan dangkal dengan energi arus yang relatif tinggi. Sedangkan spesies dari genus Quinqueloculina merupakan penghuni lingkungan perairan terbuka dengan kecepatan arus sedang sampai tinggi, serta sedimen lumpur dan pasir (Boltovskoy and Wright, 1976; Yassini and Jones, 1995; dan Rositasari dan Rahayuningsih, 2000). Suhartati (1994 dan 2010) juga menemukan Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 79 Distribusi Foraminifera Bentik Resen di Laut Arafura Quinqueloculina melimpah di Pulau Pari dan Pulau Belanda, Kepulauan Seribu pada kedalaman 26-32 m, sedangkan Barker (1960) menemukannya di bagian selatan Papua pada kedalaman 37 m. Graham dan Milante (1959) menemukan spesies-spesies tersebut sangat melimpah pada beberapa stasiun di Teluk Puerto Galera, Philipina dan termasuk spesies kosmopolitan. Spesies yang ditemukan hampir di seluruh stasiun adalah Ammonia beccarii dan Pseudorotalia schroeteriana. A. beccarii ditemukan sangat melimpah pada semua stasiun kecuali stasiun 13 dengan kedalaman yang mencapai 341 m. Menurut Hallock et. al. (2003), A. beccarii tergolong dalam spesies yang oportunis sehingga dapat ditemukan di berbagai lokasi yang berbeda. Walaupun demikian, terdapat spesies oportunis lain yang ditemukan dalam jumlah melimpah dan dominan pada stasiun 13, yaitu dari genus Bolivina. Genus tersebut didominasi oleh spesies Bolivina erlandi yang ditemukan mencapai 82 individu, sedangkan B. spathulata dan B. subspinecens masing hanya mencapai 26 dan 12 individu. Sebagai spesies penciri perairan dangkal, P. schroeteriana juga ditemukan hampir di semua stasiun kecuali stasiun 13 dan 15. Spesies tersebut ditemukan dengan kelimpahan rendah sampai tinggi. Menurut Biswas (1976), P. schroeteriana merupakan penciri perairan dangkal, terbuka dengan tingkat energi arus menengah dengan sedimen pasir halus. Oleh karena itu karakeristik sebagian besar perairan Laut Arafura merupakan perairan dangkal, terbuka dengan tingkat energi arus menengah sampai kuat karena juga ditemukan Elphidium sebagai penciri perairan berarus kuat. Selain itu, spesies dari genus Elphidium juga termasuk dalam genus oportunis sesuai dengan pernyataan 80 Hallock et al. (2003), terbukti dengan ditemukannya spesies tersebut pada 6 stasiun dari 11 stasiun yang diteliti. Namun, genus yang diwakili oleh E. craticulatum dan E. crispum tersebut hanya ditemukan dengan kelimpahan rendah sampai sedang (tidak lebih dari 50 individu). Spesies-spesies tersebut ditemukan pada stasiun yang memiliki kisaran kedalaman antara 35-60 m. Hal ini sesuai dengan hasil peneletian yang dilakukan oleh Murray (1973) dan Boltovskoy dan Wright (1976) yang menyatakan bahwa E. craticulatum dan E. crispum memiliki penyebaran yang luas dari daerah pantai hingga neritik tengah. Menurut Katili (1986), sedimen yang mendominasi landas kontinen perairan Arafura berasal dari masa Paleozoikum akhir, Mesozoikum sampai Kenozoikum yang dilandasi oleh lapisan granit pada bagian bawahnya. Hasil analisis sedimen yang diperoleh pada lokasi penelitian menunjukkan bahwa sebagian besar sedimen di perairan Laut Arafura adalah lumpur pasiran. Sedimen berupa lempung ditemukan disekitar Kepulauan Tanimbar dengan kedalaman 341 m (stasiun 13) dan di sekitar Pulau Dolak (stasiun 14). Sedangkan sedimen pasir sedang sampai kasar yang bercampur dengan fragmen karang dan moluska ditemukan pada stasiun 23 yang terletak di bagian selatan Kepulauan Aru (Tabel 2 dan 3). Jenis spesies yang ditemukan pada stasiun tersebut sebanyak 13 spesies. Jumlah tersebut relatif lebih banyak dibandingkan dengan stasiun lainnya, namun kelimpahan masing-masing spesies tergolong sangat rendah. Kelimpahan tertinggi hanya mencapai 14 individu, yaitu pada spesies Ammonia beccarii. Secara keseluruhan jumlah foraminifera bentik yang ditemukan di stasiun 23 hanya mencapai 48 individu. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Natsir dan Rubiman Selain foraminifera bentik, juga ditemukan foraminifera planktonik yang menyebar hampir di semua stasiun kecuali stasiun 16, 17 dan 22. Begitu pula dengan fragmen moluska yang juga terdapat di hampir semua stasiun (Tabel 3). Hal ini diduga karena karakteristik perairan Laut Rafura yang terbuka dengan arus yang relatif kuat memungkinkan distribusi foraminifera planktonik dan fragmen moluska tersebut ke beberapa staiun disekitarnya, termasuk perairan dalam (stasiun 23). IV. KESIMPULAN Jumlah spesies yang ditemukan di perairan Laut Arafura dari sekitar Kepulauan Tanimbar, Kepulauan Aru hingga Pulau Dolak adalah 37 spesies yang termasuk dalam 29 genus. Sebagian besar spesies yang ditemukan merupakan anggota dari subordo Rotaliina dan beberapa spesies yang merupakan anggota Milioliina dan Textulariina. Spesies yang ditemukan merata hampir di semua stasiun adalah Ammonia beccarii dan Pseudorotalia schroeteriana. Berdasarkan distribusi foraminifera bentik yang ditemukan, karakeristik sebagian besar perairan Laut Arafura merupakan perairan dangkal, terbuka dengan tingkat energi arus menengah sampai kuat. Selain P. schroeteriana, juga ditemukan spesies penciri lainnya seperti dari genus Elphidium dan Quinqueloculina. Selain itu, pada perairan terbuka tersebut juga ditemukan foraminifera planktonik yang dtersebar merata hampir di setiap stasiun. Jenis sedimen yang mendominasi perairan Laut Arafura adalah Lumpur pasiran dengan sedikit lempung. Jumlah individu terbanyak diperoleh dari stasiun dengan sedimen lumpur pasiran, sedangkan jumlah spesies terbanyak diperoleh dari sedimen pasir lumpuran dengan butiran pasir sedang sampai kasar. Tabel 3. Organisme selain foraminifera bentik yang ditemukan dari sampel yang berasal dari perairan Laut Arafura Keterangan Stasiun 13 14 15 16 17 18 19 20 21 22 23 Foraminifera planktonik - - - Moluska - - - - - - Bryozoa - - - - - - Gastropoda - - - - - - - - - - Ostracoda - - - - - - - - Fragmen karang - - - - - - - - Fragmen moluska - - Keterangan: = terdapat dalam jumlah banyak; = terdapat dalam jumlah sedang; = terdapat dalam jumlah sedikit; − = tidak ada Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 81 Distribusi Foraminifera Bentik Resen di Laut Arafura DAFTAR PUSTAKA Barker, R.W. 1960. Taxonomic Notes. Society of Economic Paleontologist and Mineralogist. Special Publication No. 9. Tulsa. Oklahoma, USA. 238 pp. Boltovskoy, E. and R. Wright. 1976. Recent Foraminifera. Dr. W. June, B. V. Publisher, The Haque, Netherland. Graham, J.J. and Militante. 1959. Recent Foraminifera from The Puerto Galera Area Northern Mindoro, Philippines. Stanford University, California. Hallock, P., B.H. Lidz, E.M. CockeyBurkhard, and K.B. Donnelly. 2003. Foraminifera as bioindicators in coral reef assessment and monitoring: the FORAM Index. Environmental Monitoring and Assessment, 81(13):221-238. Katili, J.A. 1986. Geology and hydrocarbon potential of the Arafura Sea. In: Future Petroleum Provinces of the World. AAPG Memoir 40, M.T. Halbouty (editor) 487-501. Murray, J. W. 1973. Distribution and Ecology of Living Foraminifera. The John Hopkins Press. Baltimore. Rositasari R. dan S. K. Rahayuningsih. 2000. Foraminifera Bentik: Dalam Foraminifera sebagai bioindikator pencemaran, hasil studi di perairan estuarin Sungai Dadap, Tangerang. Pusat Penelitian dan Pengembangan Oseanografi. Lembaga Ilmu Pengetahuan Indonesia: 3-26. Shepard, F. P. 1954, Nomenclature based on sand-silt-clay ratios: Journal of Sedimentari Petrology, 24:151-158. 82 Suhartati, M.N. 1994. Benthic Foraminifera In The Seagrass Beds of Pari Island, Seribu Islands, Jakarta. Proceedings. Third ASEANAustralia Symposium on Living Coastal Resources. Volume 2: Research Papers. Chulalongkorn University Bangkok, Thailand. 323p. ________. 2010. Sebaran Foraminifera Bentik di Pulau Belanda, Kepulauan Seribu pada Musim Barat. Ilmu Kelautan, Edisi khusus, 2:381–387. Wagey, T., Arifin, Z. 2008. Marine Biodiversity Review of The Arafura and Timor Seas. Ministry of Marine Affairs and Fisheries, Indonesian Institute of Sciences, United Nation Development Program, and Cencus of Marine Life. Jakarta. 136 pp. Wentworth, C. K. 1922, A scale of grade and class term for clastic sediment. Jour. Geol. 30:337-392 Yassini, I. and B.G. Jones. 1995. Foraminiferida and Ostracoda from estuarne and shelf environments on The South Eastern Coast of Australia. University press., Wollonggong. 270 pp. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 83-91, Desember 2010 PERENCANAAN WAKTU TETAS TELUR IKAN KERAPU DENGAN PENGGUNAAN SUHU INKUBASI YANG BERBEDA PLANNING ON HATCHING TIME OF GROUPER EGGS THROUGH DIFFERENT INCUBATION TEMPERATURES Regina Melianawati, Philip Teguh Imanto, dan Made Suastika Balai Besar Riset Perikanan Budidaya Laut Bali; email: [email protected] ABSTRACT Groupers were known as a high economically marine commodity and in order to support groupers production, the seed availability was the most important. Eggs are still as limited factor in hatchery production, for this reason the success of eggs transportation is one as base of successful production of seed. Planning on hatching time of eggs through different incubation temperature was an option to solve that problem. This experiment was aimed to find out the optimum temperature for groupers eggs and the minimum temperature to arrange incubation time and to plan the hatching time. Fertilized eggs were incubated into three beaker glasses of 1 liter in volume with the density of ± 250 eggs/liter. The incubation was done under laboratory condition at controlled temperature, i.e. (A) 21-22 ºC, (B) 24-25 ºC, (C) 27-28 ºC and (D) 30-31 ºC. The eggs that used were including orange spotted grouper (Epinephelus coiodes), brown marbled grouper (E. microdon), tiger grouper (E. fuscoguttatus) and humpback grouper (Cromileptes altivelis). Investigated variables were embryonic development pattern, incubation time and hatching rate. The result showed that the eggs incubated in temperature range of 24-31°C had the normal sequence of embryonic development pattern, but in temperature of 21-22°C performed irregular sequence and the embryonic development stopped at blastula or gastrula stage or even the eggs could still develop but the body of hatched larvae were abnormal. In lower temperature incubation, the incubation time was longer and the hatching rate of eggs was lower than those in higher temperature. Therefore the optimum temperature for incubation of orange spotted grouper, marbled grouper, tiger