2. tinjauan pustaka
advertisement

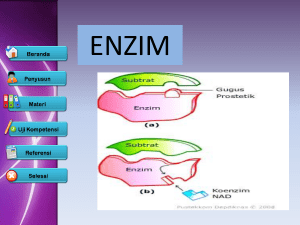
4 2. TINJAUAN PUSTAKA 2.1 Enzim Transglutaminase Transglutaminase termasuk ke dalam kelompok enzim transferase dan mempunyai nama sistematis, yaitu protein glutamin γ-glutamyltransferase (EC 2.3.2.13). Transglutaminase merupakan enzim yang mengkatalisis reaksi perpindahan gugus asil antara kelompok γ-carboxyamide residu glutamin pada protein, peptida dan berbagai amina primer. Apabila kelompok ε-amino residu lisin aktif sebagai aseptor asil, maka akan menghasilkan proses polimerisasi dan interaksi silang inter- atau intra-molekul protein melalui pembentukan ikatan ε-(γ-glutamyl) lisin. Pada proses ini terjadi pertukaran amonia dari kelompok ε-amino residu lisin ke kelompok carboxyamide residu glutamin pada molekul protein. Apabila amin primer tidak ada, maka air dapat berperan sebagai aseptor asil dan menghasilkan proses deamidasi kelompok γ-carboxyamide glutamin untuk membentuk asam glutamat (Haard dan Simpson, 2000). Reaksi katalisasi enzim transglutaminase dapat dilihat pada Gambar 1. Gambar 1. Transglutaminase mengkatalisis reaksi: a) reaksi perpindahan asil b) interaksi silang residu lisin dan glutamin dari protein c) deamidasi (Folk, 1980). Istilah “transglutaminase” pertama kali diperkenalkan oleh Clarke et al., (1957) yang menjelaskan aktivitas transamidasi pada liver marmut. Sekarang, transglutaminase sudah ditemukan pada tumbuhan, invertebrata, amfibi, 5 burung, ikan dan mikroorganisme. Enzim dari berbagai sumber tersebut mengkatalis perubahan post-translational protein dengan membentuk ikatan isopeptida melalui interaksi silang protein atau penggabungan amina (Folk, 1980). Transglutaminase tersebar secara luas pada organisme eukariot dan prokariot. Akan tetapi karakteristik transglutaminase yang paling baik, yaitu berasal dari mamalia. Berdasarkan sumbernya, enzim transglutaminase dibagi ke dalam dua golongan, yaitu: a). Mammalian Transglutaminase Aktivitas transglutaminase ditemukan dan tersebar secara luas dalam plasma, jaringan dan cairan ekstraseluler dari beberapa mamalia. Transglutaminase yang merubah protein seluruhnya ada dalam tubuh. Enzim transglutaminase tersebut dapat digolongkan ke dalam lima golongan, yaitu faktor XIII (plasma dan plasenta), transglutaminase jaringan, keratinosit, epidermis dan prostata (Greenberg et al., 1991; Aeschilman dan Paulsson, 1994 dalam Hemung, 2007). Sifat biokimia dari lima jenis mammalian transglutaminase dapat dilihat pada Tabel 1. Perbedaan dari kelima jenis mammalian transglutaminase terletak pada berat molekul, struktur subunit dan aktivitas protease, tetapi semuanya memiliki ketergantungan yang sama pada kalsium. Tabel 1. Sifat biokimia dari lima jenis mammalian transglutaminase TGase Berat molekul (KDa) Struktur subunit Aktivitas protease Ketergantungan pada kalsium Faktor XIII - Plasma 360 a2b2 ada ada - Plasenta 166 a2 ada ada Jaringan 85 monomer tidak ada Keratinosit 90 monomer tidak ada Epidermis 80 monomer ada ada Prostata 150 homodimerik tidak ada Sumber : Greenberg et al., (1991); Aeschilman dan Paulsson (1994) dalam Hemung (2007) b). Varietas Transglutaminase lainnya Enzim yang mengkatalisis pembentukan ikatan ε-(γ-glutamyl) lisin ditemukan pada berbagai organisme. Contohnya, aktivitas transglutaminase yang ditemukan pada ikan (Araki dan Seki, 1993), lobster (Myhrman dan BrunerLorand, 1970), kepiting sepatu kuda (Tachypleus tridentatus) 6 (Tokunaga et al., 1993) serta yeast Candida