makalah biokimia
advertisement

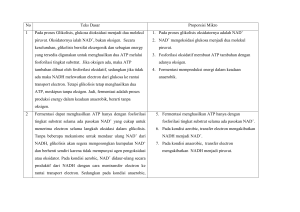
MAKALAH BIOKIMIA II “DEKARBOKSILASI OKSIDATIF, SIKLUS ASAM SITRAT, DAN FOSFORILASI OKSIDATIF” OLEH KELOMPOK IV NAMA ANGGOTA : 1. LALU SINGGIH AJI PUTRA 2. NONI MULIANA LISTIAWATI 3. SAMSUL RIZAL UMAMI 4. SITI FATIMAH 5. QOMARIATUL KIBITIA FAKULTAS KEGURUAN DAN ILMU PENDIDIKAN UNIVERSITAS MATARAM 2016 RESPIRASI SEL Pada metabolisme karbohidrat salah satu substansi yang penting yang akan dipelajari adalah oksidasi glukosa yang disebut dengan respirasi sel. Proses oksidasi ini terjadi secara aerobik. Dimana glukosa ini akan di oksidasi lebih lanjut menjadi energi. Karena glukosa termasuk ke dalam senyawa organik yang mengandung karbon, hidrogen dan oksigen maka dapat dioksidasi secara sempurna menjadi CO2 dan H2O. Selama proses ini energi yang berasal dari senyawa organik sebagian besar dikonversi menjadi panas atau kalor. Di sebagian besar sistem biologi, energy dilepaskan dari molekul, tidak berasal dari pembakaran tetapi oleh kontrol metabolisme oksidasi yang disebut dengan proses respirasi. Oksidasi aerobik glukosa ini memerlukan partisipasi dari tiga proses metabolisme yang saling terkait (Siklus asam sitrat, transpor elektron dan fosforilasi oksidatif) semua proses ini terjadi di mitokondria. Mitokondria sering disebut dengan pabrik energi dari sel. Di dalam materi ini akan dibahas salah satu bagian dari sebagian besar proses respirasi sel yaitu oksidasi rantai karbon dari piruvat untuk membentuk ko-enzim dan beberapa ATP. Piruvat yang digunakan dibentuk oleh proses glikolisis, piruvat yang dihasilkan dari proses glikolisis ini tidak dikonversi menjadi laktat, tetapi piruvat ini dioksidasi untuk membentuk CO2 dan Asetil-KoA yang melalui proses enzimatik. Langkah pertama dalam oksidasi piruvat ini adalah reaksi dekarboksilasi oksidatif yang melibatkan ko-enzim A. Produk dari reaksi ini adalah CO2 dan asetil ko-A. Dua karbon ditambahkan dari ko-enzim A. Selanjutnya Asetil-Koa yang dihasilkan dibawa ke siklus kreb untuk diubah lebih lanjut menjadi ATP (Horton, 2002: 372). DEKARBOKSILASI OKSIDATIF, SIKLUS ASAM SITRAT, TRANSPORT ELEKTRON & FOSFORILASI OKSIDATIF 1. Dekarboksilasi Oksidatif Dekarboksilasi oksidatif adalah reaksi yang mengubah asam piruvat yang beratom 3 C menjadi senyawa baru yang beratom 2 C, yaitu asetil koenzim-A (asetil koA). Reaksi dekarboksilasi oksidatif ini (disingkat DO) sering juga disebut sebagai tahap persiapan untuk masuk ke siklus Krebs. Reaksi DO ini mengambil tempat di intermembran mitokondria. Setelah melalui reaksi glikolisis, jika terdapat molekul oksigen yang cukup maka asam piruvat akan menjalani tahapan reaksi selanjutnya, yaitu siklus Krebs yang bertempat di matriks mitokondria. Jika tidak terdapat molekul oksigen yang cukup maka asam piruvat akan menjalani reaksi fermentasi. Akan tetapi, asam piruvat yang mendapat molekul oksigen yang cukup akan meneruskan tahapan reaksi yaitu ke siklus krebs. Namun, asam piruvat ini tidak dapat begitu saja masuk ke dalam siklus Krebs, karena asam piruvat memiliki atom C terlalu banyak, yaitu 3 buah. Persyaratan molekul yang dapat menjalani siklus Krebs adalah molekul tersebut harus mempunyai dua atom C (2 C). Karena itu, asam piruvat akan menjalani reaksi dekarboksilasi oksidatif. Gambar tahap dekarboksilasi oksidatif. Reaksi oksidasi piruvat hasil glikolisis menjadi asetil koenzim-A, merupakan tahap reaksi penghubung yang penting antara glikolisis dengan jalur metabolisme lingkar asam trikarboksilat (daur Krebs). Reaksi yang dikatalisis oleh kompleks piruvat dehidrogenase dalam matriks