peranan poliamin dalam regenerasi tanaman
advertisement

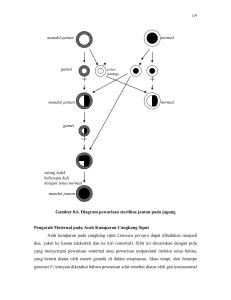
TINJAUAN PUSTAKA Asal dan Taksonomi Padi Padi (Oryza sativa L.), seperti halnya gandum, jagung dan barley termasuk dalam famili Graminae (Poaceae) atau rumput-rumputan. Genus Oryza terdiri atas 23 spesies, 21 diantaranya adalah padi liar (wild relatives) yang memiliki genom diploid atau tetraploid, sedangkan 2 lainnya merupakan padi budidaya. Studi variasi karakter morfologi terhadap 16 spesies padi, telah dilaporkan oleh Morishima dan Oka (1960) yang menyatakan bahwa spesies Oryza dapat dibagi ke dalam tiga grup utama, yaitu (1) Oryza sativa dan kerabat dekatnya, (2) Oryza officinalis dan kerabatnya serta (3) spesies kerabat jauh lainnya. Kush (2000) melakukan klasifikasi ulang terhadap spesies Oryza berdasarkan jumlah kromosom, komposisi genom dan daerah penyebarannya (Tabel 1). Spesies padi liar seperti O. nivara dan O. glaberrima dilaporkan banyak mengandung gen-gen bermanfaat yang dapat ditransfer ke genom padi budidaya (O. sativa). Introgresi gen-gen bermanfaat tersebut dapat dilakukan melalui hibridisasi interspesifik (Brar et al. 1996). Menurut Kush (2000), berdasarkan tingkat kemudahan transfer gen-gen dari spesies liar ke spesies budidaya, spesies padi dapat diklasifikasikan ke dalam tiga gene pool. Gene pool utama terdiri atas sejumlah spesies liar, yaitu O. rufipogon, O. nivara, O. glumaepatula, O. meridionalis, O. breviligulata, O. longistaminata serta dua spesies budidaya, yaitu O. sativa dan O. glaberrima. Kelompok ini memiliki genom AA dan transfer gen antar spesies ini dapat dilakukan melalui persilangan dan metode seleksi tertentu. Gene pool sekunder adalah spesies kelompok O. officinalis komplek. Persilangan antara O. sativa dan spesies-spesies dalam kelompok ini lebih sulit dilakukan dan harus menggunakan bantuan teknik penyelamatan embrio (embryo rescue). Hal ini terjadi karena adanya keterbatasan homologi antara genom AA (O. sativa) dengan genom BB, CC, CCDD, EE dan FF (spesies padi liar), sehingga jumlah gen yang dapat ditransfer terbatas. Gene pool tersier terdiri atas kelompok O. meyeriana, O. ridleyi dan O. schlechteri. Persilangan antara O. sativa dengan kelompok ini sangat sulit dilakukan dan transfer gen antar spesies ini jarang dilakukan. 8 Tabel 1 Klasifikasi Padi menurut Khush (2000) 9 Oryza sativa terdiri atas banyak subspesies dan telah terdistribusi ke seluruh dunia. Dikenal dua grup varietas padi yaitu indica dan japonica, kedua grup ini memiliki banyak perbedaan karakter dibandingkan antar varietas tipikalnya, tetapi variasinya saling overlap. Kedua grup ini telah dikarakterisasi berdasarkan resistensinya, panjang bulu di apikulus dan reaksinya terhadap phenol. Morinaga (1954) dalam Kush (2000) mengusulkan adanya grup ketiga yang terdiri atas varietas bulu dan gundil dari Indonesia. Grup ini disebut javanica, dan Galszmann (1987) dalam Kush (2000) menyatakan bahwa javanica berada di antara grup japonica berdasarkan analisis isozym. Saat ini javanica dikenal sebagai tropical japonica, sedangkan typical japonica disebut sebagai temperate japonica (Gambar 2). Gambar 2 Lintasan evolusi dua species padi budidaya (Kush 2000) Padi Hibrida Padi hibrida di Indonesia dikembangkan dengan sistem 3 galur, yang melibatkan tiga galur tetua meliputi galur mandul jantan sitoplasmik, galur pelestari dan galur pemulih kesuburan. Padi hibrida merupakan generasi F1 hasil persilangan antara galur mandul jantan sebagai tetua betina dengan galur pemulih kesuburan sebagai tetua jantan, sehingga sifat-sifat varietas padi hibrida ditentukan oleh sifat-sifat dari kedua tetuanya. Tetua-tetua yang superior dapat meningkatkan penampilan agronomis dan bobot hasil hibrida turunan berbagai kombinasi persilangan antara galur mandul jantan dan galur pemulih kesuburan 10 (You et al. 2006). Oleh karena itu, untuk mendapatkan varietas padi hibrida yang baik dengan sifat-sifat yang diinginkan seperti daya hasil tinggi (ditunjukkan oleh nilai heterosis yang tinggi), dan tahan terhadap hama dan penyakit utama, maka penelitian padi hibrida diutamakan pada proses perbaikan galur-galur tetua padi hibrida serta proses pembentukan kombinasi persilangan yang