II KAJIAN KEPUSTAKAAN 2.1 Itik Cihateup Klasifikasi itik lokal
advertisement

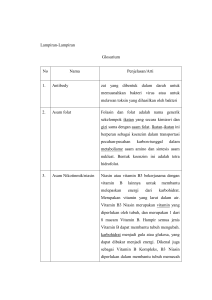
II KAJIAN KEPUSTAKAAN 2.1 Itik Cihateup Klasifikasi itik lokal menurut Campbell dan Lack (1985) adalah sebagai berikut : Kingdom Subkingdom Filum Subfilum Kelas Ordo Famili Genus Spesies : Animalia : Metazoa : Chordata : Vertebrata : Aves : Anseriformes : Anatidae : Anas : Anas platyrhyncos Itik Cihateup berasal dari Desa Cihateup, Kecamatan Rajapolah, Kabupaten Tasikmalaya, Jawa Barat, hidup dan beradaptasi pada daerah ketinggian 378 m dpl (Wulandari dkk., 2005; Matitaputty, 2012) dan sering disebut dengan itik gunung. Itik Cihateup penyebarannya sudah sampai di daerah Kabupaten Garut. Selain itik Cihateup, ada juga itik Cirebon yang juga berasal dari Jawa Barat dan memiliki ciri-ciri yang hampir sama dengan itik Cihateup (Wahid, 2003). Beberapa hasil penelitian secara deskriptif menjelaskan bahwa yang menjadi ciri khas itik Cihateup dari itik lainnya termasuk itik Alabio dan Mojosari adalah ukuran panjang leher, sayap yang lebih panjang. Perbedaan ukuran tubuh tersebut diduga karena adanya pengaruh lingkungan pemeliharaan di kawasan pengunungan (Suryana dan Matitaputty, 2014). Itik jantan dan betina memiliki warna hitam di bagian paruh dan kaki, leher yang lebih panjang dibandingkan itik lokal lainnya (Mito dan Johan, 2011). Warna paruh dan shank 8 yang hitam pada dipengaruhi oleh adanya gen dermal melanin dan mirip dengan itik lainnya yang ada di Jawa seperti itik Tegal dan Mojosari. (Wulandari dkk., 2005). Penelitian karakteristik corak bulu, warna paruh dan shank pada itik Cihateup dilaporkan Wulandari dkk. (2005) dan dijelaskan bahwa warna bulu bagian leher itik Cihateup jantan didominasi warna penciled dan ekor warna polos, sedangkan paruh dan shank didominasi warna hitam. Itik betina warna bulu bagian leher, dada, shank dan ekor sedikit berbeda dengan jantan yakni warna laced dan buttercup, sementara pada shank dan paruh tetap didominasi warna hitam. 2.2 Aspek Metabolisme sebagai Dampak Pemeliharaan Minim Air Itik memiliki kebiasaan berenang di kolam air ataupun di persawahan. Sistem pemeliharaan tradisional di mana itik digembalakan di sawah atau di tempat-tempat yang banyak air, namun dengan cepat mengarah pada pemeliharaan secara intensif yang sepenuhnya terkurung, akibatnya kebiasaan berenang dan membasahi tubuhnya tidak dapat dilakukan oleh itik, ditambah lagi dengan suhu lingkungan yang tinggi membuat itik mengalami cekaman panas. Cekaman panas menginduksi suatu seri reaksi kaskade pada sistem saraf dan endokrin sehingga terjadi peningkatan aktivitas jalur hipotalamus-hipofisa-korteks adrenal yang dikenal sebagai jalur hipotalamus, hipofisa, adrenal. Hipotalamus mensekresikan corticotropine releasing factor (CRF), selanjutnya mengaktifkan adrenocorticotropic hormone (ACTH) dari hipofisa anterior yang akhirnya merangsang sekresi glukokortikoid dari korteks adrenal (Ladd dkk., 1996). Hormon ini bekerja dengan meningkatkan glukoneogenesis melalui peningkatan katabolisme asam amino di hati akibat induksi pada aminotransferase (dan enzim 9 lain, misalnya triptofan dioksigenase) serta enzim-enzim penting pada glukoneogenesis (Bender, 2009; Botham dan Mayes, 2009). Ilustrasi 1. Skema Hipotalamus-Pituitary-Adrenal (HPA) Axis, menjelaskan regulasi dan negative feedback pada kortisol melalui reseptor glukokortikoid. Keterangan : GRs : Glucocorticoid Receptors MRs : Mineralcorticoid Receptors CRF : Corticotropine Releasing Factor ACTH : Adrenocorticotropic Hormone AVP : Vasopressin Glukneogenesis merupakan sintesis glukosa dari senyawa non karbohidrat