mitokondria melibatkan tiga macam enzim yaitu piruvat dehidrogenase, dihidrolipoamid asetil transasetilase, dan dihidrolipoamid dehidrogenase dan lima macam gugs prostetik yaitu Tiamin pirofosfat (TPP), asam lipoat, koenzim-A, Flavin Adenin Dinukleotida (FAD), dan Nikotinamid Adenin Dinukleotida (NAD) yang berlangsung dalam lima tahap reaksi. Pertama-tama, molekul asam piruvat yang dihasilkan reaksi glikolisis akan melepaskan satu gugus karboksilnya yang sudah teroksidasi sempurna dan mengandung sedikit energi, yaitu dalam bentuk molekul CO2. Setelah itu, 2 atom karbon yang tersisa dari piruvat akan dioksidasi menjadi asetat (bentuk ionisasi asam asetat). Selanjutnya, asetat akan mendapat transfer elektron dari NAD+ yang tereduksi menjadi NADH. Kemudian, koenzim A (suatu senyawa yang mengandung sulfur yang berasal dari vitamin B) diikat oleh asetat dengan ikatan yang tidak stabil dan membentuk gugus asetil yang sangat reaktif, yaitu asetil koenzim-A, yang siap memberikan asetatnya ke dalam siklus Krebs untuk proses oksidasi lebih lanjut. Selama reaksi transisi ini, satu molekul glukosa yang telah menjadi 2 molekul asam piruvat lewat reaksi glikolisis menghasilkan 2 molekul NADH. Dekarboksilasi oksidatif akan mengubah asam piruvat menjadi asetil ko-A. Tahap ini terjadi dalam beberapa reaksi yang dikatalisis oleh kompleks enzim yang disebut piruvat dehidrogenase. Enzim ini terdapat pada mitokondria pada sel eukariotik, sedangkan pada prokariotik terdapat pada sitoplasma. Hasil dari dekarboksilasi oksidatif adalah molekul asetil ko-A, NADH, dan CO2. Satu molekul glukosa akan diubah menjadi dua molekul asam piruvat dalam glikolisis, artinya proses dekarboksilasi oksidatif untuk satu molekul glukosa akan menghasilkan 2 molekul asetil ko-A, 2 NADH, dan 2 CO2. Keseluruhan reaksi dekarboksilasi ini bersifat irreversible. 2. Siklus Asam Sitrat Jalur metabolisme daur asam trikarboksilat ( asam sitrat ) pertama kali ditemukan oleh Hans Krebs pada tahun 1937. Oleh karena itu jalur ini disebut pula daur krebs. Siklus asam sitrat adalah serangkaian reaksi kimia dalam sel, yaitu pada mitokondria, yang berlangsung secara berurutan dan berulang, bertujuan mengubah asam piruvat menjadi CO2, H2O dan sejumlah energi. Proses ini adalah proses oksidasi dengan menggunakan oksigen atau aerob. Jalur daur ini merupakan jalur metabolisme yang utama dari berbagai senyawa hasil metabolisme, yaitu hasil katabolisme karbohidrat, lemak, dan protein. Asetil koenzim–A ( sebagai hasil katabolisme lemak dan karbohidrat ), oksalasetat, fumarat, dan α – ketoglutarat ( sebagai hasil katabolisme asam amino dan protein), masuk ke dalam daur krebs untuk selanjutnya dioksidasi melalui beberapa tahap reaksi yang kompleks menjadi CO2, H2O dan energi ATP. Didalam sel eukariot, metabolisme asam trikarboksilat berlangsung didalam mitokondrion. Sebagai enzim dalam metabolisme ini terdapat didalam cairan matrik dan sebagian lagi terikat pada bagian dalam membran mitokondrion. Telah pula diketahui bahwa beberapa enzim dalam siklus asam sitrat ( seperti akonitat hidratase, isositrat dehidrogenase, fumarase, dan malat dehidrogenase ) terdapat di dalam sitoplasma sel jaringan tertentu meskipun di situ tidak ditunjukan adanya kegiatan siklus asam sitrat. Fungsi utama siklus krebs adalah : 1. Menghasilkan karbon dioksida terbanyak pada jaringan manusia. 2. Menghasilkan sejumlah koenzim tereduksi yang menggerakkan rantai pernapasan untuk produksi ATP. 3. Mengkonversi sejumlah energi serta zat intermidiet yang berlebihan untuk digunakan pada sintesis asam lemak. 