menghasilkan produksi dan heterosis tinggi. Padi hibrida dikembangkan dengan memanfaatkan adanya fenomena heterosis yang menyebabkan produksi yang lebih tinggi dibanding galur murni. Beberapa hipotesis telah diajukan untuk menjelaskan dasar genetis dari heterosis. Hipotesis dominansi (Bruce 1910 diacu dalam You et al. 2006) menjelaskan faktor dominan salah satu tetua menutupi gangguan mutasi resesif tetua lain pada populasi F1 heterosigos. Sebaliknya hipotesis overdominan (Shull 1908 diacu dalam You et al. 2006) percaya bahwa heterosigositas pada lokus tunggal lebih superior dibanding yang homosigos. Kedua hipotesis tersebut telah diverifikasi melalui penelitian biologi molekuler (Stuber et al. 1992; Xiao et al. 1995). Hipotesis ketiga menduga bahwa heterosis mungkin akibat terjadinya epistasi antara alel-alel pada lokus yang berbeda (Yu et al. 1997; Goodnight 1999). Beberapa hasil penelitian menunjukkan bahwa epistasis merupakan penyebab utama heterosis secara genetik (Li et al. 2001; Luo et al. 2001). Penelitian dan pengembangan padi hibrida pertama dilakukan oleh peneliti China pada tahun 1964. Galur mandul jantan pertama dikembangkan pada tahun 1972 yang ditemukan pada tanaman mandul jantan steril pada populasi padi liar. Padi hibrida komersial pertama dikenalkan pada tahun 1976. Padi hibrida tersebut memberikan kenaikan hasil sebesar 20% dibandingkan varietas unggul yang ada. Penelitian padi hibrida di daerah tropis dimulai di IRRI, Philipina, pada tahun 1978 dengan mengintroduksi beberapa padi hibrida komersial dari China seperti Shan You 6, Wei You 6 dan Shen You 2. Namun tidak satupun yang menunjukkan kelebihan hasil terhadap varietas pembanding, seperti IR36 dan IR42. Hal ini disebabkan terutama karena varietas-varietas hibrida China tidak mampu beradaptasi terhadap kondisi tropis dan rentan terhadap hama dan penyakit yang ada di daerah tropis. Untuk mengatasi hal tersebut, IRRI melakukan penelitian lebih intensif sehingga berhasil memperoleh galur-galur restorer tropis, dan hibrida hasil persilangan GMJ asal China dengan restorerrestorer tersebut menunjukkan heterosis terhadap IR36 dan IR42. Namun komersialisasi hibrida tersebut masih sulit dilakukan karena terkendala oleh 11 kerentanan GMJ China terhadap hama dan penyakit (Virmani & Kumar 2004). Oleh karena itu IRRI menitikberatkan penelitian padi hibrida terutama pada perakitan galur-galur tetua padi hibrida dan telah banyak menghasilkan kombinasi hibrida unggul yang telah dilepas sebagai varietas di banyak negara, termasuk Indonesia. Suksesnya penelitian dan pengembangan padi hibrida di China dan IRRI, mendorong Indonesia mulai mengintensifkan pemuliaan padi hibrida pada tahun 1998, dengan menguji bahan pemuliaan introduksi dan perakitan galur tetua serta membentuk berbagai kombinasi hibrida sendiri (Badan Litbang 2007). Indonesia, dalam hal ini Balai Besar Penelitian Tanaman Padi (BB Padi) berhasil melepas varietas hibrida pertama pada tahun 2002, yaitu Maro dan Rokan. Kedua varietas tersebut merupakan hasil introduksi dari IRRI. Varietas pertama yang menggunakan tetua jantan lokal Indonesia pertama dilepas pada tahun 2007, yaitu Hipa6 Jete. Sampai tahun 2010, di Indonesia telah dilepas 69 varietas hibrida dan 17 diantaranya dilepas oleh BB Padi, sedangkan varietas lainnya dilepas oleh perusahaan swasta nasional maupun multinasional yang ada di Indonesia seperti PT BISI, Kondo, Bangun Pusaka, KNB Mandiri, Makmur Sejahtera, Bayer, DuPont, Primasid, Tri Usaha Sari Tani, Sang Hyang Seri, Sumber Alam Sutra, Biogene Plantation, Metahelix dan Advanta Indonesia. Semua varietas hibrida yang telah dilepas menunjukkan heterosis pada karakter hasil terhadap IR64 dan Ciherang, tetapi sebagian besar varietas tersebut tidak memiliki ketahanan terhadap hama dan penyakit padi di negara tropis. Selain sistem tiga galur, terdapat dua sistem yang dapat digunakan dalam pengembangan padi hibrida, yaitu sistem dua galur dan sistem satu galur. Sistem satu galur adalah penggunaan sistem apomiktik (tanpa pembuahan), namun sampai saat ini masih dalam skala penelitian. Sistem dua galur melibatkan penggunaan galur mandul jantan TGMS/Temperature-sensitive Genic Male Sterility dan galur fertil. Keuntungan sistem ini