albicans (Ruizherrer et al., 1995). Selain itu, transglutaminase ditemukan pada berbagai bakteri seperti Bacillus subtilis (Suzuki et al., 2000), Streptoverticillium sp. (Ando et al., 1989), dan Streptomyces sp. (Umezawa et al., 2002) serta tumbuhan Medicago sativa L. (Margosiak et al., 1990). Struktur kristal dari microbial transglutaminase dapat dilihat pada Gambar 2. Aktivitas transglutaminase yang dihasilkan oleh seluruh organisme tersebut tergantung pada Ca2+, kecuali bakteri dan tumbuhan. Gambar 2. Struktur kristal dari microbial transglutaminase (Kashiwagi et al., 2002) 2.2 Karakterisasi Biokimia Enzim Aktivitas enzim dipengaruhi oleh beberapa faktor tertentu yang menentukan efektivitas kerja suatu enzim. Apabila faktor pendukung tersebut berada pada kondisi yang optimum, maka kerja enzim juga akan optimal. Hasil karakterisasi enzim walaupun masih bersifat ekstrak kasar, namun dapat menggambarkan karakter enzim di dalamnya. Kondisi lingkungan harus menunjang kondisi yang dibutuhkan enzim untuk dapat berfungsi sebagai katalis reaksi (Ryta, 2001). 2.2.1 Suhu Reaksi enzim umumnya terdiri dari beberapa tahapan reaksi dan respon terhadap suhu dari masing-masing tahap berbeda. Setiap enzim memiliki aktivitas pada suhu tertentu. Aktivitas akan meningkat dengan meningkatnya suhu, akan tetapi setelah suhu optimum tercapai maka yang terjadi adalah sebaliknya, yaitu akan menurun dengan peningkatan suhu. Hal terdenaturasinya protein enzim (Pelczar dan Chan, 1986). ini disebabkan telah 7 Enzim adalah suatu protein, maka kenaikan suhu dapat menyebabkan terjadinya denaturasi. Apabila terjadi proses denaturasi, maka bagian aktif enzim akan terganggu dan dengan demikian konsentrasi efektif enzim makin menjadi berkurang dan kecepatan reaksinya juga akan menurun. Kenaikan suhu sebelum terjadinya proses denaturasi dapat menaikan kecepatan reaksi, akan tetapi kenaikan suhu pada saat mulai terjadinya proses denaturasi akan mengurangi kecepatan reaksi (Poedjiadi, 1994). Peningkatan suhu tertentu menyebabkan semakin meningkatnya aktivitas katalitik dan semakin bertambahnya proses kerusakan enzim (Palmer, 1991). Peningkatan suhu menyebabkan putusnya ikatan hidrogen dan hidrofobik lemah yang mempertahankan struktur sekunder-tersier dari enzim, sehingga enzim mengalami denaturasi (Suhartono, 1989). Denaturasi adalah rusaknya bentuk tiga dimensi enzim yang menyebabkan enzim tidak dapat lagi berikatan dengan substrat. Denaturasi menyebabkan aktivitas enzim menurun. Dan jika suhu di bawah suhu optimum maka enzim tidak dapat bekerja dengan baik atau energi aktivasinya juga akan menurun. Struktur protein menentukan aktivitas enzim, jika srukturnya terganggu maka aktivitasnya akan berubah pula. Kenaikan suhu sampai batas tertentu dalam suatu reaksi menyebabkan peningkatan kecepatan reaksi enzim karena bertambahnya energi kinetik yang mempercepat gerak vibrasi, translasi dan rotasi enzim dan substrat sehingga memperbesar peluang keduanya untuk bereaksi. Pada suhu yang lebih besar dari batas reaksi, protein enzim dapat mengalami perubahan konformasi yang bersifat detrimal, yaitu berubahnya susunan tiga dimensi yang khas dari rantai polipeptida. Hal yang sama juga dapat terjadi pada substrat yang perubahan konformasinya dapat menyebabkan gugus reaktifnya akan mengalami kesulitan pada saat memasukan sisi aktif enzim (Suhartono, 1989). Suhu optimum untuk aktivitas transglutaminase, yaitu antara 45-55oC, tergantung pada spesies Streptoverticillium. Transglutaminase