grouper and humpback grouper eggs ranged between 24-31 ºC, while the lowest possible temperature was 24 ºC. Keywords: incubation temperature, embryonic development pattern, grouper eggs, hatching rate ABSTRAK Ikan kerapu merupakan komoditas bernilai ekonomis tinggi dan benihnya sangat diperlukan Pasok telur masih menjadi faktor pembatas dalam produksi benih, dan transportasi telur menjadi salah satu kunci keberhasilan produksinya. Transportasi telur jarak jauh dapat dilakukan dengan perencanaan waktu tetas melalui manipulasi suhu media penetasan. Tujuan penelitian untuk mengetahui kisaran suhu optimum dan batas toleransi suhu terendah untuk mengatur masa inkubasi dan perencanaan waktu tetas telur ikan kerapu. Penelitian menggunakan telur fertil yang diinkubasikan dalam 3 buah beaker glass volume 1 liter dengan kepadatan ± 250 butir/liter. Suhu inkubasi yang diujikan adalah (A) 21-22 ºC ; (B) 24-25 ºC ; (C) 27-28 ºC dan (D) 30-31 ºC. Jenis telur ikan kerapu yang digunakan dalam penelitian ini adalah telur kerapu lumpur (Epinephelus coiodes), kerapu batik (E. microdon), kerapu macan (E. fuscoguttatus) dan kerapu bebek (Cromileptes altivelis). Parameter pengamatan meliputi pola perkembangan embrio, masa inkubasi dan tingkat penetasan. Hasil penelitian menunjukkan bahwa telur yang diinkubasikan pada suhu 24-31°C menghasilkan pola perkembangan embrio yang teratur, sedang pada suhu 21-22°C perkembangan embrio terhenti pada stadia blastula atau gastrula atau terus berkembang tetapi menghasilkan larva yang cacat. Pada suhu rendah, masa inkubasi berlangsung lebih lama dan tingkat penetasan telurnya lebih rendah dibandingkan pada suhu yang lebih tinggi. Kisaran suhu optimum bagi penetasan telur kerapu lumpur, kerapu batik, kerapu macan dan kerapu bebek adalah 24-31oC, dengan batas toleransi suhu terendah dalam kaitannya untuk mengatur masa inkubasi dan perencanaan waktu tetas adalah 24oC. Kata Kunci: suhu inkubasi, pola perkembangan embrio, telur ikan kerapu, tingkat penetasan ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 83 Perencanaan Waktu Tetas Telur Ikan Kerapu dengan… I. PENDAHULUAN Ikan kerapu merupakan komoditas perikanan laut yang bernilai ekonomis tinggi. Permintaan terhadap jenis ikan ini tidak saja berasal dari dalam negeri tetapi juga sebagai komoditas ekspor yang banyak diminati, baik di Asia seperti Taiwan, Jepang, Singapura dan Hongkong, maupun di Eropa, Australia dan Amerika (Andamari et al., 2005). Beberapa jenis ikan kerapu yang bernilai ekonomis tinggi diantaranya adalah ikan kerapu lumpur Epinephelus coiodes (Lau dan Jones, 1999), kerapu batik E. microdon (Slamet, et al., 1997), kerapu macan E. fuscoguttatus (Kohno et al., 1990; Sudjiharno et al., 2001) dan kerapu bebek Cromileptes altivelis (Putro et al., 1999; Sudaryanto et al., 1999). Selama ini pemenuhan kebutuhan pasar mayoritas diperoleh dari hasil penangkapan di alam. Untuk memenuhi permintaan pasar yang semakin tinggi dan sekaligus menjaga kelestarian populasi ikan kerapu di alam, maka kegiatan budidaya mutlak diperlukan. Hal ini menjadikan produksi benih yang berasal dari hatchery (panti benih) menjadi sangat penting peranannya. Dalam usaha pembenihan, pasok telur menjadi faktor pembatas produksi karena tidak semua panti benih mampu mencukupi sendiri kebutuhan telurnya. Penanganan telur, termasuk transportasinya hingga sampai di lokasi pembenihan merupakan tahap awal yang menentukan keberhasilan usaha tersebut. Hambatan yang sering dialami pada transportasi telur ini adalah masa inkubasinya yang singkat sehingga sering terjadi telur telah menetas sebelum tiba di lokasi tujuan. Apabila hal ini terjadi akan menyebabkan kualitas air media menjadi buruk dan mengakibatkan kematian pada telur atau larva yang baru menetas dalam proses transportasi (Slamet, 1993). Suhu merupakan salah satu faktor yang penting 84 dalam transportasi karena berpengaruh terhadap nilai metabolisme mahkluk hidup dan perkembangan embrio (Mulyanto, 1990 dalam Setiadharma et al., 1997). Penelitian ini bertujuan mengetahui kisaran suhu optimum dan batas toleransi suhu terendah dalam kaitannya untuk mengatur masa inkubasi dan perencanaan waktu tetas telur ikan kerapu. II. METODE PENELITIAN Penelitian dilaksanakan di laboratorium Balai Besar Riset Perikanan Budidaya Laut Bali. Telur yang digunakan untuk pengamatan adalah telur yang fertil dan merupakan hasil pemijahan induk secara alami pada tangki beton. Telur fertil yang telah diseleksi selanjutnya diinkubasi dalam wadah beaker glass volume 1 liter, masingmasing beaker glass diisi dengan 250 butir. Setiap ulangan (3 beaker glass) ditempatkan dalam sebuah wadah plastik berbentuk empat persegi panjang dengan volume ± 46 liter dan semua wadah tersebut kemudian ditempatkan dalam sebuah cooler waterbath yang dilengkapi dengan alat pengatur suhu. Suhu pada waterbath diatur pada 21-22°C untuk menyesuaikan dengan suhu terendah yang diujikan dalam penelitian ini. Untuk mengatur suhu dalam masingmasing wadah tersebut digunakan heater yang suhunya disesuaikan dengan perlakuan yang diujikan. Penelitian dilaksanakan dengan 4 perlakuan tingkat suhu inkubasi yang berbeda yaitu (A) 21-22 ºC, (B) 24-25 ºC, (C) 27-28 ºC dan (D) 30-31 ºC. Masingmasing perlakuan terdiri dari 3 ulangan. Jenis telur ikan kerapu yang digunakan dalam penelitian ini meliputi kerapu lumpur (Epinephelus coiodes), kerapu batik (E. microdon), kerapu macan (E. fuscoguttatus) dan kerapu bebek http://www.itk.fpik.ipb.ac.id/ej_itkt22 Melianawati et al. (Cromileptes altivelis). Parameter yang diamati meliputi pola perkembangan embrio, masa inkubasi dan tingkat penetasan. Pengamatan perkembangan embrio, baik menurut waktu dan suhu media inkubasi, dilakukan dengan cara mengambil 10-15 butir sampel telur secara acak dari ketiga media inkubasi masing-masing perlakuan dengan menggunakan pipet, menempatkannya pada single concave object glass dan kemudian mengamatinya di bawah mikroskop. Pengamatan dilakukan setiap interval waktu satu jam sampai dengan saat telur menetas. Masa inkubasi dihitung mulai dari saat inkubasi dilakukan hingga telur menetas seluruhnya. Klasifikasi perkembangan embrio (Chen et al., 1977) didasarkan atas 7 tingkat stadia (Tabel 1) untuk mempermudah dalam pengamatan perkembangan embrio. III. HASIL DAN PEMBAHASAN Hasil pengamatan terhadap perkembangan embrio telur ikan kerapu yang diamati dalam penelitian ini menunjukkan bahwa masing-masing stadia ditandai dengan karakteristik perkembangan embrio (Gambar 1). Hasil pengamatan menunjukkan bahwa perbedaan suhu inkubasi berpengaruh pada pola perkembangan embrio dan masa inkubasi telur ikan kerapu. Secara umum terlihat bahwa pada telur yang diinkubasikan pada suhu yang lebih tinggi perkembangan embrionya terjadi lebih cepat dibandingkan dengan yang diinkubasi pada suhu yang lebih rendah (Gambar 2, 3, 4, 5). Hal ini disebabkan karena pada suhu yang lebih tinggi proses metabolisme terjadi lebih cepat. Tabel 1. Klasifikasi perkembangan embrio (Chen et al., 1977) 1 Stadia Perkembangan embrio 1 Deskripsi Pembuahan hingga multisell 2 Blastula 3 Gastrula 4 Pembentukan bayangan embrio 5 Pembentukan kuppfer vesicle 6 7 Pergerakan embrio Penetasan embrio 2 3 4 5 6 7 Gambar 1. Stadia perkembangan embrio ikan kerapu (1. Multisel; 2. Blastula; 3. Gastrula; 4. Pembentukan bayangan embrio; 5. Pembentukan kuppfer vesicle; 6. Pergerakan embrio; 7. Penetasan embrio) Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 85 Perencanaan Waktu Tetas Telur Ikan Kerapu dengan… Hasil pengamatan pada telur ikan kerapu lumpur yang diinkubasikan pada kisaran suhu 24-31°C (perlakuan B, C dan D) cenderung membentuk pola yang hampir sama, dimana dari tujuh stadia perkembangan, perkembangan embrio pada stadia 5 cenderung berlangsung paling lama. Pola perkembangan embrio telur yang diinkubasi pada suhu terendah 21-22oC (perlakuan A) berbeda dengan pada perlakuan lainnya. Pada suhu inkubasi ini terjadi ketidakteraturan pola perkembangan embrio mulai dari awal hingga akhir masa inkubasi. Kondisi suhu media yang rendah diduga berpengaruh terhadap ketidakteraturan tersebut sehingga menghambat proses perkembangan embrio telur ikan kerapu lumpur (Gambar 2). Hasil pengamatan terhadap telur kerapu batik menunjukkan bahwa telur yang diinkubasi pada suhu 27-31 ºC cenderung memiliki pola perkembangan yang sama pada masing-masing stadianya. Perkembangan embrio pada stadia 1 sampai dengan stadia 3 berlangsung lebih lama daripada stadia 4, sedangkan pada stadia 5 cenderung berlangsung paling lama dan pada stadia 6 serta stadia 7 berlangsung paling singkat. Perbedaannya adalah pada kecepatan perubahan pada masingmasing stadia perkembangan embrio, dimana telur yang diinkubasi pada suhu 30-31 ºC perkembangan embrionya terjadi lebih cepat daripada yang diinkubasi pada media dengan suhu 2728 ºC. Perkembangan embrio dari telur yang diinkubasi pada suhu 24-25 ºC menunjukkan pola yang tidak teratur mulai stadia 1 sampai dengan stadia 3. 100% 80% 60% 40% P 20% e 0% r k 100% 80% e m 60% b 40% a 20% n 0% g a 100% n 80% 60% E 40% m 20% b 0% r i 100% o 80% 30-31oC 27-28oC 24-25oC 60% 40% 20% 0% 0:00 21-22oC 5:00 10:00 15:00 20:00 25:00 30:00 35:00 40:00 TIME (HOUR) S2 S3 S4 S5 S6 S7 Gambar 2. Pola perkembangan embrio telur ikan kerapu lumpur yang diinkubasi pada suhu berbeda (S: Stadia) 86 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Melianawati et al. Setelah melewati stadia 3 pola perkembangan embrio menunjukkan kecenderungan yang sama dengan telur yang diinkubasi pada suhu 27-31ºC. Telur yang diinkubasi pada suhu 21-22 ºC tidak berhasil menetas. Dari hasil pengamatan terlihat bahwa telur mengalami kematian embrio pada stadia 3. Hal ini menunjukkan bahwa suhu inkubasi berpengaruh terhadap perkembangan embrio telur ikan kerapu batik (Gambar 3). Pola perkembangan embrio telur ikan kerapu macan yang diinkubasikan pada kisaran suhu 24-31°C cenderung hampir sama. Dari 7 stadia perkembangan yang diamati, stadia 4 berlangsung dalam waktu yang paling singkat, sedangkan stadia 5 berlangsung paling lama. Perbedaan yang nampak dari setiap perlakuan adalah lama waktu perkembangan pada masing-masing stadia. Perkembangan masing-masing stadia pada telur yang diinkubasi pada suhu 24-25oC terjadi lebih lambat dibandingkan pada telur yang diinkubasi pada suhu 27-28oC. Sedangkan perkembangan stadia pada telur yang diinkubasi pada suhu 27-28oC itu sendiri masih lebih lambat dibandingkan pada telur yang diinkubasi pada suhu 30-31oC. Dengan demikian nampak bahwa suhu media inkubasi berpengaruh terhadap kecepatan perkembangan masing-masing stadia pada perkembangan embrio ikan kerapu macan. Sedangkan perkembangan embrio telur yang diinkubasi pada suhu 21-22oC hanya berlangsung sampai stadia 3 atau fase gastrula. 