4. Menyediakan sebagian bahan keperluan untuk sintesis protein dan asam nukleat. 5. Melakukan pengendalian langsung atau tidak langsung (alosterik) terhadap sistem enzim lain melalui komponen-komponen siklus. Tabel reaksi enzimatik dari siklus asam sitrat TAHAP REAKSI DAUR KREBS Pada tahap reaksi pertama, enzim sitrat sintase mengkatalisis reaksi kondensasi antara asetil koenzim–A dengan oksaloasetat menghasilkan sitrat. Reaksi ini merupakan suatu reaksi kondensasi aldol antara gugus metil dari asetil koenzim–A dan gugus karbonil dari oksaloasetat dimana terjadi hidrolisis ikatan tioester dan pembentukan senyawa koenzim–A bebas. Reaksi ini adalah suatu hidrolisis eksergonik yang menghasilkan energi dan merupakan reaksi pendorong pertama untuk daur krebs. Laju reaksi ditentukan oleh jumlah asetil koenzim–A yang tersedia, oksaloasetat dan kadar suksinil koenzim–A yang merupakan zat penghambat saing ( inhibitor kompetitif ) dan sitrat sintase. Tahap reaksi kedua merupakan pembentukan asam isositrat dari sitrat melalui cis-akonitat, dikatalisis secara reversible oleh enzim akonitase. Enzim ini mengkatalisis reaksi reversibel penambahan H2O pada ikatan rangkap cis-akonitat dalam dua arah, yang satu ke pembentukan sitrat dan yang lain ke pembentukan isositrat. Asam sitrat adalah molekul prokiral, yaitu suatu molekul yang dapat bereaksi secara senjang meskipum tidak mempunyai atom karbon yang senjang. Hal ini dimungkinkan karena setengah bagian dari molekulnya bersifat bayangan cermin yang tak dapat ditumpangtindahkan terhadap setengah bagian lainnya. Keadaan ini dapat dibedakan oleh enzim akonitase yang stereospesifik sehingga enzim ini hanya akan menyerang salah satu dari setengah bagian tersebut. percobaan dengan radioisotop membuktikan, akonitase hanya menyerang bagian molekul sitrat yang berasal dari oksalat saja. Telah diketahui akonitase terdapat di dalam jaringan hewan dalam dua bentuk isoenzim, satu dalam mitokondrion dan yang lain dalam sitosol. Tahap reaksi ketiga, Oksidasi isositrat menjadi α – ketoglutarat berlangsung melalui pembentukan senyawa antara oksalosuksinat yang berikatan dengan enzim isositrat dehidrogenase dengan NAD berperan sebagai koenzimnya. Telah diketahui adanya dua macam isositrat dehidrogenase, yang satu berikatan dengan NAD ( NAD – isositrat dehidrogenase ) dan yang lainnya dengan NADP ( NADP – isositrat dehidrogenase ). NAD – Isositrat dehidrogenase terdapat hanya di dalam mitokondrion maupun sitoplasma. Enzim yang pertama mengkatalisis proses oksidasi isositrat menjadi oksalosuksinat dan dekarboksilasi oksalosuksinat menjadi α – ketoglutarat. Pengubahan isositrat ke oksaloasetat dapat dihambat oleh difenilkloroarsin, sedangkan dekarboksilasi oksaloasetat dihambat oleh pirofosfat. Tahap reaksi keempat adalah oksidasi α-ketoglutarat menjadi suksinatmelalui pembentukan subsinil ke koenzim-A. pembentukan suksinil koenzim-A dari α- ketoglutarat adalah reaksi yang irreversible dan dikatalisis oleh enzim kompleks αketoglutarat dehidrogenase. Reaksi ini berlangsung yang sama seperti proses oksidasi piruvat menjadi asetilkoenzim-A dan CO2, yaitu dengan melibatkan koenzim tiamin pirofosfat, asam lipoat, koenzim-A, FAD, dan NAD+. Suksinil koenzim-A adalah suatu senyawa tioester berenergi tinggi. Selanjutnya suksinil koenzim-A melepaskan koenzim-A nya dengan dirangkaikan dengan reaksi pembentukan energy, GTP dari GDP dan fosfat. Reaksi ini dikatalisis oleh enzim suksinil koenzim-A sintetase yang khas untuk GDP. Selanjutnya GTP yang terbentuk dari reaksi ini dipakai untuk sintesis ATP dari ADP dengan enzim nukleosida difosfat kinase. Reaksi : GTP + ADP GDP + ATP Pembentukan GTP ( atau ATP) yang dikaitkan dengan reaksi deasilasi suksinil koenzim-A ini disebut fosforilase tingkat substrat. Ini berlainan dengan proses fosforilasi yang dihubungkan dengan rantai pernapasan (fosforilasi bersifat oksidatif). Pada reaksi tahap kelima, suksinat dioksidasi menjadi fumarat oleh enzim suksinat dehidrogenase yang berkaitan dengan flavin adenin dinukleotida (FAD) sebagai koenzimnya. Enzim ini terbuat pada membrane dalam mitokondrion. Dalam reaksi ini FAD berperan sebagai gugus penerima hydrogen. Reaksi tahap keenam merupakan reaksi reversible penambahan satu molekul H2O ke ikatan rangkap fumarat, menghasilkan L-malat, dengan dikatalisis enzim fumarase tanpa koenzim. Enzim ini bersifat stereospesifik, bertindak hanya terhadap bentuk L-stereoisomer dari malat. Dalam reaksi ini fumarase mengkatalisis proses penambahan trans atom H Dan gugus OH ke ikatan rangkap fumarat. Pada reaksi tahap terakhir, siklusasam trikarboksilat, L-malat diooksidasi menjadi oksalasetat oleh enzim L-malat dehidrogenase yang berikatan dengan NAD. Reaksi ini adalah endergonik tetapi laju reaksinya berjalan lancar kekanan. Hal ini dimungkinkan karena reaksi berikutnya, yaitu reaksi kondensasi oksalasetat dengan asetil koenzim-A adalah reaksi eksergonik yang irreversible. Malat dehidrogenase adalah enzim yang berzifat stereospesifik untuk bentuk L-stereoisomer dari malat. 3. Fosforilasi Oksidatif Fosforilasi oksidatif merupakan nama yang diberikan untuk sintesis ATP (fosforilasi) yang terjadi ketika NADH dan FADH2 dioksidasi (sehingga oksidatif) oleh transpor elektron melalui rantai respirasi. Rantai respirasi adalah rangkaian proses transfer elektron hidrogen yang terjadi pada bagian membran dalam mitokodria dengan melibatkan sejumlah enzim. Hasil akhir dari rangkaian proses transfer electron ialah sejumlah energi berbentuk ATP yang diperlukan dalam berbagai aktivitas organisme hidup. Aliran electron dari substrat ke oksigen merupakan sumber energy ATP Pada setiap putaran siklus asam sitrat, 4 pasang atom hydrogen dipindahkan dari isositrat, α-ketoklutarat, suksinat, dan malat melalui aktifitas dehidrogenase spesifik. Atom hidrogen ini, pada beberapa tahap memberikan elektronnya kepada rantai transport electron dan menjadi ion H+ yang terlepas kedalam medium cair. Electron tersebut diangkut disepanjang rantai molekul pembawa electron, sampai electron-elektron ini mencapai oksidase sitokrom, yang menyebabkan pemindahan electron ke oksigen, yakni molekul penerima electron pada organisme aerobic. Pada saat masing-masing atom oksigen menerima 2 elektron dari rantai tersebut, 2 atom H+, yang setara dengan 2H+ yang dilepaskan sebelumnya dari 2 atom hydrogen yang dipindahkan oleh dehidrogenase diambil dari medium cair untuk membentuk H2O. Rantai respirasi terdiri dari serangkaian protein dengan gugus prostetik yang terikat kuat, dan mampu menerima dan memberikan electron. Setiap anggota dapat menerima electron dari anggota sebelumya, dan memindahkan electron ke molekul anggtoa berikutnya, dalam uraian reaksi yang spefisik. Elektron yang masuk ke dalam rantai transport electron yang kaya akan energy, tetapi pada saat electron tersebut melalui rantai, menuju ke oksigen dengan cara setahap demi setahap, electron tersebut kehilangan kandungan energy bebasnya. Banyak dari energy tersebut yang disimpan dalam bentuk ATP oleh mekanisme molekul pada membrane mitokondria sebelah dalam. Pada saat masing-masing pasangan elektron melalui rantai respirasi dari NADH menuju oksigen sintesis 3 molekul ATP dari ADP dan posfat berlaangsung