dari o Stv. mobaraense memiliki aktivitas optimum pada suhu 55 C sedangkan suhu optimum untuk transglutaminase dari Stv. griseocarneum dan Stv. cinnamoneum spp. adalah 45oC. Stabilitas enzim akan menurun dengan meningkatnya suhu. Aktivitas transglutaminase dari Stv. mobaraense bertahan 8 lama pada suhu 40oC sedangkan pada suhu diatas 50oC aktivitasnya menurun (Ando et al., 1989). Suhu optimum untuk transglutaminase dari mamalia lebih tinggi dari suhu tubuhnya, yaitu sekitar 55oC. Apabila suhunya lebih dari 55 oC, aktivitas enzim akan turun dengan cepat, karena enzim mengalami perubahan konformasi atau terdenaturasi (Jiang dan Lee, 1992 dalam Negus, 2001). 2.2.2 pH Seperti protein pada umumnya, struktur ion enzim tergantung pada pH lingkungannya. Enzim dapat bermuatan ion positif, ion negatif, atau ion bermuatan ganda (zwitterion). Dengan demikian perubahan pH lingkungan akan berpengaruh terhadap aktivitas bagian aktif enzim dalam membentuk kompleks enzim-substrat. Disamping pengaruh terhadap struktur ion pada enzim, pH rendah, atau pH tinggi dapat pula menyebabkan terjadinya proses denaturasi dan ini akan mengakibatkan menurunnya aktivitas enzim (Poedjiadi, 1994). Enzim memiliki pH optimum yang khas, yaitu pH yang menyebabkan aktivitas maksimal. Profil aktivitas pH enzim menggambarkan pH pada saat gugus pemberi atau penerima proton yang penting pada sisi katalitik enzim berada dalam tingkat ionisasi yang diinginkan. Nilai pH optimum tidak perlu sama dengan pH lingkungan normalnya, dengan pH yang mungkin sedikit berada di atas atau di bawah pH optimum. Aktivitas katalitik enzim di dalam sel mungkin diatur sebagian oleh perubahan pada pH medium lingkungan (Lehninger, 1993). Pada umumnya, enzim bersifat ampolitik, yang berarti enzim mempunyai konstanta disosiasi pada gugus asam maupun gugus basanya terutama pada gugus residu terminal karboksil dan gugus terminal aminonya. Diperkirakan perubahan keaktifan enzim ini adalah akibat perubahan pH lingkungan. Perubahan pH lingkungan ini terjadi karena adanya perubahan ionisasi pada gugus ionik enzim pada sisi aktifnya atau sisi lain yang secara tidak langsung mempengaruhi sisi aktif. Gugus ionik berperan dalam menjaga konformasi sisi aktif dalam meningkatkan substrat menjadi produk. Perubahan ionisasi juga dapat dialami oleh substrat atau kompleks enzim-substrat, yang juga berperan terhadap aktivitas enzim (Webb dan Dixon, 1979). 9 2.2.3 Ion logam Banyak enzim yang memerlukan tambahan komponen kimia bagi aktivitasnya. Komponen ini disebut dengan kofaktor. Kofaktor bisa berupa molekul anorganik, seperti ion Fe2+, Mn2+, Zn2+, atau mungkin juga suatu molekul organik kompleks yang disebut koenzim, seperti thiamin pirofosfat, FAD, serta koenzim A. Beberapa enzim membutuhkan baik koenzim maupun satu atau lebih ion logam bagi aktivitasnya. Pada beberapa enzim, koenzim atau ion logam lainnya hanya terikat secara lemah atau dalam waktu sementara. Akan tetapi pada enzim lain senyawa ini terikat secara kuat dan permanen. Dalam hal ini disebut gugus protetik. Enzim yang strukturnya sempurna dan aktif mengkatalisis bersama-sama dengan koenzim atau gugus logamnya disebut haloenzim. Koenzim dan ion logam bersifat stabil selama pemanasan, sedangkan bagian protein enzim disebut apoenzim akan terdenaturasi oleh pemanasan (Lehninger, 1993). Ion logam mempunyai peranan penting dalam menjaga kestabilan enzim. Logam biasanya berperan sebagai pengatur aktivitas enzim (Harper et al., 1979). Ion