Berfungsi dalam katabolisme dan juga anabolisme
advertisement

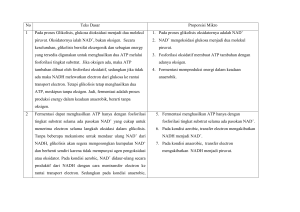
Siklus Krebs dr. Ismawati, M.Biomed Berfungsi dalam katabolisme dan juga anabolisme amfibolik Katabolisme memproduksi molekul berenergi tinggi Anabolisme memproduksi intermedier untuk prekursor biosintesis makromolekul Berbagai daur mengambil senyawa antara dlm siklus kreb berkurang hrs ada mekanisme utk mengganti senyawa antara tadi daur anaplerotik Hubungan antara Glikolisis dan siklus krebs Glycolysis : Glucose to Pyruvate Pyruvate Acetyl CoA The Link Acetyl CoA TCA cycle Metabolisme gliserol dan Asam Lemak Gliserol dan asam lemak berasal dari pemecahan trigliserida. Gliserol memasuki jalur metabolisme diantara glukosa dan piruvat. Kemudian gliserol diubah menjadi glukosa atau piruvat ,selanjutnya menjadi asetil Koa siklus krebs. Asam lemak oksidasi asetil KoA siklus krebs Biosintesis pada siklusTCA pyrimidines pyruvate asparagine aspartate acetyl CoA lipids oxaloacetate citrate glucose phosphoenolpyruvate (PEP) serine, glycine cysteine, tyrosine phenylalanine tryptophan purines malate pyruvate -ketoglutarate succinyl CoA porphyrins, heme, chlorophyll glutamate glutamine proline arginine Siklus asam trikarboksilat = siklus krebs Fungsi siklus ini adalah untuk menyimpan (konservasi) energi dari oksidasi ini dalam bentuk koenzim pemindah-elektron NADH dan FADH2. Hasil keseluruhan dari senyawa yang mengandung energi dalam siklus asam trikarboksilat adalah 3 NADH, 1 FAD(2H) dan 1 GTP. Siklus asam trikarboksilat Reaksi oksidasi reduksi dikatalisis oleh 4 dehidrogenase : isositrat dehidrogenase, -ketoglutarat dehidrogenase, suksinat dehidrogenase dan malat dehidrogenase. Ada 5 koenzim pada siklus asam trikarboksilat : NAD, FAD, tiamin pirofosfat, lipoat dan koenzim asilasi (KoA) . Oksidasi asetil KoA Gugus asetil berfungsi sebagai sumber karbon untuk CO2 dan sumber elektron untuk pemindahan ke NAD+ dan FAD. Merupakan bahan bakar bagi siklus krebs. Oksidasi gugus asetil terjadi dengan melepaskankan elektron sebagai bagian dari ion hidrogen Pembentukan dan oksidasi isositrat Pembentukan sitrat dikatalisis oleh enzim sitrat sintase. Pembentukan isositrat dikatalisis oleh akonitase. Oksidasi isositrat oleh isositrat dehidrogenase membentuk -ketoglutarat dan dihasilkan juga NADH dan CO2. Dekarboksilasi oksidatif -ketoglutarat Dekarboksilasi oksidatif -ketoglutarat membentuk suksinil KoA dikatalisis oleh -ketoglutarat dehidrogenase. Tahap ini menghasilkan energi yang besar, terutama tersimpan dalam NADH dan sebagian kecil disimpan dalam ikatan tioester suksinil koA. Pembentukan suksinat Energi pada ikatan tioester suksinil KoA digunakan untuk membentuk GTP dalam reaksi yang dikatalisis oleh suksinat tiokinase (suksinil KoA sintetase). Reaksi ini merupkan fosforilasi tingkat substrat. Fosforilasi tingkat substrat adalah pembentukan ikatan fosfat berenergi tinggi dimana sebelumnya tidak ada, tanpa menggunakan O2. Oksidasi suksinat menjadi oksaloasetat Oksidasi suksinat oleh suksinat dehidrogenase membentuk fumarat dan energi yang dilepaskan disimpan dalam FAD(2H). Fumarat oleh fumarase diubah menjadi malat. Malat dioksidasi oleh malat dehidrogenase membentuk oksaloasetat dan energi yang dilepaskan disimpan dalam NADH. Perbedaan NAD dan FAD FAD mampu menerima elektron tunggal (H.) dan membentuk zat antara elektron tunggal separuh tereduksi.NAD+ menerima ion hidrida (H:) . Perbedaan struktur kimia kdua koenzim ini menyebabkan peran fisiologis yang berbeda. Bentuk elektron tunggal yang radikal bebas sangat reaktif. Perbedaan NAD dan FAD FAD tetap terikat kuat, kadang-kadang kovalen pada enzimnya selama melepaskan dan menerima elektron, misal enzim suksinat dehidrognase. Enzim ini terdapat pada bagian dalam membran mitokhondria, sedangkan semua enzim lain pada siklus krebs terdapat pada matriks. Perbedaan NAD dan FAD Sebaliknya NAD biasanya berada bebas dalam medium, berikatan dengan dehidrogenase dan menerima elektron, lalu dibebaskan dan melarut NAD+ dan NADH lebih mirip substrat dan produk daripada koenzim. NADH