100% 75% 30-31 oC 50% P e r k e m b a n g a n 25% 0% 100% 75% 27-28 oC 50% 25% 0% 100% 75% E m b r i o 24-25 oC 50% 25% 0% 100% 75% 21-22 oC 50% 25% 0% 0:00 2:05 4:45 7:45 10:45 13:45 16:45 19:45 22:45 25:45 28:45 Time (hour) S1 S2 S3 S4 S5 S6 S7 Gambar 3. Perkembangan embrio telur kerapu batik yang diinkubasi pada suhu berbeda (S: Stadia) Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 87 Perencanaan Waktu Tetas Telur Ikan Kerapu dengan… Dengan demikian dapat dikatakan bahwa batas suhu terendah untuk inkubasi telur kerapu macan adalah 240C (Gambar 4). Hasil pengamatan terhadap telur ikan kerapu bebek yang diinkubasi pada kisaran suhu 24-31ºC cenderung memiliki pola perkembangan embrio yang sama. Perbedaan yang terjadi adalah pada lama waktu perkembangan masingmasing stadia, dimana perkembangan stadia terjadi lebih lambat pada suhu inkubasi yang lebih rendah dan sebaliknya. Perkembangan embrio pada stadia 1 sampai dengan stadia 4 terjadi relatif cepat, sedang perkembangan embrio yang paling lama terjadi pada stadia 5 (Gambar 5). Pola perkembangan embrio pada suhu inkubasi 21-22ºC nampak kurang teratur pada stadia 1 hingga stadia 2. Ketidakteraturan ini diduga karena ketidakmampuan telur untuk berkembang pada kondisi suhu inkubasi yang rendah. P e r k e m b a n g a n E m b r i o Hal ini terlihat dari paling banyaknya kematian telur yang terjadi pada stadia tersebut. Perkembangan embrio mulai teratur pada stadia 3 dan hal ini terus berlanjut hingga menetas. Pada suhu inkubasi ini waktu perkembangan yang paling lama terjadi pada stadia 6. Hal ini mungkin disebabkan karena embrio kesulitan untuk menetas. Suhu inkubasi yang rendah di dalam media penetasan diduga juga berpengaruh terhadap lambatnya proses penetasan telur. Dari hasil pengamatan dapat diketahui bahwa suhu inkubasi berpengaruh terhadap masa inkubasi. Telur ikan kerapu yang diinkubasi pada suhu lebih rendah, masa inkubasinya berlangsung lebih lama (dibandingkan telur yang diinkubasikan pada suhu lebih tinggi) (Tabel 2). Srihati (1997) mengemukakan bahwa suhu air berpengaruh terhadap penetasan telur dimana makin tinggi suhu air makin cepat terjadi penetasan telur. 100% 80% 60% 40% 20% 0% 100% 80% 60% 40% 20% 0% 100% 80% 60% 40% 20% 0% 100% 80% 60% 40% 20% 0% 0:00 3:00 6:00 9:00 12:00 15:00 18:00 21:00 S6 S7 TIME (hour) S1 S2 S3 S4 S5 Gambar 4. Pola perkembangan embrio telur ikan kerapu macan yang diinkubasi pada suhu berbeda (S: Stadia) 88 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Melianawati et al. Tabel 2. Masa inkubasi dan Tingkat penetasan telur ikan kerapu yang diinkubasi pada beberapa tingkatan suhu Jenis telur Kerapu Lumpur Suhu inkubasi Masa Inkubasi 21-22oC * 24-25oC 24 jam 30 menit 27-28oC 17 jam 30 menit 30-31oC 14 jam 30 menit Kerapu batik 21-22oC * 24-25oC 29 jam 45 menit 27-28oC 19 jam 45 menit 30-31oC 15 jam 45 menit Kerapu macan 21-22oC * 24-25oC 21-23 jam 27-28oC 18-22 jam 30-31oC 16-21 jam Kerapu bebek 21-22oC 45 jam 45 menit 24-25oC 27 jam 45 menit 27-28oC 18 jam 45 menit 30-31oC 14 jam 45 menit * Perkembangan embrio terhenti pada stadia 2 dan 3 Tingkat penetasan 0% 36% 50% 82% 0% 58% 100% 100% 0% 84% 100% 100% 3% 57% 100% 100% 100% 75% 50% 30-31 ºC 25% 0% 100% 75% 50% 27-28 ºC 25% 0% 100% 75% 24-25 ºC 50% 25% 0% 100% 75% 21-22 ºC 50% 25% 0% 1 7 13 19 25 31 37 43 WAKTU (jam) S1 S2 S3 S4 S5 S6 S7 Gambar 5. Pola perkembangan embrio telur ikan kerapu bebek yang diinkubasi pada suhu berbeda (S: Stadia) Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 89 Perencanaan Waktu Tetas Telur Ikan Kerapu dengan… Secara alami masa inkubasi telur ikan kerapu lumpur tergantung pada suhu media, seperti halnya pada kegiatan pembenihan ikan laut di Singapore, masa inkubasi telur ikan kerapu lumpur (E. tauvina) dilaporkan berlangsung selama 23-25 jam pada suhu 27-28°C dan selama 20-22 jam pada suhu 29-30°C (Cheong dan Chuan, 1980) atau setelah 23-25 jam masa inkubasi pada suhu 27°C (Chen et al., 1977), sedangkan di Kuwait penetasan telur jenis ikan tersebut terjadi setelah 26-35 jam inkubasi pada kondisi suhu 27-30°C (Hussain et al., 1975). Masa inkubasi telur ikan kerapu batik secara alami berlangsung sekitar 18 jam 30 menit pada suhu air 27-29 ºC (Slamet dan Tridjoko, 1997). Pada kondisi normal telur ikan kerapu bebek menetas setelah inkubasi selama 17 jam 45 menit pada kisaran suhu 27,5-30,5ºC dan salinitas 31-33 ppt (Tridjoko et al., 1996). Dari keempat jenis telur kerapu yang digunakan dalam penelitian ini hanya telur kerapu bebek yang embrionya dapat berkembang pada suhu inkubasi 21-22oC, sedangkan pada telur kerapu lumpur, kerapu batik dan kerapu macan perkembangan embrio terhenti pada stadia 2 atau 3. Hasil ini menunjukkan bahwa pemberian suhu inkubasi yang rendah pada fase awal inkubasi mengakibatkan terhambatnya proses pembelahan sel. Sugama et al (2001) mengemukakan bahwa fase yang sangat peka dalam perkembangan telur adalah sebelum stadia embrio, terutama sebelum mencapai stadia blastula. Untuk telur-telur yang dapat melewati fase kritis tersebut, selanjutnya dapat terus berkembang dengan baik hingga mencapai stadia embrio dan menetas dengan bentuk tubuh normal. Telur kerapu bebek yang diinkubasi pada suhu 21-22ºC mempunyai masa inkubasi yang paling lama yaitu 45 jam 45 menit. Larva yang 90 berhasil menetas tubuhnya tidak normal dan bengkok. Ketidaknormalan tubuh larva ini kemungkinan dipengaruhi oleh masa inkubasi, dimana masa inkubasi yang terlalu lama mengakibatkan pertumbuhan embrio di dalam telur juga terlalu lama dan kurang sempurna sehingga embrio menjadi tidak normal. Larva yang bentuk tubuhnya tidak normal tidak akan dapat bertahan hidup. IV. KESIMPULAN Telur yang diinkubasikan pada suhu 24-31°C menghasilkan pola perkembangan embrio yang teratur, sedangkan pada suhu 21-22°C perkembangan embrio terhenti pada stadia blastula atau gastrula atau embrio terus berkembang tetapi menghasilkan larva yang tidak normal tubuhnya. Suhu inkubasi 24-25oC dapat dimanfaatkan untuk menunda waktu tetas dalam kaitannya untuk transportasi telur jarak jauh, meskipun tingkat penetasan telurnya lebih rendah dibandingkan pada suhu 27-31oC. Kisaran suhu optimum bagi penetasan telur kerapu lumpur, kerapu batik, kerapu macan dan kerapu bebek adalah 24-31oC, sedangkan batas toleransi suhu terendah dalam kaitannya untuk mengatur masa inkubasi dan perencanaan waktu tetas adalah 24oC. DAFTAR PUSTAKA Andamari, R., B. Teguh dan Mujimin. 2005. Kajian ekspor kerapu dari propinsi Bali. Dalam: Sudrajat et al. (2005). Buku Perikanan Budidaya Berkelanjutan. Hal:259-268. Chen, F.Y., M. Chow, T.M. Chao and R. Lim. 1977. Artificial spawning and larval rearing of the grouper, Epinephelus tauvina (Forskal) in Singapore. Singapore J. Pri. Ind., 5(1):1-21. http://www.itk.fpik.ipb.ac.id/ej_itkt22 Melianawati et al. Cheong, L. and L.L. Chuan. 1980. Current work on the induced breeding of grouper Epinephelus tauvina, F. in Singapore. 4pp. Hussain, N., M. Saif, and M. Ukawa. 1975. On the culture of Epinephelus tauvina (Forskal). Kuwait institute for scientific research. 14 pp. Kohno, H., P.T. Imanto, S. Diani, B. Slamet, and P. Sunyoto. 1990. Reproductive performance and early life history of the grouper, Epinephelus fuscoguttatus. Bull. Penelitian perikanan, 1(special edition):27-35. Lau, P.P.F. and R.P. Jones, 1999. The Hongkong trade in live reef fish for food. TRAFFIC East Asia and World Wide Fund for Nature Hongkong. 64 p. Putro, D.H., Evalawati dan P. Hartono. 1999. Pengamatan pendahuluan pembesaran kerapu bebek (Cromileptes altivelis) di karamba jaring apung. Bulletin Budidaya Laut, 12:5-8. Setiadharma, T., A.Prijono dan T. Ahmad. 1997. Pengaruh kepadatan pasa pengangkutan dengan system tertutp terhadap daya tetas telur bandeng (Chanos chanos. Forsskal). Jurnal Penelitian Perikanan Indonesia, 3(1):68-72. Slamet, B. 1993. Pengaruh penurunan suhu media terhadap penundaan penetasan dan peningkatan optimasi kepadatan pada transportasi telur ikan kerapu macan, Epinephelus fuscoguttatus. J. Penelitian Budidaya Pantai, 9(5):30-36. Slamet, B. dan Tridjoko. 1997. Pengamatan pemijahan alami, perkembangan embrio dan larva ikan kerapu batik, E. microdon, dalam bak terkontrol. J. Penelitian Perikanan Indonesia. Edisi khusus 3(4):40-50. Srihati. 1997. Pengaruh suhu terhadap penetasan telur, pertumbuhan dan daya tahan hidup larva ikan bandeng (Dicentranchus labrax L.) Seminar biologi XV. Bandar Lampung: 872-876. Sudaryanto, Sudjiharno dan P. Hartono. 1999. Upaya mengubah kelamin pada kerapu bebek (Cromileptes altivelis). Bulletin budidaya laut 12, Lampung:1-4. Sudjiharno, H. Minjoyo, E. Sutrisno dan Mustamin. 2001. Teknologi produksi massal benih kerapu macan (Epinephelus fuscogut-tatus) di Balai Budi Daya Laut Lampung. Dalam Sudradjat, A., Heruwati, E.S., Poernomo, A., Rukyani, A., Widodo, J. dan Danakusumah, E. 2001. Teknologi Budi Daya Laut dan Pengembangan Sea Farming di Indonesia. Puslitbang Eksplorasi Laut dan Perikanan. 489 p. Sugama, K., Tridjoko, B. Slamet, S. Ismi, E. Setiadi, dan S. Kawahara 2001. Petunjuk teknis produksi benih ikan kerapu bebek Cromileptes altivelis. Balai Riset Budidaya Laut Gondol dan Japan International Cooperation Agency. 