logam dapat mengaktifkan enzim melalui berbagai kemungkinan seperti: (a) menjaga bagain internal enzim, (b) menghubungkan enzim dengan substrat, (c) merubah konstanta keseimbangan reaksi enzim, (d) merubah tegangan permukaan protein enzim, (e) menghilangkan inhibitor, (f) menggantikan ion logam yang tidak efektif pada sisi aktif enzim maupun substrat, dan (g) merubah konformasi enzim menjadi konformasi yang lebih aktif (Richardson dan Hylop, 1985). Lebih dari 25% jenis enzim mengandung ion logam yang terikat atau memerlukan ion logam bagi aktivitasnya. Metaloenzim mengandung ion logam fungsional dalam jumlah pasti, yang dipertahankan selama proses pemurnian. Enzim yang diaktifkan oleh logam memperlihatkan ikatan yang lebih lemah dengan logam, dengan demikian memerlukan logam tambahan. Oleh karena itu, perbedaan metaloenzim dengan enzim yang diaktifkan oleh logam terletak pada afinitas suatu enzim tertentu terhadap ion logamnya (Harper et al., 1979). 2.2.4 Senyawa inhibitor Telah diketahui bahwa mekanisme enzim dalam suatu reaksi ialah melalui pembentukan kompleks enzim-substrat (ES). Oleh karena itu, hambatan atau inhibisi pada suatu reaksi yang menggunakan enzim sebagai katalis dapat terjadi 10 apabila penggabungan substrat pada bagian aktif enzim mengalami hambatan. Molekul atau ion yang dapat menghambat reaksi tersebut dinamakan inhibitor (Poedjiadi, 1994). Hambatan yang dilakukan oleh inhibitor dapat berupa hambatan irreversible dan reversible. Hambatan irreversible pada umumnya disebabkan oleh terjadinya proses destruksi atau modifikasi sebuah gugus fungsi atau yang terdapat pada molekul enzim. Hambatan reversible dapat berupa hambatan bersaing atau hambatan tidak bersaing (Poedjiadi, 1994). Inhibitor merupakan senyawa yang cenderung menurunkan kecepatan reaksi yang dikatalisis oleh enzim. Inhibitor dapat bereaksi dengan substrat, kofaktor atau dengan enzim langsung. Microbial transglutaminase tidak dipengaruhi ion logam Ca2+, sehingga adanya senyawa pengkelat logam, seperti ethylenediamine tetraacetic acid (EDTA) tidak menghambat aktivitasnya (Lin et al., 2008). Sebaliknya, transglutaminase yang dipengaruhi ion logam Ca 2+ dapat dihambat oleh EDTA, misalnya tilapia transglutaminase (Worratao dan Yongsawatdigul 2005). Aktivitas microbial transglutaminase dapat dihambat kuat oleh (p-Chloromercuribenzoate) PCMB serta sedikit dihambat oleh (N-ethylmaleimide) NEM dan (Phenyl methyl sulfonyl fluoride) PMSF. 2.3 Streptoverticillium ladakanum Klasifikasi bakteri Streptovercillium ladakanum adalah sebagai berikut (Wikispesies, 2007) : Filum : Actinobacteria Kelas : Actinobacteria Ordo : Actinomycetales Famili : Streptomycetaceae Genus : Streptoverticillium Spesies : Streptoverticillium ladakanum Streptoverticillium memiliki ukuran besar dengan diameter antara 0,8-1,2 µm. Ciri pemersatu ialah pleomorfisme sel-selnya dan kecenderungan membentuk filamen (hifa) bercabang. Pada beberapa spesies, hifa-hifa itu bersatu membentuk miselium (Pelczar dan Chan, 1986). Setiap cabang vertisil memiliki puncak 11 cabang (umbel) yang terdiri dari dua atau beberapa rantai spora yang berbentuk bola dan spiral (Holt et al., 1994). Beberapa spesies menghasilkan pigmen yang dapat larut, substrat berwarna dan aerial mycelium. Bakteri ini resisten pada lysozyme dan neomycin serta mampu menghasilkan komponen yang menunjukkan aktivitas seperti antifungal, antibakteri, antiprotozoa dan antitumor serta sensitif terhadap agen antibakteri dan