dan terjadi di dalam hati. Substansi glukogenik utama adalah asam amino dapat dikonversikan menjadi α-ketogacids oleh deaminasi dan reaksi transaminasi. Beberapa ketoacids (piruvat, oksaloasetat, α-ketoglutarat) dikonversi menjadi 10 glukosa dan glikogen. Senyawa bebas N yang berasal dari asam amino oleh deaminasi dapat juga dikonversi menjadi salah satu α-ketoacids dan bertindak sebagai prekursor karbohidrat (Allen, 1970). Tiga urutan reaksi pada glukoneogenesis yang berbeda pada proses glikolisis melibatkan perubahan piruvat menjadi fosfoenolpiruvat (PEP) dan reaksi ini mengeluarkan fosfat dari fruktosa 1,6-bifosfat untuk membentuk fruktosa 6-fosfat dan dari glukosa 6- fosfat untuk membentuk glukosa. Serangkaian enzim mengkatalis perubahan piruvat menjadi fosfoenolpiruvat. Reaksi yang mengeluarkan fosfat dari fruktosa 1,6 bifosfat dan dari glukosa 6fosfat masing-masing menggunakan enzim yang berbeda dengan enzim padanan pada glikolisis. Fosfat dikeluarkan oleh fosfatase yang membebaskan Pi selama glukoneogenesis. Prekursor glukoneogenesis adalah asam amino, laktat, dan gliserol. Reaksi glukoneogenesis menghasilkan ATP (King, 2010). 2.3 Buah Makasar Buah makasar mempunyai rasa yang sangat pahit dan banyak mengandung senyawa qassinoid. Senyawa tersebut dalam buah makasar sudah dilakukan penelitian sebagai anti tumor (Lee dkk., 1984; Fukamiya, dkk., 1992; Rahma dkk., 1999). Penggunaan minyak esensial tidak hanya mempengaruhi populasi mikroba, tetapi pada saat yang sama berpengaruh positif terhadap aktivitas enzim pencernaan dan intermediate metabolism. 11 Ilustrasi 2. Buah Makasar (Brucea javanica (L.) Merr.) Minyak buah makasar menggunakan GC-MS menunjukkan bahwa terdapat dua kelompok senyawa di dalamnya yaitu asam lemak dan senyawa organik lainnya. Asam lemak yang paling banyak terkandung dalam minyak buah makasar adalah asam linoleat (CH3(CH2)16COOH), yaitu sebesar 52,89%. Asam linoleat atau dikenal dengan istilah asam lemak omega 6 adalah asam lemak yang memiliki rantai karbon sebanyak 18 dan mengandung dua ikatan rangkap pada posisi 9 (C9-C10) dan 12 (C12-C13) dengan isomer geometris cis. Asam lemak tak jenuh jamak (Poly Unsaturated Fatty Acid/PUFA) adalah asam lemak yang mengandung dua atau lebih ikatan rangkap, bersifat cair pada suhu kamar bahkan tetap cair pada suhu dingin, karena titik lelehnya lebih rendah dibandingkan dengan MUFA atau SFA. Long Chain Poly Unsaturated Fatty Acid (LCPUFAs) disintesisis dari asam linoleat, seperti dihomo-gamma-linolenic acid (DGLA) dan asam arakhidonat, dimana mereka merupakan prekursor eicosanoid (Bergstrom dkk., 1964; Van Dorp dkk., 1964). Eicosanoid merupakan susbtansi messenger yang sangat berpengaruh terhadap regulasi berbagai macam proses termasuk sekresi asam lambung, kontraksi uterus, reproduksi, tekanan darah dan inflammasi (Samuelsson, 1981). 12 Asam lemak esensial merupakan prekursor sekelompok senyawa eicosanoid yang mirip hormon, yaitu prostaglandin, prostasiklin, tromboksan dan leukotrien. Senyawa-senyawa ini mengatur tekanan darah, denyut jantung, fungsi kekebalan, rangsangan sistem saraf, kontraksi otot serta penyembuhan luka (Murray dkk., 2009). 2.4 Asam Urat Darah Asam urat merupakan hasil metabolisme normal dari pencernaan protein (terutama dari daging, hati, ginjal, dan beberapa jenis sayuran seperti kacang dan buncis) atau dari penguraian senyawa purin yang seharusnya akan dibuang melalui ginjal, feses, atau keringan (Sustrani, 2004). Ilustrasi 3. Struktur Kimia Asam Urat Proses pembentukan asam urat sebagian besar di peroleh dari metabolisme nukleotida purin endogen, guanosine monophosphate (GMP), inosine monophosphate (IMP) dan adenosine monophosphate (AMP). Enzim xantine oxidase mengkatalis hipoxantine dan guanine dengan produk akhir asam urat (Mustafiza, 2010). 