40p. Tridjoko, B. Slamet, D. Makatutu, dan K. Sugama.1996. Pengamatan pemijahan dan perkembangan telur ikan kerapu bebek (Cromileptes altivelis) pada bak secara terkontrol. Jurnal Penelitian Perikanan Indonesia, 2(2):55-62 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 91 Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Hal. 92-108, Desember 2010 MASSA AIR SUBTROPICAL DI PERAIRAN HAMAHERA SUBTROPICAL WATER MASSES IN HALMAHERA WATERS Hadikusumah Bidang Dinamika Laut – Pusat Penelitian Oseanografi – LIPI Jl. Pasir Putih No.1, Ancol Timur, P.O. Box 480 /JKTF. 14430 E-mail: [email protected] U U ABSTRACT Research of water masses by using conductivity temperature depth (CTD), are conducted in the eastern path of the Indonesia Throughflow (ITF) in the Halmahera, Seram and Banda seas during March-April 2007 under the Expedition of Widya Nusantara (EWIN). The objective of this research is to see maximum salinity spread of South Pacific Subtropical Water (SPSW) water masses enter the eastern Indonesia Waters. The temperature and salinity profiles show the presence of the presence of SPSW have been very much confined to the Halmahera Sea only. Little of this water masses have been detected in the eastern Seram Sea, but none in Banda Sea. Early data of Arlindo Mixing (ARMIX) experiment in southeast monsoon 1993 indicated that this water masses SPSW may entered the southern most part of the Moluccas Sea. Type of South Pacific Subtropical Water (SPSW) water masses appears in the Halmahera Sea at an average depth of 200m and the dominant flows between Halmahera and Obi Islands (Moluccas Sea continues to the east). Type of South Pacific Intermediate Water (SPIW) water mass appeared on average Halmahera Sea at a depth of 750m and the dominant flows between Halmahera and Obi Islands (Moluccas Sea continues to the east). Type of North Pacific Subtropical Water (NPSW) water masses at an average depth of ~ 150m found in the northern part of Halmahera, the dominant flow to the Celebes Sea, Makassar Strait, Flores Sea and partly flows into Lombok Straits. Type of minimum salinity water mass of North Pacific Intermediate Water (NPIW) obtained at an average depth of ~ 400m dominant flow towards the Celebes Sea, Makassar Strait and Flores Sea. Keywords: maximum salinity, SPSW, Halmahera, Seram, and Banda Seas ABSTRAK Penelitian massa air dengan menggunakan conductivity temperature depth (CTD), dilakukan di bagian timur Arus Lintas Indonesia (ARLINDO – ITF) di Laut Halmahera, Laut Seram dan Laut Banda dari bulan Maret - April 2007 pada pelayaran Ekspedisi Widya Nusantara (EWIN). Tujuan penelitian ini untuk melihat penyebaran salinitas maksimum dari massa air South Pacific Subtropical Water (SPSW) masuk ke bagian timur Indonesia. Profil suhu dan salinitas menunjukkan adanya kehadiran SPSW sudah sangat terbatas ke Laut Halmahera saja. Sebagian kecil massa air ini telah terdeteksi di bagian timur Laut Seram, tapi tidak ke Laut Banda. Awal data Arlindo Mixing (ARMIX) eksperimen di musim tenggara 1993 menunjukkan bahwa massa air SPSW ini dapat masuk ke bagian paling selatan Laut Maluku. Jenis masa air South Pacific Subtropical Water (SPSW) muncul di Laut Halmahera pada rata-rata kedalaman 200m serta dominan mengalir di antara Pulau Halmahera dan Pulau Obi (terus ke Laut Maluku bagian timur). Jenis massa air South Pacific Intermediate Water (SPIW) muncul di Laut Halmahera rata-rata pada kedalaman 750m serta dominan mengalir di antara Pulau Halmahera dan Pulau Obi (terus ke Laut Maluku bagian timur).Jenis massa air North Pacific Subtropical Water (NPSW) pada rata-rata kedalaman ~150m didapatkan di bagian utara Halmahera, dominan mengalir menuju ke Laut Sulawesi, Selat Makassar, Laut Flores dan sebagian mengalir ke Selat Lombok. Jenis massa air bersalinitas minimum North Pacific Intermediate Water (NPIW) didapatkan pada rata-rata kedalaman ~400m dominan mengalir ke arah Laut Sulawesi, Selat Makassar, dan Laut Flores. Kata Kunci: salinitas maksimum, SPSW, Laut Halmahera, Laut Seram dan Laut Banda 92 ©Ikatan Sarjana Oseanologi Indonesia dan Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB Hadikusumah I. PENDAHULUAN Perairan Indonesia terletak diantara Samudera Pasifik dan Hindia. Massa air dari Pasifik masuk dan menyebar di perairan Indonesia sebelum mengalir keluar Indonesia (Wyrtki, 1956). Empat jenis massa air telah diketahui dan menyebar pada lapisan termoklin dan lapisan dalam, yaitu massa air North Pacific Subtropical Water (NPSW) dan North Pacific Intermediate Water (NPIW) yang merupakan massa air yang dibawa oleh arus Mindanao Eddy dan arus North Equatorial Current (NEQ), massa air South Pacific Subtropical Water (SPSW) dan South Pacific Intermediate Water (SPIW) yang merupakan massa air yang dibawa oleh arus New Guinea Coastal Current (NGCC) dan arus South Equatorial Current (SEQ) (Tomczak & Godfrey, 1994). NPSW adalah massa air dari Pasifik utara yang bergerak ke arah selatan dan masuk ke Laut Sulawesi pada lapisan dangkal (lapisan termoklin); SPSW yaitu massa air dari Pasifik Selatan yang bergerak ke arah Laut Halmahera lewat perairan pantai utara Pulau Papua di lapisan dangkal (lapisan termoklin). Keberadaan massa air tersebut diindikasikan oleh salinitas maksimum (Smaks). Di bawah lapisan salinitas maksimum (NPSW) terdapat lapisan massa air NPIW yaitu massa air dari Pasifik utara yang bergerak ke arah selatan dan masuk ke Laut Sulawesi pada lapisan dalam; SPIW adalah massa air dari Pasifik selatan yang bergerak ke arah Laut Halmahera lewat utara Pulau Papua terletak di lapisan dalam (Wyrtki, 1962) dan (Ffield, 1994). Keberadaan massa air tersebut diindikasikan oleh salinitas minimum (Smin). Pada lapisan pycnocline (100 sampai 300 m) ada lapisan salinitas maksimum dari utara dan selatan Samudera Pasifik. Massa air ini disebut Northern/Southern Subtropical Lower Waters (Wyrtki, 1961) atau North/South Pacific Tropical Waters (Fine et al., 1994) dalam (Yuji et al., 1996). Gambaran umum dari sirkulasi di perairan lautan Indonesia yang menunjukkan dengan jelas pembalikan aliran musiman pola arus permukaan dan distribusi massa air laut dalam yang lebih jelas berasal dari berbagai lokasi Samudra Pasifik bagian utara atau bagian barat Samudera Pasifik. Lautan Indonesia merupakan satu-satunya penghubung antar samudera antara Pasifik bagian barat dengan Samudra Hindia bagian timur (Birowo 1990). South Pacific Central Water (SPCW) melewati Halmahera Sea (HS) ke South Banda (BS) dan Timor Sea (TS). North Pacific Central Water (NPCW) melewati Makassar Strait (MS) ke Timor Sea (TS). Keduanya kemudian diubah menjadi Indian Central Water (ICW) (Ffield et al., 1992). Air bersalinitas asal Pasifik Selatan (South Pacific) diamati di bawah lapisan termoklin di Laut Seram dan Laut Maluku bagian selatan terutama pada musim barat laut (Ilahude dan Gordon, 1996). Di dalam perairan Indonesia, mass air berstratifikasi dari Samudera Pasifik secara nyata diubah oleh pencampuran vertikal seperti maksimum salinitas yang berbeda yang berasal dari North Pacific (salinitas 34,8psu pada 100m) dan South Pacific (salinitas 35,4psu pada 150m) akhirnya hilang. Akibatnya, pada saat air throughflow meninggalkan Indonesia untuk memasuki Samudra Hindia mereka membawa salinitas homogen (34,6psu) melalui termoklin bagian atas, dinyatakan oleh (Ffield et al., 2005). Air subtropical Samudera Pasifik bagian selatan bersalinitas tinggi tidak menyebar ke bagian atas termoklin di Laut Banda dari pintu masuknya di Laut Halmahera (Gordon 2005). Air bagian atas South Pacific mengalir menuju searah jarum Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 93 Massa Air Subtropical Di Perairan Hamahera jam sekitar Halmahera eddy (HE) dan kembali untuk menggabungkan dengan arus kearah timur dari Midanau Current (MC). Sejumlah kecil air massa air South Pacific memasuki Laut Seram dan terbagi menjadi dua cabang. Cabang ke arah utara kembali ke Samudera Pasifik dan ke arah selatan mengalir memasuki Laut Banda (Liu et al., 2005). Tujuan penelitian EWIN Ambon 2007 salah satunya ialah untuk melihat sebaran salinitas maksimum masa air South Pacific Subtropical Water (SPSW) dan North Pacific Subtropical Water (NPSW) dan sebaran salinitas minimum masa air South Pacific Intermediate Water (SPIW) dan North Pacific Intermediate Water (NPIW) di perairan Indonesia Timur. II. METODE PENELITIAN Pengukuran parameter Oseanografi fisika di sekitar perairan Laut Halmahera, Laut Seram, Laut Banda dan Teluk Ambon telah dilakukan dari tanggal 8 Maret s.d 3 April 2007 dengan menggunakan KR. Baruna Jaya VII dalam program Ekspedisi Widya Nusantara (EWIN). Jumlah stasiun oseanografi CTD yaitu 30 stasiun (Gambar 1). Parameter fisika yang diukur adalah suhu, salinitas dan turbiditas dengan menggunakan instrumen CTD Model SBE-91. Profil data pengukuran CTD dengan interval kedalaman 0.5 94 meter di stasiun-stasiun perairan dangkal (stasiun biologi) dan 1 meter di stasiun laut dalam. Stasiun CTD di bagian selatan Halmahera dan bagian tengah Laut Seram hampir berimpit dengan program Arlindo Mixing (ARMIX) 1993 pada musim tenggara (Ilahude & Gordon 1996). Untuk melengkapi analisa massa air Pacific bagian selatan ke perairan Laut Halmahera bagian selatan dan Laut Seram telah ditambah data WOCE Line 08 dan 09. Analisa data suhu dan salinitas yaitu dengan membuat distribusi salinitas penampang vertikal dan korelasi antara salinitas dan suhu dalam diagram T-S. III. HASIL DAN PEMBAHASAN 3.1. Kondisi Arus Arus di stasiun MBIO-2A di bagian timur Pulau Misol pada kedalaman 5m diperoleh kecepatan arus bervariasi dari 0,18 – 74,34 cm/dt dan hasil analisis didapatkan rata-rata komponen arus timur barat (TB) ialah 0,96 cm/dt dan komponen arus utara selatan (US) ialah 1,99 cm/dt. Hasil analisis progressive vector diagram (PVD) didapatkan mampu menempuh jarak 3,37 km dalam waktu 42 jam 22 menit dan resultante arahnya (current direction) ialah 154,22° atau arus bergerak ke arah tenggara, serta arus sisanya (residual current) sebesar 2,21 cm/dt (Gambar 2). http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah 14 4 PACIFIC OCEAN 2 Latitude, N/S 27 0 Waigeo 37 38 39 Biak 1 2 3 4 4041 54243 6 Misol 7 8 44 9 10 Seram Sea 13 14 11 12 45 46 15 Seram 161718 Buru 192021 222324 252627 2829 30 Obi -2 Mangole -4 Yapen 105 PAPUA BANDA SEA -6 126 128 130 132 134 136 138 140 142 144 Longitude, E Gambar 1. Peta dan stasiun oseanografi Ekspedisi Widya Nusantara (EWIN) pada tahun 2007 di stasiun bertanda lingkaran hitam dan Arlindo mixing (ARMIX) pada ekspedisi selama musim tenggara tahun 1993 di stasiun bertanda lingkaran biru dan merah dan WOCE di stasiun bertanda hijau PVD arus, Jam 21:10 s/d 13:00, 10 - 12 Maret 2007 Posisi: 2° 4.063 S; 130° 15.133 E, Kampung Lelintah - P. Misol 0 -5 -4 -3 -2 -1 0 1 2 22 Pergeseran Komponen Arus, US (km) -0.5 0 -1 3 2 1 6 18 -1.5 12 9 -2 0 -2.5 3 6 12 9 -3 -3.5 Pergeseran Komponen Arus, TB (km) Gambar 2. Proressive Vector Diagram arus di stasiun MBIO-1, Jam 21:10 s/d 13:00, 10 - 12 Maret 200, Posisi: 2° 4.063 S; 130° 15.133 E, Kampung Lelintah - P. Misol Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 95 Massa Air Subtropical Di Perairan Hamahera Arus di stasiun MBIO-13 di bagian timur Pulau Seram pada kedalman 5m didapatkan kecepatan arus bervariasi dari 0,6 – 31,9 cm/dt dan rata-rata komponen arus TB ialah 5,5 cm/dt dan komponen arus utara