actinophage. Streptoverticillium termasuk ke dalam kelompok bakteri Gram positif. Dinding selnya mengandung L-diaminopimelic acid (L-DAP). Kandungan utama sel bakteri ini adalah saturated, iso- dan anteiso-fatty acid, MK-9 (H6) dan MK-9 (H8) menaquinone phosphatidylethanolamine, serta phospholipid phosphatidylinositol (diphosphatidylglycerol, dan phosphatidylinositol mannoside) (Holt et al., 1994). Gambar 3. Bakteri Streptoverticillium sp. (Tresner et al., 1971) Streptoverticillium bersifat aerobik, yaitu tumbuh pada kondisi lingkungan yang banyak oksigen atau konsentrasi karbondioksida sedikit. Bakteri ini juga bersifat kemoorganotrof dan mesopilik. Tumbuh optimum pada temperatur 26-32oC dan pH 6,5-8,0. Bakteri ini kebanyakan hidup saprofit pada tanah (Holt et al., 1994). Reproduksi bakteri yang termasuk Streptoverticillium terjadi dari salah satu miselium aerial atau dari germinasi spora. Spora tersebut memiliki permukaan yang halus sampai sedikit kasar (Holt et al., 1994). 2.4 Medium Pertumbuhan Mikroba Medium pertumbuhan adalah tempat untuk menumbuhkan mikroba. Mikroba memerlukan nutrisi untuk memenuhi kebutuhan energi dan untuk bahan 12 pembangun sel, untuk sintesa protoplasma dan bagian-bagian sel lain. Setiap mikroba mempunyai sifat fisiologi tertentu, sehingga memerlukan nutrisi tertentu pula. Media pertumbuhan dapat berupa media sintetik maupun media alami. Pada media sintetik, setiap komponen merupakan senyawa yang relatif murni serta konsentrasi komponen dalam media dan strukturnya dapat diketahui dengan pasti. Media sintetik mempunyai keuntungan, antara lain: setiap komponen dapat dengan mudah diubah, disamping kesalahan atau kelainan yang mungkin terjadi selama fermentasi akibat komposisi media dapat dicegah (Yuniasari, 1994). Faktor yang penting dalam optimasi media fermentasi adalah pemilihan komposisi sumber karbon dan nitrogen karena sel-sel mikroorganisme dan berbagai produk fermentasi sebagian besar terdiri dari unsur karbon dan nitrogen serta garam-garam dalam jumlah seimbang (Suhartono, 1989). Gula sederhana seperti glukosa merupakan sumber karbon yang mudah dicerna dan digunakan mikroorganisme sebagai sumber energi. Penggunaan glukosa dalam media harus dibatasi, karena selain pertimbangan ekonomis juga untuk menghindari efek represi produksi enzim (Suhartono, 1989). Bakteri akan melakukan hidrolisa secara perlahan pada media yang mengandung senyawa karbon dan sumber nitrogen untuk mencegah terjadinya represi katabolit (proses yang menyebabkan fase lag menjadi berkepanjangan sehingga mempengaruhi sifat perumbuhan dan pembentukan produk yang menyebabkan penurunan aktivitas enzim) dan produksi enzim meningkat (Bierbaum et al., 1994). Sumber nitrogen yang digunakan dalam media fermentasi dapat berupa nitrogen organik maupun nitrogen anorganik. Contoh sumber nitrogen organik untuk pertumbuhan mikroba, antara lain pepton, ekstrak khamir, tepung kedelai dan limbah organik (Suhartono, 1989). Ekstrak khamir dan pepton merupakan sumber asam amino atau nitrogen dan vitamin B sebagai stimulan pertumbuhan bakteri. Mikroba membutuhkan vitamin B untuk pertumbuhannya, seperti vitamin B1 (thiamin), B2 (riboflavin), B6 (piridoksin), dan B12 (kobalamin). Vitamin B ini digunakan dalam pembentukan koenzim yang akan berikatan dengan enzim dengan ikatan yang tidak begitu kuat (Moat, 1979). 