bersama-sama. Ketiga bagian rantai respirasi yang memberikan energy untuk menghasilkan ATP melalui fosforilasi oksidatif disebut sisi penyimpanan energi. Senyawa pengangkut electron selalu berfungsi dalam urutan spesifik Pertama, potensial redoks bakunya secara berturut-turut semakin positif pada arah menuju oksigen, karena electron cendrung mengalir dri sistem elektro negative ke sistem elekro positif menyebabkan penurunan dalam energy bebas. Kedua, setiap rantai anggota respirasi bersifat spesifik bafi senyawa pemberi dan penerima electron tertentu. Sebagai contoh. NADH dapat memindahkan electron ke NADH dehidrogenase, tetapi tidak dapat memindahkan electron ini secara lagsung ke sitokrom b atau ke sitokrom c. Ketiga, kompleks structural protein pengangkut electron yang fungsinya serupa telah dapat diisolasi dari membrane mitokondria. Kompleks I terdiri dari NADH dhidrogenase dan pusat besisulfurnya yang erat berhubungan dalam fungsinya. Kompleks II terdiri dari saksinat dehidrogenase dan pusat besi sulfurnya. Komplek III terdiri dari sitokrom b dan c2, serta pusat besi sulfur spesifik. Sitrokrom a dan a3 bersama-sama menyusun kompleks IV. Ubikuinon merupakan rantai penghubung di antara kompleks I, II, dan III, serta sitokrom c merupakan rantai penghubung diantra kompleks III dan IV. Fosforilasi Oksidatif Rantai respirasi terjadi di dalam mitokondria sebagai pusat tenaga. Di dalam mitokondria inilah sebagian besar peristiwa penangkapan energi yang berasal dari oksidasi respiratorik berlangsung. Sistem respirasi dengan proses pembentukan intermediat berenergi tinggi (ATP) ini dinamakan fosforilasi oksidatif. Fosforilasi oksidatif memungkinkan organisme aerob menangkap energi bebas dari substrat respiratorik dalam proporsi jauh lebih besar daripada organisme anaerob. NADH dan FADH2 yang terbentuk pada reaksi oksidasi dalam glikolisis, reaksi oksidasi asam lemak dan reaksi-reaksi oksidasi dalam siklus asam sitrat merupakan molekul tinggi energi karena masing-masing molekul tersebut mengandung sepasang elektron yang mempunyai potensial transfer tinggi. Bila elektron-elektron ini diberikan pada oksigen molekuler, sejumlah besar energi bebas akan dilepaskan dan dapat digunakan untuk menghasilkan ATP. Fosforilasi oksidatif merupakan proses pembentukan ATP akibat transfer electron dari NADH atau FADH2 kepada O2 melalui serangkaian pengemban electron. Proses ini merupakan sumber utama pembentukan ATP pada organisme air. Sebagai contoh, fosforilasi oksidatif menghasilkan 26 dari 30 molekul ATP yag terbentuk pada oksidasi sempurna glukosa menjadi CO2 dan H2O. Aliran electron dari NADH atau FADH2 ke O2 melalui kompleks-kompleks protein, yang terdapat pada membran dalam mitokondria, akan menyebabkan proton terpompa keluar dari matriks mitokondria. Akibatnya, terbentuk kekuatan daya gerak proton yang terdiri dari gradient pH dan potensial listrik trans membran. Sintesis ATP teradi bila proton mengalir kembali kedalam matriks mitokondria melalui suatu kompleks enzim. Jadi, oksidasi dan fosforilasi terangkai melalui gradient proton melintasi membran dalam mitokondria. Proses fosforilasi oksidatif Organisme kemotrop memperoleh energi bebas dari oksidasi molekul bahan bakar, misalnya glukosa dan asam lemak. Pada organisme aerob, akseptor elektron terakhir adalah oksigen. Namun elektron tidak langsung ditransfer langsung ke oksigen, melainkan dipindah ke pengemban-pengemban khusus antara lain nikotinamida adenin dinukleotida (NAD+) dan flavin adenin dinukleotida (FAD). Pengemban tereduksi ini selanjutnya memindahkan elektron ke oksigen melalui rantai