selatan (US) ialah -2.2 cm/dt. Hasil analisis arus PVD didapatkan mampu menempuh jarak 5,84 km dalam waktu 27 jam 20 menit dan resultante arahnya (current direction) ialah 111,7° atau arus menuju ke arah tenggara, serta arus sisanya (residual current) ialah 5,92 cm/dt. Arus di stasiun MBIO-14 diperoleh kecepatan arus bervariasi dari 0.30 – 43.80 cm/dt dan hasil analisa diperoleh rata-rata komponen arus TB ialah -1.11 cm/dt dan komponen arus US ialah -0.13 cm/dt. Hasil analisa progressive vector diagram diperoleh bahwa PVD arus dapat menempuh jarak 0.32 km dalam waktu 7 jam 55 menit dan resultante arahnya (current direction) ialah 262.95 ° atau arus ke arah barat daya, serta arus sisanya (residual current) ialah 1.12 cm/dt (Gambar 3). Pola arus di stasiun MBIO- 2A, MBIO-13 dan MBIO-14 menandakan bahwa masa air tersebut bergerak ke arah tenggara sampai barat daya dan ini merupakan masa air ARLINDO di bagian permukaan. Berdasarkan data arus mooring 1993-1994 selama setahun pada stasiun 29; dari Program Kerjasama antara ASEAN dan Australia di Laut Halmahera bahwa kecepatan arus rata-rata maksimum pada kedalaman 428m didapatkan tertinggi (25,95cm/dt) pada bulan Februari, jika dibandingakan kecepatan arus pada kedalaman 720m (19,42 cm/dt) pada bulan Desember dan kecepatan arus paling rendah pada kedalaman 912m (16,90cm/dt) bulan Desember. Hasil analisis progressive vector diagram (PVD) menunjukkan bahwa pergeseran arus selama satu tahun pada kedalaman 720 m didapatkan terbesar (2875,6km) dengan arah arus dominan bergerak ke selatan (188°) atau masuk ke perairan Indonesia (ARLINDO), dibandingkan dengan kedalaman 428m, pergeseran arusnya 0.45 PVD arus di stasiun MBIO-14 di Tl. Seleman Posisi 2° 57.321' S; 129° 07.19' E tanggal 2 April 2007 0.4 15 0.35 0.3 Pergeseran komponen arus, US (km) 12 0.25 0.2 0.15 0.1 0.05 0 -0.35 -0.3 -0.25 18 -0.2 -0.15 -0.1 -0.05 0 0.05 -0.05 -0.1 Pergeseran komponen arus, TB (km) Gambar 3. Proressive Vector Diagram arus di stasiun MBIO-14 di Teluk Seleman Posisi: 2° 57.321' S; 129° 07.19' E tanggal 2 April 2007 96 http://www.itk.fpik.ipb.ac.id/ej_itkt22 0.1 Hadikusumah (556,86km) dengan arah arus dominan ke barat daya (204°) serta kedalaman 912m, pergeseran arusnya paling rendah (453km) dengan dominan arah arus bergerak ke barat daya (264°) atau ke perairan Indonesia. Pola arus antara bulan Mei s.d September arah arus dominan menuju ke arah barat laut, sebaliknya antara bulan Oktober s.d April arah arus dominan menuju ke arah dominan ke arah selatan (Gambar 4). 500 Current displacement comp_NS (km) October August 93 -600 -500 -400 -300 -200 0 -100 0 100 December January 94 -500 -1000 March -1500 June Mooring #5, 428 m depth Halmahera Strait -2000 Current displacement comp_EW (km) Gambar 4. Proressive Vector Diagram arus di stasiun mooring di Laut Halmahera pada kedalaman 428m dari tahun 1993/1994 3.2. Karakteristik dan Distribusi Tegak massa Air Salinitas maksimum 34,988psu (St.1) pada profil salinitas didapatkan pada kedalaman 236m yaitu pada lapisan termoklin. Dari perairan yang diobservasi ternyata didapatkan ada dua massa air yang berbeda dari permukaan sampai kedalaman ~700 m, yaitu masa air besalintas rendah di Laut Banda Seram timur dan salinitas tinggi di Laut Halmahera sampai Laut Seram bagian barat. Sedangkan massa air permukaan bersalinitas rendah (kedalaman <185m) di St.15 didapatkan sama dengan salinitas Laut Banda, sedangkan di kedalaman >185m didapatkan masa air bersalinitas tinggi, demikian pula salinitas maksimum masih didapatkan di St-16 (34,65psu) di kedalaman >164m. Ini menandakan bahwa pertemukan atau front masa air besalinitas tinggi dan rendah ada di antara St.15 sampai 16. Distribusi horizontal salinitas di bagian permukaan (1m) didapatkan antara 32 s.d 34psu, dimana salinitas Selat Halmahera dan Laut Seram didapatkan lebih tinggi dibandingkan salinitas Laut Banda atau perairan Seram timur. Distribusi horizontal salinitas pada kedalaman 50m didapatkan antara 33,5 s.d 34,3psu, di mana salinitas lepas pantai Selat Halmahera (>34,2psu) dan salinitas perairan lepas pantai Laut Seram Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 97 Massa Air Subtropical Di Perairan Hamahera (>34,1psu) didapatkann lebih tinggi dibandingkan salinitas Laut Banda (<34psu) atau perairan Seram timur, sedangkan salinitas perairan pantai lebih rendah (<33,5 psu). Distribusi tegak salinitas antara St.1 sampai 4 bahwa antara St.1 sampai 2 didapatkan inti (core) salinitas <34,9psu pada kedalaman 200m dengan ketebalan inti salinitas makin ke arah timur makin berkurang. Salinitas minimum (<34,6psu) juga didapatkan di laut dalam pada kedalaman >465m dan diduga sampai dasar. Distribusi salinitas vertikal antara St.1 sampai 11 didapatkan bahwa core salinitas maksimum di St.1 (<34,9psu) dengan ketebalan ~120m dan makin ke arah selatan (St.11) core salinitasnya makin berkurang (>34,8psu) dengan ketebalan <70m. Sedangkan salinitas minimum (<34,6psu) didapatkan pada kedalaman >440m dan diduga sampai dasar. Distribusi tegak salinitas antara St.2 sampai 12 didapatkan bahwa core salinitas (>34,9 psu) di St.2 dengan ketebalan ~ 60m dan makin ke arah selatan nilai salinitasnya makin berkurang atau mengecil (>34,7) yaitu antara St.8 s.d (di Laut Seram) dengan ketebalan ~60m. Sedangkan salinitas minimum di laut dalam didapatkan pada kedalaman ~ 460m (St.2) dan makin ke arah selatan (dangkal sampai kedalaman ~360m). Distribusi tegak salinitas antara 98 St.11 sampai 14 (di Laut Seram) bahwa core salinitas (>34,8 psu) didapatkan pada kedalaman 200m dengan ketebalan ~ 65m. Core salinitas makin ke arah timur didapatkan nilai salinitasnya makin berkurang dan sampai di St.14 (>34,6psu) dengan ketebalan <20m. Salinitas <34,6psu didapatkan di laut dalam di St.11 dikedalaman ~ 400m dan makin ke arah timur salinitas (<34,6psu) didapatkan di kedalaman ~ 240m (makin dangkal atau naik). Distribusi salinitas vertikal antara St.1 sampai 30 yaitu dari Selat Hamahera, Laut Seram dan Laut Banda bahwa salinitas di bagian lapisan permukaan antara St.1 sampai St.13 didapatkan salinitas lebih besar (>34 psu) dibandingkan salinitas di bagian permukaan ke arah selatan Laut Banda (<34psu). Inti salinitas di St.1 adalah sama seperti pembahasan di atas, didapatkan bahwa salinitas makin ke arah selatan core salinitas Selat Halmahera sudah tidak muncul lagi. Core salinitas >34,6 psu masih muncul di St.15 pada kedalaman 200m. Ini artinya bahwa salinitas maksimum tersebut tidak mengalir secara kuat ke arah timur ke arah Laut Banda. Karena di St.24 sampai 27 dan 30 nilai salinitas >34,6psu sudah tidak muncul lagi, sehingga yang disebut lapisan minimum salinitas (<34,6 psu) di lapisan dalam sudah tidak terstatifikasi lagi (Gambar 5). http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah St_30 27 24 21 18 15 13 8 5 1 0 depth (m) -200 > -400 -600 -800 Salinity (psu) -1000 -4.5 -4.0 -3.5 -3.0 -2.5 -2.0 -1.5 Latitude, S Gambar 5. Distribusi tegak salinitas antara St.1 sampai 30 dari Laut Halmahera, Laut Seram dan Laut Banda. Distribusi tegak salinitas antara St.4 sampai 28 bahwa salinitas perairan dangkal di lapisan permukaan (mixed layer depth) bagian timur Pulau Misol didapatkan salinitas (>34 psu) yaitu antara St.4 sampai St.14. Sedangkan salinitas makin ke arah selatan, salinitas makin berkurang (<34psu). Demikian juga bahwa core salinitas (>34,6psu) masih muncul didapatkan di St.15 pada kedalaman ~ 200m. Ini menandakan bahwa salinitas (>34,6psu) masih menyebar ke arah timur, meskipun core salinitas tersebut sudah berkurang 0,3psu dari St.1. Demikian salinitas <34,6psu didapatkan salinitas minimum di kedalaman laut dalam sudah tidak terstatifikasi lagi, sama seperti pada distribusi tegak salinitas antara St.1 sampai St.30 (Gambar 6). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 99 Massa Air Subtropical Di Perairan Hamahera St_28 25 22 19 17 15 14 10 7 4 0 Depth (m) -200 -400 -600 Salinity (psu) -800 -1000 -4.5 -4.0 -3.5 Latitude, S -3.0 -2.5 -2.0 Gambar 6. Distribusi tegak salinitas antara St.4 sampai 28 di Laut Seram sampai Laut Banda Distribusi tegak salinitas Line 08 bahwa inti salinitas maksimum >35,3psu didapatkan pada kedalaman rata-rata 110m (~100 – 175m) antara St.23 sampai St.27. Salinitas maksimum tersebut berada dibagian mulut Laut Halmahera antara Pulau Halmahera dan Pulau Papua, dan makin ke arah utara nilai salinitas makin berkurang. Salinitas >35,2psu, posisi penyebarannya bergerak naik ke arah permukaan antara kedalaman 90m s.d 100m (St.23 sampai St.19) yang disebut masa air South Pacific Subtropical Water (SPSW). Salinitas 35,1psu didapatkan di St.27 berada antara kedalaman 90m s.d 220m dan makin ke arah utara ketebalan lapisan salinitas makin menipis (80m sampai 100m) di St.13. Salinitas 35,0psu 100 didapatkan di St.27 terletak antara kedalaman 90m s.d 225m. Gradasi sebaran salinitas 35,0 didapatkan makin ke arah utara ketebalan lapisan salinitas makin menipis (100m s.d 200m) atau disebut masa air North Pacific Subtrophical Water (NPSW). Inti salinitas minimum <34,5psu didapatkan rata-rata pada kedalaman 360m antara St.23 sampai St.13 (5°N). Di bagian selatan antara St.27 s.d 24 inti salinitas minimum <34,5psu tidal muncul lagi. Inti salinitas <34,6psu didapatkan pada lapisan atas yaitu antara kedalaman ~520m (St.27) dan ~250m (St.25 sampai St.13), dan pada lapisan bawah rata-rata pada kedalaman ~1500m (antara suhu 3°C s.d 8°C). Masa air bersalinitas rendah di lapisan bawah tersebut disebut http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah dengan masa air North Pacific Intermediate Water (NPIW). Selanjutnya makin kebawah didapatkan salinitas >34,6psu dengan suhu <3°C s.d <1°C. Distribusi tegak salinitas Line 09 bahwa inti salinitas maksimum lebih besar 35,5psu didapatkan antara kedalaman 150m s.d 180m antara St.105 s.d St.103. Salinitas >35,3psu didapatkan antara kedalaman 125m s.d 140m antara St.105 s.d St.92 dan salinitas >35,0psu didapatkan antara kedalaman 60m s.d 300m (St.105), dan makin ke arah utara ketebalan sebaran salinitas makin menipis antara kedalaman 95m s.d 140m (St.79), makin ke arah utara lagi (St.76) nilai salinitas didapatkan makin mengecil (>34,8psu). Nilai salinitas 34,6psu didapatkan antara kedalaman 70m s.d 520m (St.105), dan makin ke arah utara ketebalan lapisan salinitas 34,6psu makin menipis dari kedalaman ~140 m s.d 75m (St.76). Sedangkan inti salinitas <35,5psu secara ruang baik memanjang maupun vertikal di Line 08 pada kedalaman yang sama tidak muncul lagi. Sedangkan salinitas <34,6psu didapatkan antara kedalaman ~500m s.d 1500m dan makin ke arah utara lapisannya makin menebal antara ~150m s.d 1500m (St.76). Masa air bersalinitas demikian disebut dengan massa air South Pacific Intermediate Water (SPIW). Massa air SPSW dan SPIW di Selat Damper, Raja Empat sudah didapatkan oleh Hadikusumah (2009). Penampang tegak salinitas Line 09 di utara Papua dapat disimpulkan bahwa inti salinitas (>35,5psu) yang paling tinggi berada di pantai utara Papua, dan makin ke arah utara (5°N), inti salinitas >35,5psu makin berkurang atau menurun. Ketebalan lapisan salinitas (>35psu) di bagian selatan (St.105) sampai mencapai 240m dan dibagian utara (5°N) didapatkan hanya 45m (St.79). Salinitas maksimum demikian disebut dengan massa air South Pacific Subtropical Water (SPSW) dari Samudera Hindia. Penampang tegak salinitas di Laut Flores antara St.1 sampai St.7 dapat disimpulkan bahwa inti salinitas (34,551psu) adalah massa air yang sudah terencerkan oleh masa air Laut Jawa, dibandingkan dengan salinitas maksimum Selat Makassar (34,862psu) disebut dengan masa air North Pacific Subtrophical Water (NPSW). Salinitas minimum (<34,5) pada kedalaman ~400m yang mempunyai pola kedalaman yang sama dengan salinitas minimum laut dalam Selat Makassar disebut dengan masa air North Pacific Intermediate Water (NPIW) (Gambar 7) seperti diuraikan oleh Hadikusumah (2008). 