13 Asam nukleat (di makan dalam bentuk nukleoprotein dan dari penghancuran sel-sel tubuh) Enzim proteolitik......di usus Asam nukleat Nuklease (DNAse dan RNAse)......di getah pankreas Nukleotida Polinukleotidase = fosfoesterase......di usus Mononukleotida Nukleotidase dan fosfatase Nukleosida Fosforilase...... usus Basa Purin dan Pirimidin Guanosin Adenosin Xantine Asam Urat Hipoxantin Inosin Ilustrasi 3. Metabolisme Nukleotida Purin Purin berasal dari metabolisme makanan dan asam nukleat endogen, serta didegradasi menjadi asam urat melalui enzim xantine oksidase. Purin diubah menjadi adenosin selanjutnya diubah menjadi adenin dan isonine oleh enzim adenin deaminase dan phosphorilase. Keduanya diubah menjadi hipoxantine. Xantine oksidase mengubah hipoxantine menjadi xantine dan akhirnya diubah 14 menjadi asam urat. Adenosin, selain berasal dari metabolisme purin, dapat berasal dari jaringan yang mengalami hipoksia (Emmerson, 1996; Waring dkk., 2000; Johnson dkk., 2003). 2.5 Urea Darah Urea adalah suatu senyawa organik, berperan penting dalam metabolisme senyawa yang mengandung nitrogen (Kurzer dan Sanderson, 1956) dan produk akhir dari metabolisme nitrogen yang disintesa dari amonia, karbon dioksida dan nitrogen amida aspartat (Murray dkk., 1999). Sintesis urea terjadi di hati melalui siklus urea atau juga dikenal sebagai siklus ornithine dan siklus Krebs-Henseleit (Krebs, 1964). Langkah awal dalam produksi urea adalah sintesis karbamoilfosfat dari bikarbonat dan amonia. Bikarbonat diperoleh dari hidrasi karbon dioksida, dikatalis oleh karbonik anhidrase (Dodgson dan Forster, 1986). Amonia diperoleh dari darah portal atau dari produksi intramitochondrial amonia dari glutamin, dikatalis oleh glutaminase (Haussinger dkk., 1990). Pembentukan karbamoilfosfat memerlukan jumlah amonia yang signifikan, maka peran glutaminase dalam mitokondria adalah untuk meningkatkan konsentrasi amonia. Glutaminase diaktifkan oleh produk akhirnya yaitu amonia, menciptakan "nitrogen feed-forward activation" dan memungkinkan meningkatkan konsentrasi amonia ke tingkat yang diperlukan untuk sintesis urea (Cooper, 1990). 15 Ilustrasi 4. Pembentukan Urea dari CO2 dan Amonia di dalam Sel Hati Pembentukan karbamoilfosfat terjadi di mitokondria, dikatalisi oleh karbamoil fosfat sintetase I (CPS I). Reaksi ini membutuhkan magnesium adenin trifosfat (MgATP) dan melepaskan Mg2+, dengan N-acetylglutamate sebagai alosterik activator (Hall dkk., 1958; Meijer dkk., 1990). CPS I adalah enzim pembatas dari siklus urea. Arginine mengaktifkan produksi N-acetylglutamate dan dengan demikian memberikan kontrol tambahan atas produksi urea (Meijer dkk., 1990). Langkah selanjutnya adalah pembentukan citrulline dari karbamoil fosfat dan ornithine, dikatalisasi oleh transkarbamoilase ornithine, enzim mitokondria yang lain, diikuti oleh komponen sitosol dari siklus urea, dimulai dengan enzim arginosuccinate sintetase. Enzim ini mengkatalisis reaksi yang menggabungkan citrulline dengan aspartat, pendonor nitrogen kedua untuk membentuk 16 arginosuccinate. Aspartat diperoleh dari oksaloasetat (dari siklus asam sitrat) dan nitrogen disumbangkan dari glutamate. Arginosuccinate dipecah menjadi arginin dan fumarat melalui aksi arginosuccinate liase, selanjutnya fumarat dapat masuk ke siklus asam sitrat (Visek, 1979). Arginin dipecah oleh arginase menjadi urea dan ornitin. Urea dilepaskan ke dalam sirkulasi, dan ornithine masuk kembali ke dalam siklus urea (Meijer dkk., 1990).