transport elektron yang terdapat pada sisi dalam membran mitokondriaGradien proton yang terbentuk sebagai hasil aliran elektron ini kemudian mendorong sintesis ATP dari ADP dan Pi dengan bantuan enzim ATP sintase. Proses tersebut dinamakan fosforilasi oksidatif. Dalam hal ini energi dipindahkan dari rantai transport elektron ke ATP sintase oleh perpindahan proton melintasi membran. Proses ini dinamakan kemiosmosis. Secara ringkas fosforilasi oksidatif, terdiri atas 5 proses dengan dikatalisis oleh kompleks enzim, masing-masing kompleks I, kompleks II, kompleks III, kompleks IV dan kompleks V Tabel 1 Informasi tentang enzim yang berperan dalam fosforilasi oksidatif Nama Penyusun NADH dehydrogenase (or) Kompleks I Kompleks II NADH-coenzyme Q reductase Succinate dehydrogenase (or) Succinate-coenzyme Q reductase kDa Polypeptides 800 25 140 4 Kompleks III Cytochrome C – coenzyme Q oxidoreductase 250 9-10 Kompleks IV Cytochrome oxidase 170 13 Kompleks V ATP synthase 380 12-14 Dalam fosforilasi oksidatif, daya gerak elektron diubah menjadi daya gerak proton dan kemudian menjadi potensial fosforilasi. Fase pertama adalah peran komplek enzym sebagai pompa proton yaitu NADH-Q reduktase, sitokrom reduktase dan sitokrom oksidase. Komplek-komplek transmembran ini mengandung banyak pusat oksidasi reduksi seperti flavin, kuinon, besi-belerang, heme dan ion tembaga. Fase kedua dilaksanakan oleh ATP sintase, suatu susunan pembentuk ATP yang digerakkan melalui aliran balik proton kedalam matriks mitokondria. Transport electron dan fosforilasi oksidatif terjadi pada Membran Mitokondria sebelah dalam Pada sel eukariotik, hampir semua dehidrogenasa spesifik yang diperlukan pada oksidasi piruvat dan bahan bakar lain melalui siklus asam sitrat terletak pada bagian sebalah dalam mitokondria, yaitu matriks. Molekul pemindahan elektron dari rantai respirasi dan molekul enzim yang melakukan sitesa ATP dari ADP dan fosfat terbenam dalam membran sebelah dalam. Bahan bakar siklus asam sitrat seperti piruvat, harus dipindahkan dari sitosol ( tempat dilakukannya sintesi molekul-molekul tersebut) melalui membran mitokondria kedalam bagian matrik disebelah dalam sebagai tempat aktivitas dehidrogenase. Demikian pula, ADP yang dibentuk dari ATP selama aktivitas yang memerlukan energy didalam sitosol harus dipindahkan didalam metrics mitokondria, untuk mengikat posfat kembali menjadi ATP. ATP baru yang terbentuk harus dikembalikan kesitosol. Sistem transport membran yang khusus pada membrane mitokondria sebah dalam tidak hanya melangsungkan masuknya piruvat dan bahan bakar lain kedalam mitokondria, tetapi juga masuknya posfat dan ADP. Dan keluarnya ATP selama fosforilasi oksidatif. Jadi, membrana mitokondria sebalah dalam merupakan sruktur komplek yang mengandung molekul pembawa electron, sejumlah enzim, dan beberapa sistem transport membran. Yang bersama-sama menyusun sampai 75% atau lebih berat total membrane, sisanya merupakan lipida. Struktur membrane sebelah dalam amat komplek, berliku-liku, dan bersifat mosaic; integritas membran ini penting bagi pembentukan ATP yang menunjang aktivitas hidup. DAFTAR PUSTAKA Anonim. 2014. Siklus Krebs. Di akses pada tanggal 20 Maret https://intanpambua.wordpress.com/2014/03/26/tugas-siklus-krebs/ . 2016 di Anonim. 2010. Dekarboksilasi Oksidatif. Di akses pada tanggal 20 Maret 2016 di http://biologipedia.blogspot.co.id/2010/10/dekarboksilasi-oksidatif.html . Horton, et al. 2002. Principles Of Biochemistry Third Edition. International: Canada. Strayer, L. 1995. Biochemistry. W. H. Freeman And Company: New York. Person Education