3.3. Diagram T-S Dalam membahas TS-diagram sekaligus dibandingkan dengan data ARMIX 93 dan WOCE, didapatkan ada enam jenis masa air yaitu: (1). Jenis masa air campuran antara Laut Banda dan Laut Seram yaitu antara sigma-t <21 s.d <22 pada lapisan antara permukaan sampai kedalaman <100m dengan suhu antara 29,24 °C s.d 28,56 °C dan salinitas antara 33,068 psu a/d 33,942 psu disebut masa air lokal Laut Banda dan Seram (BSW); (2). Jenis masa air campuran antara Selat Halmahera dan perairan Misol dengan sigma-t >21 s.d 23 yaitu dari permukaan s.d <100m, dengan suhu antara 29,26 s.d 27,00 °C dan salinitas antara 33,901 s.d 34,213psu; (3). Jenis massa air Laut Banda antara sigma-t 25 – 26, dengan suhu antara 17,47 s.d 13,10°C dan salinitas antara 34,512 s.d 34,512psu antara kedalaman <200 m s.d 400m; (4). Jenis masa air South Pacific Subtropical Water (SPSW) merupakan masa air Laut Halmahera bagian selatan, Laut Seram antara salinitas 34,987 s.d 34,721psu dalam sigma-t ~25 s.d ~26 dengan suhu 16,04 s.d 15,95°C pada rata-rata Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 101 Massa Air Subtropical Di Perairan Hamahera kedalaman 200m; merupakan massa air Laut Banda bagian Seram timur pada salinitas 34,663 psu dalam sigma-t ~26 dengan suhu 14,96°C (Gambar 8), St.1 2 0 merupakan massa air yang mengalir diantara Pulau Halmahera dan Pulau Obi dan Laut Seram tengan (ARMIX 93). 3 5 4 6 7 < 33.3 -100 Depth (m) >34.54 >34.54 -200 >34.54 -300 < 34.5 -400 Salinity (psu) >34.5 -500 -8.1 -8.0 -7.9 -7.8 -7.7 -7.6 -7.5 -7.4 -7.3 Latitude, S -6 -6.5 Latitude, S -7 -7.5 -8 -8.5 -9 120 7 6 5 4 3 2 1 8 FLORES SEA FLORES 120.5 121 121.5 122 Longitude, E Gambar 7. Distribusi tegak salinitas di Laut Flores, bulan Mei 2005 102 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah 0 WAIGEO Halmahera Sea HALMAHERA -1 37 38 39 PAPUA OBI MISOL Latitude, S -2 Seram Sea -3 SERAM -4 Banda Sea -5 127 128 129 130 131 132 Longitude, E Gambar 8. Distribusi tegak salinitas antara St.37 sampai St.39 pada program ARMIX 93 Salinitas 34,744psu s.d 34,688psu antara sigma-t ~25 s.d ~26 dengan suhu 15,15°C s.d 17,02 °C, merupakan massa air bagian utara Laut Halmahera sampai bagain utara (Line08) dengan salinitas antara 35,393 psu s.d 35,057 psu antara sigma-t ~22 - ~25 dengan suhu antara 23,54 s.d 23,71°C merupakan massa air dibagian utara Pulau Papua antara sigmat ~22 s.d ~26 dengan suhu antara 20,61°C s.d 20,76°C dan salinitas antara 35,459psu a/d 34,813psu; (5). Jenis massa air South Pacific Intermediate Water (SPIW) atau masa air dari Samudera Pasifik bagian selatan merupakan massa air Laut Halmahera dengan salinitas 34,538 psu dengan sigma-t ~ 27 dan suhu 6,45°C pada ratarata kedalaman 750m, merupakan masa air antara Pulau Halmahera dan Laut Seram bagian dibagian tengah (ARMIX 93) dengan salinitas antara 34,604 psu sampai 34,601 psu dalam sigma-t ~27 <27,5 dan suhu 5,27 s.d 6,08°C, merupakan masa air utara Pulau Papua (Line 09) dengan salinitas antara 34,471psu s.d 34,633psu dalam sigma-t (27 s.d <72,5) antara suhu 8,27°C s.d 8,48°C pada rata-rata kedalaman 750m; (6). Jenis massa air Antartic Intermediate Water (AAIW) untuk massa air Laut Halmahera dan Seram antara salinitas 34,167psu – 34,614 psu dalam sigma-t ~27,5 s.d <28 antara suhu 3,48°C s.d 3,28°C pada kedalaman >1500m, masa air Laut Halmahera bagian utara dan utara Pulau Papua bersalinitas antara 34,673 s.d 34,675psu dalam sigma-t ~28 antara suhue 1,57°C s.d 1,58°C pada kedalaman ~ 3200m (Gambar 9). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 103 Massa Air Subtropical Di Perairan Hamahera T-S Diagram Misol, Seram dan Banda bulan Maret - April 2007 35 L. Banda, Seram 30 75 m σ-t= 21 Sl. Halmahera, Misol, S 50 m 22 25 100 Temperature (°C) 23 20 24 SPSW 200 15 25 300 L.Band 10 500 26 SPIW 5 1000 27 28 0 33 33,5 34 Salinity (psu) 34,5 35 35,5 Gambar 9. Diagram T-S Laut Halmahera, Laut Seram (merah dan kuning) dan Laut Banda (biru) pada program EWIN 2007 Massa air bersalinitas maksimum antara 34,51psu s.d 34,981psu didapatkan pada sigma-t (24 s.d <27), dengan suhu antara 21,77 s.d 15,94°C dan kedalaman antara 150m s.d <300m. Kondisi ini sesuai dengan hasil analisa (Wyrtki 1961; Fine et al., 1994) bahwa pada lapisan pycnocline (100 to 300m) ada lapisan salinitas maksimum dari South Pacific. Kondisi inipun sesuai dengan (Ffield et al., 1992) bahwa South Pacific Central Water (SPCW) melewati Halmahera Sea (HS) memesuki South Banda (BS) and Timor Seas (TS). Kondisi ini sesuai dengan hasil (Ilahude and Gordon 1996) bahwa air bersalinitas dari asal South Pacific telah diobservasi dibawah thermocline di Laut Seram dan bagian selatan Laut Maluku. Inti (core) salinitas 34,98 psu di St.1 makin ke arah timur sudah tidak didapatkan lagi, bahkan inti salinitas >34,6 psu hanya diketemukan di St.15 pada kedalaman 200m. Kondisi ini menggambarkan bahwa salinitas maksimum tersebut tidak mengalir secara 104 kuat ke arah timur ke arah Laut Banda. Karena di St.24 sampai St.27 dan St.30 nilai salinitas 34,6psu sudah tidak diketemukan lagi atau tidak muncul. Kemungkinan salinitas maksimum mengalir ke arah barat ke Selat Obi dan ke barat laut Laut Seram. Arah transport salinitas maksimum tersebut adalah sesuai dengan (Liu, et al., 2005) bahwa air di atas South Pacific mengalir searah Halmahera eddy (HE) dan bergabung dengan arus ke arah timur dari Midanau Current (MC). Suatu jumlah yang kecil salinitas maksimum yang masuk Laut Seram dan Laut Banda dan kondisi demikian adalah sesuai dengan Ffiel et al. (2005). Hal ini pun sesuai dengan Gordon (2005) bahwa bersalinitas tinggi South Pacific Subtropical Water tidak menyebar kedalam lapisan thermocline di Laut Banda dari pintu gerbang Laut Halmahera. Setelah masuk perairan Indonesia bagian timur terjadi gradasi penurunan dari salinitas maksimum masa http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah air South Pacific Subtropical Water (SPSW) pada rata-rata kedalaman ~200m karena ada vertical mixing (Ffield, et al., 2005) yaitu dimulai dari salinitas maksimum 35,459psu bagian pantai utara Pulau Papua, sampai di mulut Halmahera menjadi 35,394psu, sampai di Laut Halmahera bagian selatan menjadi 34,987psu , sampai di Halmahera bagian barat antara Pulau Halmahera dan Pulau Obi menjadi 34,744psu, sampai Laut Seram bagian tengah menjadi 34,721psu, sampai di Laut Banda menjadi 34,663 psu. Sedangkan North Pacific Subtropical Water (NPSW) pada ratarata kedalaman ~150m yaitu di mulai dari salinitas 35,057psu di utara Halmahera, sampai di Selat Makassar menjadi 34,863psu, sampai di Laut Flores menjadi 34,567psu dan sampai di Selat Lombok menjadi 34,555psu. Sebaliknya salinitas minimum massa air North Pacific Intermediate Water (NPIW) justru ada kenaikan setelah bercampur dengan massa air laut dalam perairan Indonesia, dimulai dari Laut Hamahera bagian utara salinitas minimum pada rata-rata kedalaman ~400m yaitu salinitas 34,395psu, sampai di Selat Makassar menjadi 34,481 psu (Anonimus, 2003 and 2004), sampai di Laut Flores menjadi 34.464 psu (Anonimus, 2005) dan sampai di Selat Lombok menjadi 34.464 psu (Instant, 2003). Demikian untuk salinitas minimum massa air South Pacific Intermediate Water (SPIW) rata-rata pada kedalaman ~750m, dimulai dari pantai utara Pulau Papua yaitu salinitas 34.537psu, sampai di mulut Laut Halmahera salinitas menjadi 34.534 psu dan Halmahera bagian selatan 34.538 psu, sampai di Selat Obi menjadi 34.604 psu dan Seram 34.535 psu, sampai di Laut Banda Seram bagian timur menjadi 34.547 psu. Penurunan salinitas maksimum masa air NPSW pada kedalaman ~120m s.d 160m yaitu dimulai dari pantai utara Pulau Papua yang bersalinitas 35.576 psu dan sampai di mulut Halmahera menjadi 35.394 psu. Selajutnya salinitas masimum tersebut masuk ke perairan Indonesia bagian timur di Laut Halmahera bagian selatan menjadi 34.987 psu, sampai di Halmahera bagian barat antara Pulau Halmahera dan Pulau Obi menjadi 34.744 psu, sampai Laut Seram bagian tengah menjadi 34.721 psu, sampai di Laut Banda menjadi 34.663 psu. Sedangkan salinitas maksimum masa air NPSW pada rata-rata kedalaman ~150 m yaitu di mulai dari salinitas maksimum 35.057 psu di utara Halmahera, sampai di Selat Makassar menjadi 34.863 psu, sampai di Laut Flores menjadi 34.567 psu dan sampai di Selat Lombok menjadi 34.555 psu. Sebaliknya salinitas minimum massa air NPIW justru didapatkan kenaikan setelah bercampur dengan massa air laut dalam perairan Indonesia, dimulai dari Laut Hamahera bagian utara salinitas minimum pada rata-rata kedalaman ~400m yaitu salinitas 34,395psu, sampai di Selat Makassar menjadi 34,481psu, sampai di Laut Flores menjadi 34.464psu dan sampai di Selat Lombok menjadi 34,464psu. Demikian untuk salinitas minimum massa air SPIW rata-rata pada kedalaman ~750m, dimulai dari pantai utara Pulau Papua (34,537psu), sampai di mulut Laut Halmahera salinitas menjadi 34,534psu dan Halmahera bagian selatan 34,538psu, sampai di Selat Obi menjadi 34,604psu dan Seram 34,535 psu, sampai di Laut Banda Seram bagian timur menjadi 34,547psu (Gambar 10). Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 105 Massa Air Subtropical Di Perairan Hamahera 35 30 30 Seram - Banda Halmahera 50 m 14 38 Sigma_t = 21 22 39 30 45 25 Temperature (°C) 100 ♦ Flores St_6 24 ▲ Flores St_8 ▲ Halmahera St_1 ● Seram St_8 – Seram St_15 25 ♦ Banda St_21 ▲Banda St_30 ж Armix St_37 26 + Armix St_38 - Armix St_39 ■ Armix St_45 ▲Papua St_105 ♦ Halmahera St_27 ▲Halmahera St_14 20 15 10 5 27 6 23 25 NPSW 21 8 20 SPSW BANDA FLORES 105 1 8 SERAM 15 150 SPSW 15 HALMAHERA 200 10 300 NPIW 500 700 27 SPIW 5 28 1000 4700 AAIW 0 0 33 33.2 33.4 33.6 33.8 34 34.2 34.4 34.6 34.8 35 35.2 35.4 35.6 35.8 36 Salinity (psu) Gambar 10. Diagram T-S bagian selatan Laut Halmahera, Laut Seram dan Laut Banda (2007), Laut Floras (2005), Selat Obi (ARMIX 1993). Bagian utara Pulau Halmahera dan Pulau Papua pada Expedisi WOCE IV. KESIMPULAN Arus bagian permukaan dan bagian dalam lebih dominan bergerak ke arah selatan sampai barat daya, ini menggambarkan bahwa arus tersebut adalah arus ARLINDO yaitu dari massa air Samudera Pasifik bagian selatan melalui Laut Halmahera. Kedalaman lapisan permukaan perairan Halmahera, Seram dan Banda didapatkan berkisar antara 18m s.d 74m. Kedalaman lapisan batas atas termoklin berkisar antara 25m s.d 96m dengan rata-rata 70m, sedangkan kedalaman termoklin batas bawah berkisar antara 75m - 267m dengan rata-rata kedalaman 180m. Suhu di lapisan permukaan (mixed layer) di Laut Halmahera didapatkan lebih tinggi dibandingkan suhu Laut Seram dan Laut Banda, demikian pula di lapisan termoklin pada kedalaman antara 106 225m – 385m. Pada kedalaman antara 545m s.d 612m didapatkan mempunyai nilai salinitas maksimum. Perairan yang diobservasi ternyata didapatkan ada dua massa air yang berbeda dari permukaan sampai kedalaman ~700m, yaitu masa air besalintas rendah di Laut Banda dekat dengan Seram bagian timur dan salinitas tinggi di Selat Halmahera sampai Laut Seram di bagian barat. Massa air bersalinitas maksimum (34,988psu) antara densitas <25 s.d >26 didapatkan pada kedalaman 236m terdapat pada lapisan termoklin, dan mengecil ke arah barat dan timur Laut Seram sampai Laut Banda. Hasil analisa diagram T-S didapatkan ada enam jenis masa air yaitu massa air Selat Halmahera dan perairan Misol (HMW); massa air Laut Seram dan Laut Banda (SBW); masa air Laut Banda (BW); masa air South Pacific Subtropical Water (SPSW); massa air South Pacific http://www.itk.fpik.ipb.ac.id/ej_itkt22 Hadikusumah Intermediate Water (SPIW); massa air Antartic Intermediate Water (AAIW). UCAPAN TERIMAKASIH Penulis mengucapkan banyak terimakasih kepada rekan peneliti dan awak kapal KR. Baruna Jaya VII. Penulis menyampaikan penghargaan yang tinggi yaitu kepada mereka, Prof Dr Lukman Hakim, sebagai Ketua LIPI, Dr Herry Harjono, sebagai Deputi Bidang IPK LIPI, Dr Suharsono sebagai Ketua Pusat Penelitian Oseanografi - LIPI untuk saran dan dukungan kebijakan penerapan program. Terima kasih dan penghargaan kepada Fredy Letemia sebagai Koordinator Program EWIN di Ambon. DAFTAR PUSTAKA Anonimus. 2003. Laporan akhir studi dinamika Selat Makassar serta interaksinya dengan daratan PULAU Kalimantan dan PULAU Sulawesi. Program kompetitif Kalimantan Timur dan Bangka – Belitung – LIPI. 105 hal. Anonimus. 2004. Laporan akhir studi dinamika Selat Makassar serta interaksinya dengan daratan PULAU Kalimantan dan PULAU Sulawesi. Program kompetitif Kalimantan Timur dan Bangka – Belitung - LIPI. 146 hal. Anonimus. 2005. Laporan akhir Penelitian Arlindo dan Efeknya pada Stratifikasi Massa Air Laut Flores dan Sekitarnya. P2O - LIPI. 36 hal. Birowo. 1990. Introduced the scientific knowledge of circulation in Indonesia Seas. In. Intergovernmental Oceanographic Commission Workshop Report No. 72. Ffield, A. 1994. Tidal mixing in Indonesian Seas. Paper presented at International Scientific Symp. of the IOC-WESTPAC, Intergov. Oceanogr. Comm. Bali. Indonesia. Ffield, A. and A.L. Gordon, 1992. Vertical mixing in the Indonesian thermocline, J. Phys. Oceanogr., 22:184-195. Ffield, A. and R. Robertson. 2005. Indonesian seas fine structure variability. Oceanography., 18(4):108–111. Fine, R.A., R. Lukas, F.Bingham, M. Warner, and R. Gammon. The western equatorial Pacific: A water masses crossroads, J. Geophys. Res., 99:25,063-25,080 Gordon, A.L. 2005. Oceanography of the Indonesian Seas and their trough flow. Oceanography, 18(4):14–27. Hadikusumah. 2008. Masa Air Subtropikal dan Intermediate Melalui Selat Makassar. J. Segara, 4(2):111-120 Hadikusumah. 2009. The Circulation of Raja Ampat Waters – Papua with Pacific intrusion to the East Indonesian Waters. Jurnal Ichthyos, 8(1):27-34. Ilahude, A.G. and A.L. Gordon. 1996. Thermocline Stratification Within the Indonesian seas. Journal of Geophysical Research, 101(C5):12,401–12,409. Liu Hailong, Li Wei, Yu Yongqiang, and Zhang Xuehong. 2005: Climatology and variability of the Indonesian throughflow in an Eddy-permitting Oceanic GCM, Adv. Atmos. Sci., 22(4):496-508. Tomczak, M. and J. S. Godfrey. 1994. Regional Oceanography: An Introduction. Pergamon. Wyrtki, K. 1956. The subtropical lower water between the Philippines and New Guinea. Mar. Res. Indonesia, 1:21-45. Jurnal Ilmu dan Teknologi Kelautan Tropis, Vol. 2, No. 2, Desember 2010 107 Massa Air Subtropical Di Perairan Hamahera Wyrtki, K. 1961. Physical oceanography of the Southeast Asian Waters. Scientific results of Marine investigations of the South China Sea and the Gulf of Thailand 19591961. The University of California, Scripps Institution of Oceanography La Jolla, California: 195 pp. Wyrtki, K. 1962. Physical oceanography of the Southeast Asian waters, Naga Report, Vol.2. Scripps Institution of Oceanography, California:195 pp. Yuji, K., M., Aoyoma, T., Kawano, N. Hendiarti, Syaefudin, Y., Anantasena, K. Muneyama, and H., Watanabe. 1996. The water masses between Mindanao and New Guinea. J. Geophys. Res., 101(C5):12,391-12,400. 108 http://www.itk.fpik.ipb.ac.id/ej_itkt22 Pedoman Penulisan Naskah Artikel 1. Ketentuan Umum Artikel merupakan hasil penelitian atau sanggahan (note) ilmiah bidang ilmu dan teknologi kelautan tropis dan belum pernah dimuat maupun dalam proses pengajuan dalam publikasi ilmiah lain. Artikel yang diusulkan dapat berasal dari bidang ilmu dan teknologi kelautan tropis sebagai berikut: biologi laut, ekologi laut, biologi oseanografi, kimia oseanografi, fisika oseanografi, geologi oseanografi, dinamika oseanografi, coral reef ecology, akustik kelautan, remote sensing kelautan, sistem informasi geografis kelautan, mikrobiologi kelautan, pencemaran laut, akuakultur kelautan, teknologi hasil perikanan, bioteknologi kelautan, air-sea interaction, dan ocean engineering. Artikel ditulis dalam bahasa Indonesia atau bahasa Inggris diketik dengan MSWord, font Times New Roman ukuran 12 pada kertas kuarto atau A4 termasuk Gambar dan Tabel dengan margin top and bottom 3 cm serta left and right 3cm. Untuk artikel dalam bahasa Indonesia, tulisan dilengkapi dengan abstract (bahasa Inggris) dan abstrak (bahasa Indonesia). Sedangkan artikel dalam bahasa Inggris, tulisan hanya menyertakan abstract (bahasa Inggris). Semua komunikasi dengan penerbit dilakukan secara electronic (email). Naskal artikel harap dikirim ke Pemimpin Redaksi Jurnal Ilmu dan Teknologi Kelautan Tropis dengan email address: [email protected]. Semua naskah yang masuk akan mendapat balasan penerimaan. Hasil review dari reviewers (mitra bebestari) juga akan dikirim via email. 2. Sistematika Susunan Artikel 2.1. Sistematika susunan artikel hasil penelitian umumnya sebagai berikut: Judul (sesingkat mungkin) dan disertai dengan terjemahan dalam bahasa Inggris yang diketik secara miring (italic). Nama Penulis, nama dan alamat instansi, dan e-mail corresponding author. Abstract dalam Bahasa Inggris (memuat sedikit latar belakang, tujuan, metode, dan hasil penelitian serta tidak lebih dari 250 kata. Semua ditulis dalam Bahasa Inggris dengan cetak miring) Artikel dalam bahasa Indonesia (memuat sedikit latar belakang, tujuan, metode, dan hasil penelitian serta tidak lebih dari 250 kata). Keywords maximum 8 words (English) Kata kunci maksimal 8 kata (Bahasa Indonesia) Pendahuluan (tanpa sub judul, memuat latar belakang, masalah, rumusan masalah, rangkuman kajian teoretik, ulasan ilmiah terkait judul berdasarkan rujukan (pustaka) terkini, dan tujuan penelitian). Dalam pendahuluan ini juga disajikan pertanyaan ilmiah (scientific question) yang akan dijawab dalam penelitian tersebut. Metode Penelitian (ditulis dengan jelas waktu, lokasi, bahan, dan analisis data penelitian sehingga memungkinkan peneliti lain untuk mengulangi percobaan yang 109 terkait). Bagian ini dapat dibuat dalam beberapa sub-bab. Hasil dan Pembahasan (sajikan hasil terlebih dahulu kemudian diikuti dengan penjelasan atau pembahasan. Pembahasan harus menggunakan rujukan atau dibandingkan (diulas) dengan rujukan (pustaka) terkini). Kesimpulan dan Saran (ditulis dalam bentuk essay (paragraph) secara ringkas dan jelas dan harus bisa menjawab (menjelaskan) judul, tujuan, dan hasil penelitian). Ucapan Terima Kasih (ditulis dengan jelas dan ringkas kepada siapa ucapan terima kasih itu diberikan. Penelitian yang dibiayai DIPA, hibah, atau sejenisnya agar mencantumkan nomor kontraknya). Daftar Pustaka (lihat ketentuan berikutnya) Lampiran (jika ada) 2.2. Sistematika susunan sanggahan (note) ilmiah umumnya adalah sebagai berikut: Judul (sesingkat mungkin) dan disertai dengan terjemahan berbahasa Inggris yang ditulis dalam bentuk miring (italic). Nama penulis, nama dan alamat instansi, dan e-mail address corresponding author. Pendahuluan (tanpa sub judul, memuat pengantar topik utama diakhiri dengan rumusan tentang hal-hal pokok yang akan dibahas) Pembahasan diikuti dengan Subjudul sesuai kebutuhan Kesimpulan (bila perlu) Daftar Pustaka Lampiran (jika ada) 3. Teknik Penulisan 3.1. Judul Judul ditulis dengan huruf kapital, dicetak tebal, di tengah (center), font Times New Roman 12, hitam. Di bawah judul naskah dalam bahasa Indonesia, diberikan terjemahan judul dalam bahasa Inggris dengan huruf miring (italic). Contoh: STUDI INTERAKSI PADA HUMIN UNTUK ADSORPSI Mg (II) DAN Cd (II) DALAM MEDIUM AIR LAUT STUDY OF INTERACTION ON HUMIN FOR Mg (II) AND Cd (II) IN THE SEA WATER MEDIUM 3.2. Nama Penulis Nama penulis ditulis tanpa gelar dengan huruf kapital pada awal nama, dicetak tebal, di tengah, font Times