dapat menjadi pengatur fungsi sel. Perbedaan NAD dan FAD NADH yang dihasilkan oleh satu dehidrogenase dapat menghambat dehidrogenase lain apabila elektron tidak digunakan dalam rantai transport elektron untuk sintesis ATP. Pengaturan siklus krebs oleh rasio NADH/NAD+ merupakan bagian dari mekanisme untuk menyelaraskan kecepatan oksidasi bahan bakar terhadap kecepatan penggunaan ATP. Reaksi anaplerotik Agar siklus asam trikarboksilat terus dapat berputar maka jaringan harus menyediakan zat antara 4 karbon yang cukup untuk mengganti keluarnya zat tersebut ke jalur lain, misalnya glukoneogenesis atau sintesis asam lemak. Reaksi yang menyediakan zat antara 4 karbon kepada siklus krebs ini disebut reaksi anaplerotik atau filling up. Reaksi Anaplerotik Ketika produk intermedier TCA digunakan sbg prekursor biosintesis lainnya Konsentrasi intermedier turun memperlambat kecepatan TCA Reaksi anaplerotik Reaksi anaplerotik Salah satu reaksi anaplerotik utama adalah perubahan piruvat dan CO2 menjadi oksaloasetat oleh piruvat karboksilase. Piruvat karboksilase ditemukan dalam konsentrasi tinggi di hati dan jaringan saraf karena jaringan-jaringan ini mengalami effluks zat antara yang konstan dari siklus krebs. Reaksi anaplerotik Seperti sebagian besar jalur anaplerotik, piruvat karboksilase merupakan bagian dari jalur keseluruhan yang bersilangan dengan siklus krebs. Misalnya di hati, piruvat karboksilase merupakan bagian dari jalur glukoneogenik untuk mengubah alanin dan laktat menjadi glukosa. Reaksi anaplerotik Sumber lain zat antara 4 karbon adalah asam amino. Jalur yang mengubah isoleusin, leusin, metionin dan senyawa lain menjadi suksinil KoA adalah jalur anaplerotik utama. Pengaturan Siklus Asam Trikarboksilat Tenaga pendorong utama siklus krebs adalah kecepatan penggunaan ATP. Informasi kecepatan penggunaan ATP melalui : a. Keadaan fosforilasi ATP, seperti tercermin dalam kadar ATP dan ADP b. Keadaan reduksi NAD+, seperti tercermin dalam rasio NADH/NAD+. Pengaturan Siklus Asam Trikarboksilat Didalam sel, bahkan dalam mitokhondria , simpanan adenin dinukleotida (AMP, ADP, dan ATP) dan simpanan NAD (NAD+ dan NADH) relatif konstan. Kecepatan interkonversi adenin dinukleotida sangat bervariasi, juga kecepatan oksidasi reduksi NADH dan NAD+. Pengaturan Siklus Asam Trikarboksilat Pengaturan utama pada 2 tempat : isositrat dehidrogenase dan ketoglutarat dehidrogenase. Isositrat dehidrogenase adalah enzim multisubunit yang secara alosterik diaktifkan oleh ADP dan dihambat oleh NADH. - ADP menyebabkan perubahan konformasi semua subunit enzim sehingga isositrat lebih mudah berikatan. Pengaturan Siklus Asam Trikarboksilat - Perubahan kecil konsentrasi ADP dapat menimbulkan perubahan fluks yang besar. - Perubahan kecil dalam konsentrasi produk (NADH) dan kosubstrat (NAD+) juga mempengaruhi kecepatan enzim. Pengaturan Siklus Asam Trikarboksilat ketoglutarat dehidrogenase dihambat oleh NADH dan suksinil KoA Kedua enzim tersebut diaktifkan oleh Ca2+. Pada saat otot berkontraksi terjadi pelepasan Ca2+ dari retikulum sarkoplasma aktivasi tambahan enzim. A. PengaturanSitrat Sintase Aktifitas sitrat sintase diatur oleh sitrat (inhibitor produk) dan oksaloasetat (subsrat). Bila isositrat dehidrogenase diaktifkan, konsentrasi sitrat menurun, mengurangi inhibisi produk terhadap sitrat sintase dan meningkatkan kecepatannya. Bila rasio NADH/NAD+ menurun, rasio oksaloasetat terhadap malat meningkat. Peningkatan konsentrasi oksaloasetat memacu sitrat sintase. B. Pengaturan allosterik oleh isositrat dehidrogenase Isositrat dehidrogenase secara alosterik diaktifkan oleh ADP dan dihambat oleh NADH. C. Pengaturan oleh alfa ketoglutarat dehidrogenase. Dihambat oleh NADH dan Suksinil KoA, GTP. Diaktifkan oleh Ca. Pengaturan Piruvat dehidrogenase Salah satu sumber utama asetil KoA adalah piruvat yang terbentuk dari glukosa dalam jalur glikolitik. Kompleks piruvat dehidrogenase mengandung subunit pengatur tambahan yaitu piruvat dehidrogenase kinase fosforilasi enzim inaktif. Sub unit pengatur lain : fosfatase. Kompleks piruvat dehidrogenase