13 Kebutuhan nutrisi mikroba terlihat dalam unsur-unsur yang tersusun dalam selnya. Susunan kimia sel mikroba relatif tetap, baik unsur kimia maupun senyawa yang terkandung di dalam sel. Komponen utama yang menyusun sel bakteri adalah unsur C, H, O, N, P dan S, yang jumlahnya 95% dari berat kering sel, sedangkan sisanya tersusun dari unsur-unsur lain (Irianto, 2006). Mikroba membutuhkan nitrogen sebagian besar untuk sintesis protein dan asam-asam nukleat. Nitrogen digunakan dalam bentuk amonium, nitrat, asam amino, protein, dan sebagainya. Sumber karbon untuk mikroba dapat berbentuk senyawa organik maupun anorganik. Hidrogen dan oksigen biasanya digunakan sebagai air untuk keperluan seperti pelarut, hidrolisis, ionisasi, osmosis, dan sebagianya (Irianto, 2006). Seluruh mikroorganisme membutuhkan sumber sulfur, nitrogen dan fosfat sebagai sumber mineral trace. Komponen mineral utama yang umumnya dibutuhkan semua jenis mikroorganisme adalah fosfat, kalium, kalsium, sulfur dan magnesium. Penambahan kasium seringkali meningkatkan aktivitas enzim yang dihasilkan. Beberapa jenis mineral biasanya sudah terdapat bersama-sama dengan komposisi substrat, seperti besi, tembaga, mangan, seng dan sebagainya. Fosfat, sulfat dan anion lain diberikan sebagai garam mineral (Suhartono, 1989). 2.5 Limbah Cair Surimi Limbah cair didefinisikan sebagai buangan cair yang berasal dari suatu lingkungan masyarakat dan lingkungan industri dimana komponen utamanya adalah air yang telah digunakan dan mengandung benda-benda padat yang terdiri dari zat-zat organik dan anorganik (Mahida, 1984). Air pencucian surimi merupakan hasil samping atau bahkan menjadi limbah bagi industri pengolahan surimi, sehingga berpotensi menimbulkan pencemaran perairan jika tidak ditangani dengan baik karena mengandung bahan organik yang tinggi (Uju et al., 2009). Kadar BOD (Biochemical Oxygen Demand) dari proses pencucian minced fish berkisar antara 6000-27000 mg/l (Lin et al., 1995). Air dari proses pencucian surimi tidak hanya membawa protein sarkoplasma yang larut air, akan tetapi protein miofibril, protease, hemepigmen dan zat bioaktif lainnya yang sangat potensial (Tacharatanamanee et al., 2004). Air limbah yang dihasilkan dari pencucian pertama pada proses pembuatan surimi mengandung 14 protein, non protein, nitrogen, lemak dan abu yang tinggi (Lin et al., 1995). Air cucian minced fish mengandung protein 1,58% (b/v) dan mengandung 17 asam amino dengan asam glutamat sebagai komponen dominan (Trilaksani et al., 2007). Secara spesifik, Bourtoom et al., (2009) melaporkan kadar protein air cucian minced fish tahap pencucian pertama sebesar 1,23 mg/ml, sedangkan pada pencucian kedua dan ketiga lebih rendah, yaitu 0,64 dan 0,54 mg/ml. Kadar protein ini 1,8 kali lebih tinggi dibandingkan kadar protein air pasteurisasi rajungan (Uju et al., 2008). Asam amino dari konsentrat air limbah surimi ikan nila (Oreochromis niloticus) dapat dilihat pada Tabel 2. Tabel 2. Asam amino konsentrat (Oreochromis niloticus) No. Jenis asam amino 1 Asam aspartat 2 Asam glutamat 3 Serin 4 Glisin 5 Histidin 6 Argini 7 Treonin 8 Alanin 9 Prolin 10 Tirosin 11 Valin 12 Metionin 13 Sistin 14 Isoleusin 15 Leusin 16 Fenilalanin 17 Lisin Sumber : Trilaksani et al., (2007) air limbah surimi ikan nila Nilai rata-rata (%) 3,79 + 0,04 5,49+ 0.05 1,13 + 0,05 1,64 + 0,04 0,49 + 0,03 0,94 + 0,01 2,45 + 0,06 2,03 + 0,03 1,12 + 0,03 1,35 + 0,02 0,32 + 0,06 0,98 + 0,09 0,76 + 0,01 3,23 + 0,05 2,73 + 0,01 1,64 + 0,05 1,35 + 0,04 2.6 Aplikasi Transglutaminase dalam Industri Modifikasi protein dengan menggunakan transglutaminase sudah dimanfaatkan pada berbagai