New Roman 12, hitam. Dilengkapi dengan nama dan alamat instansi dan e-mail untuk corresponding author dengan font Times New Roman 12, hitam dengan spasi 1. Contoh: 110 Tuti Wahyuni Pusat Riset Teknologi Kelautan-BRKP-DKP e-mail: [email protected] Jika artikel ditulis lebih dari satu orang dan alamat instansinya berbeda maka disetiap nama penulis diikuti dengan nomor yang ditulis secara superscript Email address yang dicantumkan hanya utk corresponding author saja. Contoh 1: Contoh 2: Tuti Wahyuni1 dan Dendy Mahabror2 1 Pusat Riset Teknologi Kelautan-BRKP-DKP e-mail: [email protected] 2 Balai Riset Observasi dan Kelautan-BRKP-DKP Tuti Wahyuni1, Dendy Mahabror2, dan Rani Ulawi3 1 Pusat Riset Teknologi Kelautan-BRKP-DKP e-mail: [email protected] 2 Balai Riset Observasi dan Kelautan-BRKP-DKP 3 Departemen Ilmu dan Teknologi Kelautan, FPIK-IPB 3.3. Abstrak Berbahasa Inggris dan Keywords Tulisan “Abstract” ditulis dengan huruf kapital, tengah (center) dicetak tebalmiring (italic), font Times New Roman 12, hitam, spasi 1. Isinya tidak dicetak tebal. Penulisan rata kiri dan kanan, tanpa alinea (abstract keseluruhan merupakan satu alinea). Tulisan “Keywords” ditulis dengan huruf kapital di awal kata, dicetak tebal, font Times New Roman 12, hitam, diberi titik dua, selanjutnya tidak dicetak tebal. Penulisannya rata kiri. Contoh: ABSTRACT A study of interaction on humin for Mg(II) and Cd(II) in the sea water medium was investigated... ... and so on. Keywords: Adsorption, Humin, Magnesium, Cadmium 3.4. Abstrak Berbahasa Indonesia dan Kata Kunci Tulisan “Abstrak” ditulis dengan huruf kapital, tengah (center) dicetak tebal, font Times New Roman 12, hitam, spasi 1. Isinya tidak dicetak tebal. Penulisan rata kiri dan kanan, tanpa alinea (abstrak keseluruhan merupakan satu alinea). Tulisan “Kata kunci” ditulis dengan huruf kapital di awal kata, dicetak tebal, font Times New Roman 12, hitam, diberi titikdua, selanjutnya tidak dicetak tebal. Penulisannya rata kiri. Contoh: 111 ABSTRAK Penelitian tentang studi interkasi pada humin untuk absorpsi Mg(II) dan Cd(II) dalam medium air laut .... dan seterusnya. Kata kunci: Adsorpsi, Humin, Magnesium, Kadmium 3.6. Bab (Chapter) dan Sub-Bab (Sub-Chapter) Bab (Chapter) ditulis dengan urutan angka romawi, huruf kapital, dicetak tebal, rata tepi kiri, font Times New Roman 12, hitam sedangkan sub-bab (sub-chapter) ditulis dengan urutan angka biasa, huruf kapital di awal kata, dicetak tebal, rata tepi kiri, font Times New Roman 12, hitam. Apabila di bagian sub-bab masih ada subnya lagi, maka penulisannya diberi nomor paralel dengan sub-bab sebelumnya diikuti titik, judul dengan huruf kapital di awal kata, cetak tebal, rata tepi kiri, font Times New Roman 12, hitam. Contohnya berikut ini: --------------------------------------------------------------I. PENDAHULUAN II. 2.1. 2.2. 2.3. METODE PENELITIAN Waktu dan Tempat Penelitian Bahan dan Data Analisis Data III. HASIL DAN PEMBAHASAN 3.1. Hasil 3.1.1. Suhu Permukaan 3.1.2. Konsentrasi Klorofil-a 3.2. Pembahasan IV. KESIMPULAN ----------------------------------------------------------------3.8. Catatan Kaki (Footnote) Catatan kaki diberi simbol angka setelah frase/istilah(1) yang akan diterangkan. Catatan kaki yang merupakan keterangan kata/frase ditulis dengan font Times New Roman 8, hitam. 3.9. Tabel Judul tabel diletakkan di atas tabel. Judul tabel ditulis dengan huruf kapital di awal kata, diletakkan di tengah (center), font Times New Roman 12. Tabel diberi nomor, diikuti titik, kemudian judul tabel (misalnya Tabel 1. Judul…, Tabel 2. Judul…). Jarak peletakan table dari kalimat diatasnya sebanyak 2 spasi dan jarak tabel ke kalimat baru dibawahnya sebanyak 2 spasi. Jarak dari judul tabel terhadap tabel itu sendiri sebanyak 1 spasi. Kalau ada catatan kaki untuk tabel tersebut maka jaraknya dari table ádalah 1 spasi. Bila lebih dari satu baris menggunakan spasi 1. Contoh: 112 Tabel 1. Kandungan Humin dan Asam Humat Hasil Isolasi Tanah Gambut ctional Group Content (cmole/kg) Humin (1) Humin (2) Total Acidity 677 543 -COOH 115 199 -OH Phenolic 562 344 1= Isolated Peat Soil from Siantan Hulu, West Kalimantan. 2= Isolated Peat Soil from Siantan Hulu, West Kalimantan (Saleh, 2004) 3.10. Gambar Gambar dapat berupa diagram, grafik, peta, foto (yang mengemukakan data) dan lain-lain. Judul gambar diletakkan di bawah gambar, ditulis dengan huruf kapital di awal kata, diletakkan di tengah (center), font Times New Roman 12. Jarak dari judul gambar terhadap gambar itu sendiri sebanyak 1 spasi. Kalau ada catatan kaki untuk gambar tersebut maka jaraknya dari table ádalah 1 spasi. Gambar diberi nomor diikuti titik, kemudian judul gambar (misal Gambar 1. Judul..., Gambar 2. Judul ...). Bila judul lebih dari dua baris menggunakan spasi 1. Contoh penulisan sebagai berikut: Gambar 1. Dermaga Tetap 3.11. Rujukan dan Daftar Pustaka Teknik penulisan rujukan dalam teks dan daftar pustaka, menggunakan gaya yang umum dipakai dalam pedoman penulisan ilmiah khususnya dalam International Journal. Daftar pustaka hanya mencantumkan sumber yang dirujuk dalam teks saja. Sebaliknya, referensi yang dirujuk dalam teks harus dicantumkan dalam daftar pustaka. Daftar pustaka diurutkan secara alfabetis, menggunakan font Times New Roman 12, hitam. Bilamana referensinya lebih dari satu maka diurutkan berdasarkan tahun terbit yang paling baru. Cara menuliskan sumber pustaka (rujukan) adalah sebagai berikut. Menulis Rujukan dalam Teks Untuk penulisan rujukan, tulis nama keluarga dari pengarang diikuti koma atau titik dan tahun terbit artikel/paper/laporan/prosiding/dll. Untuk pengarang lebih dari 2 orang dituliskan dengan menggunakan “et al.” (ditulis miring). Penulisan “dan” atau ”and” sebelum nama terakhir ditulis sesuai dengan judul tulisan tersebut yaitu ”dan” untuk Indonesia dan ”and” untuk Inggris. Contoh: Anastasi (1997) menyatakan ..... atau .....(Anastasi, 1997). Kiswara dan Winardi (1994) menyimpulkan ..... atau ..... (Kiswara dan Winardi, 1994). 113 Berk and Romly (1984) meneliti .... atau ...... (Berk and Romly, 1984). Ali et al. (2008) menjelaskan....atau...... (Ali et al., 2008). Menulis Daftar Pustaka Tulis nama keluarga diikuti koma, satu spasi jarak, singkatan nama pertama atau kedua (bila ada) diikuti titik, dua spasi jarak, tahun terbit diikuti dengan titik, dua spasi jarak, Judul artikel/paper, nama jurnal (ditulis dengan miring) diikuti titik, volume(edisi), titik dua, nomor halaman paper/artikel dalam jurnal. Bila lebih dari satu baris, maka baris selanjutnya masuk dengan 9 ketukan (1,25 cm hanging left). Contoh: Kiswara, W. dan L. Winardi. 1994. Keanekaragaman dan sebaran lamun di Teluk Kuta dan Teluk Gerupuk, Lombok Selatan. Jurnal Teknologi Kelautan Nasional: 3(1):23-36. Mardi, L.M., T.M. Nathan, R.A. Raman, and W.L. Joran. 2008. Fish stock assessment in Java Sea. J. Marine Science, 3(2):123-145. Buku dengan seorang penulis atau lebih Anastasi, A. 1997. Psychological Testing. 4th ed. MacMillan Press. New York. 234p. Berk, R.A., B.A. Romly, and N.N. Siogu. 1984. A Guide Criterion Referenced Test Construction. The John Hopkins University Press. Baltimore. 389p. Artikel dalam sebuah buku/prosiding Artikel yang terdapat dalam sebuah buku atau prosiding, ditulis dengan menulis artikel itu terlebih dahulu lalu diikuti dengan buku atau prosiding tersebut. Contoh: Berk, R.A.1988. Selecting index or realibility. In: R.A. Berk (ed.), A Guide to Criterion-Referenced Test Construction. The John Hopkins University Press. Baltimore. 200-217pp. Ramdi, N.S., B.K. Roland, and D. Torres. 2010. Variabilitas konsentrasi klorofil-a di Laut Jawa. Dalam: Nababan et al. (ed.). Prosiding Pertemuan Ilmiah Tahunan VI ISOI 2009, International Convention Center, Botani Square, Bogor, 16-17 November 2009. Hal.: 223-247. Dua artikel dalam sebuah buku Ditulis dengan cara seperti yang telah diuraikan, dengan tambahan huruf a, b, c, dan seterusnya, yang ditempatkan dibelakang tahun terbit. Contoh: Berk, R. A. 1984a. Selecting index or realibility. In: R.A. Berk (ed). A Guide to Criterion-Referenced Test Construction. The John Hopkins University Press. Baltimore, 234-345p. 114 Berk, R. A 1984b. Conducting the item analysis. In: R.A. Berk (ed). A Guide to Criterion-Referenced Test Construction. The John Hopkins University Press. Baltimore. 123-134p. Terjemahan Cara penulisannya sama dengan cara menulis pustaka lain, kecuali judul buku diganti dengan judul yang sudah diterjemahkan. Di belakang judul tersebut dituliskan nama penerjemah, yang diawali dengan nama kecilnya, yang di belakang dituliskan kata penterjemah. Contoh: Gagne, R.M., L.J. Briggs, and W.W. Wage. 1988. Prinsip-prinsip Desain Instruksional, (3rd Ed.). Soeparman, K. (penterjemah). Holt, Rineahart, and Winston press. Chicago. Artikel dari Internet Artikel yang diambil dari internet (website) maka cara menulisakannya sebagai berikut: nama pengarang artikel, tahun terbitan, judul artikel, nama majalah/ artikel/jurnal/yang lain, alamat website, tanggal akses. Contoh: Lynch, T. 1996. DS9 Trials and Tribble – Actions Review. From Psi Phi:Bradley’s Science Fiction Club, http://www.bradley.edu/compusorg/psiphi/DS9/ep/SO3r.htm. Retrieved on 23 March 2007. Artikel pada surat kabar Surat kabar atau artikel dalam surat kabar, pada umumnya dicantumkan dalam daftar pustaka dengan menulis nama penulis, tahun terbit, judul artikel, nama surat kabar dan tanggal terbit. Contoh: Nababan, B. 2009. Laut Bukan Lagi Penyerap Carbon. Antara, 12 Mei 2009. Artikel yang tidak dipublikasikan Skripsi, tesis, dan disertasi dapat digolongkan ke dalam materi yang tidak dipublikasikan. Contoh: Nababan, B. 2005. Bio-optical Variability of Surface Waters in the Northeastern Gulf of Mexico. Dissertation. College of Marine Science. University of South Florida. 158p. Buku/Laporan Hasil Penelitian Tanpa Pengarang Buku atau laporan hasil penelitian yang merupakan hasil penelitian dari institusi atau lembaga ditulis dengan menyebutkan nama institusi atau lembaga yang menerbitkan buku atau laporan hasil penelitian tersebut, diikuti tahun penerbitan, judul buku, penerbit (institusi penerbit), dan diikuti jumlah halaman. 115 Contoh: Kementerian Pendidikan Nasional. 1985. Kurikulum Sekolah Menegah Pertama (SMP). Kementerian Pendidikan Nasional. Jakarta. 219hal. Pusat Penelitian Oseanografi Lembaga Ilmu Pengetahuan Indonesia (P2O-LIPI). 2008. Prospek Perikanan Indonesia. P2O-LIPI, Jakarta. 234hal. 116