industri makanan. Transglutaminase berfungsi dalam meningkatkan sifat rheologi, enkapsulasi bahan yang berlemak dan larut lemak, memperbaiki pembentukan gel dan sifat gel, perubahan daya larut protein serta sifat busa dan daya ikat air (Motoki dan Seguro, 1998; Jaros et al., 2006 dalam Lantto, 2007). Kebutuhan industri makanan terhadap enzim transglutaminase sudah dapat dipenuhi semenjak beberapa instansi internasional menggunakan 15 microbial transglutaminase yang dihasilkan dari mikroorganisme Streptoverticillium mobaraense pada industri makanan (Grades, 2006). Microbial transglutaminase (MTGase) komersial yang diproduksi oleh Ajinomoto sudah diaplikasikan pada berbagai makanan berprotein untuk meningkatkan karakteristik beberapa produk makanan komersial, seperti kenampakan, tekstur, viskositas dan gelasi. Pembentukan interaksi silang tambahan menyebabkan perubahan pada ukuran, konformasi, viskositas, gelasi dan kestabilan beberapa makanan berprotein seperti kedelai, ketan, urat daging , miosin, globulin dan kasein sehingga penampakan dan tekstur berbagai makanan dapat dimodifikasi dengan menggunakan MTGase selama proses pembuatan makanan. Menurut Hazová et al., (2002) dalam Grades (2006), penambahan MTGase ke dalam tepung menimbulkan pengaruh positif pada tekstur roti dan kue kering dengan konsentrasi enzim sebesar 3,5-4,5 mg/kg tepung. Hasil penelitian Lauber et al., (2002) dalam Grades, (2006) menunjukan bahwa penamabahan MTGase pada yoghurt dari susu skim yang diberi perlakukan dengan suhu 80 oC selama 5 menit memberikan pengaruh yang nyata. Hasil penelitian tersebut disajikan pada Gambar 4. a) b ) Gambar 4. Yoghurt dari susu skim yang diberi perlakukan dengan suhu 80oC selama 5 menit: a) tanpa MTGase b) dengan 100 U MTGase/L (Lauber et al., 2002 dalam Grades, 2006). Transglutaminase juga dapat dimanfaatkan pada industri tekstil. Misalnya, transglutaminase digunakan untuk memperbaiki kerusakan yang disebabkan oleh bahan kimia dan protease selama pembuatan wool (Cortez et al., 2004) dan pencucian benang wool (Cortez et al., 2005). Beberapa aplikasi microbial transglutaminase pada pengolahan makanan dapat dilihat pada Tabel 3. 16 Tabel 3. Aplikasi microbial transglutaminase pada pengolahan makanan Sumber Daging Produk Hamburger, bakso, stuffeddumplings, shao-mai Daging kaleng Daging beku Ikan Daging cetak Fish paste Kolagen Terigu Imitasi sirip ikan hiu Makanan kue Kacang kedelai Mapuo doufu Tofu goreng (aburaage) Tofu Sayuran dan buahbuhaan Kasein Seledri Promotor absorpsi mineral Gelatin Ikatan silang protein Makanan manis Seasoning Seasoning Sumber : Zhu et al., (1995) Pengaruh Meningkatkan elastisitas, tekstur, rasa dan flavor Tekstur dan penampakan menjadi baik Meningkatkan tekstur dan mengurangi biaya Merestrukturisasi daging Meningkatkan tekstur dan penampakan Imitasi pelezat makanan Meningkatkan tekstur dan volume Memperpanjang daya simpan Meningkatkan tekstur Memperpanjang daya simpan Pengawet makanan Meningkatkan absorpsi mineral pada usus Mengurangi allergenicity Makanan rendah kalori dengan tekstur baik, kokoh dan elastis Meningkatkan rasa dan flavour Reference Sakamoto and Soeda (1991) Seguro and Motoki (1991) Takagaki dan Narukawa (1990) Matsui et al., (1990) Wakameda et al., (1990b) Tani et al., (1990 Ashikawa et al., (1990) Kato et al., (1991) Soeda et al., (1990) Nonaka et al., (1990) Takagaki et al., (1991) Noguchi et al., (1992) Yamauchi et al., (1991) Yamanaka and Sakai (1992) Kobata et al., (1990)