adalah (1) tidak memerlukan galur pelestari dalam produksi benih TGMS, (2) semua galur fertil dengan sifatsifat yang baik dapat digunakan sebagai tetua jantan dalam produksi benih hibrida, dan (3) potensi heterosisnya lebih besar dibanding sistem tiga galur (Virmani et al. 2003). Sistem dua galur sebenarnya cocok untuk digunakan di daerah tropis, tetapi anomali iklim yang sering terjadi di daerah tropika seperti Indonesia menjadi kendala. TGMS akan menjadi normal (fertil) ketika temperatur kurang dari 300C 12 (Liu et al. 1998), dan di Indonesia sering sekali terjadi pergantian temperatur yang sangat ekstrim, karena itu sistem tiga galur masih merupakan sistem yang dapat diterapkan. Sistem Mandul Jantan pada Padi Mandul jantan sitoplasmik adalah kondisi dimana tanaman tidak mampu memproduksi polen/tepung sari fungsional. Mandul jantan merupakan karakter yang diwariskan secara maternal, dan biasanya terkait dengan adanya gangguan pada open reading frame (ORF) di genom mitokondria (Hanson & Bentotila 2004). Pada banyak kasus, termasuk padi, kesuburan mandul jantan dapat dipulihkan oleh gen-gen nukleus yang mengkode fertility restoration (gen Rf) yang terdapat pada galur pemulih kesuburan (Restorer/galur R). Sistem mandul jantan sitoplasmik terjadi karena adanya interaksi antara genom nukleus dan mitokondria. Dalam hal ini, sterilitas akibat gen mitokondrial menyebabkan disfungsi sitoplasmik, sedangkan gen-gen nukleus akan menekan disfungsi sitoplasmik tersebut (Ekcard 2006). Penggunaan galur mandul jantan merupakan prasyarat untuk eksploitasi heterosis pada padi. Cytoplasmic-genetic male sterility (CgMS/CMS) telah digunakan secara luas untuk mengembangkan padi hibrida. Pistil galur mandul jantan tumbuh normal dan dapat memproduksi biji bila diserbuki oleh polen normal. Jika faktor genetik yang menginduksi kemandulan tersebut tidak ada dalam sitoplasma maka tanaman menjadi normal (male fertile). Jika terdapat tanaman normal (male fertile) yang memiliki faktor pengendali kemandulan pada sitoplasma dan gen inti yang memulihkan kesuburan bersifat resesif, maka tanaman tersebut dapat mempertahankan sifat mandul jantan. Tanaman atau galur tersebut disebut galur pelestari (maintainer line) yaitu suatu galur yang mempunyai sitoplasma normal tetapi gen inti yang berkaitan dengan pemulihan kesuburan resesif, galur ini berfungsi untuk melestarikan keberadaan galur mandul jantan pasangannya. Adanya gen restorer dominan di nukleus pada suatu galur mengakibatkan galur tersebut mampu memulihkan kesuburan pada hibrida hasil persilangan antara galur tersebut dan galur CMS. Galur ini disebut sebagai galur pemulih kesuburan atau restorer. Pada sistem tiga galur, galur mandul jantan selalu diperbanyak dengan cara menyilangkannya dengan galur pelestari, baik secara manual (hand 13 crossing) untuk produksi benih skala kecil, maupun melalui persilangan alami pada plot terisolasi untuk produksi benih dalam skala besar. Galur mandul jantan dan galur pelestari selalu sama secara morfologi, hanya galur mandul jantan steril sedangkan galur pelestari fertil. Namun kadang-kadang, kedua galur tersebut juga menunjukkan beberapa sifat morfologi dan agronomis yang berbeda karena dipengaruhi oleh faktor-faktor yang ada di dalam sitoplasma yang menginduksi mandul jantan (Virmani et al. 1997). Restorer yang mempunyai gen pemulih kesuburan (Rf) dominan, ketika disilangkan dengan GMJ akan menghasilkan F1 hibrida yang fertil. Perbanyakan benih galur pelestari dan pemulih kesuburan dilakukan seperti perbanyakan padi biasa, karena kedua galur ini fertil dan mempunyai bunga sempurna. Gambar 3 Skema sistem galur mandul jantan pada padi Galur mandul jantan diklasifikasikan berdasarkan beberapa kriteria, antara lain (1) berdasarkan perilaku genetik dari gen ms, GMJ dibedakan menjadi dua tipe yaitu GMJ sporofitik dan gametofitik. Pada GMJ sporofitik, sterilitas atau fertilitas polen ditentukan oleh genotipe dari sporofit sedangkan genotipe gametofit (polen) tidak berpengaruh sama sekali. Beberapa GMJ yang tergolong tipe ini antara lain wild-abortive (WA) dan Gambiaca (Gam). Pada kedua GMJ tersebut, gugurnya polen terjadi pada fase awal perkembangan mikrospora. Fertilitas GMJ gametofitik secara langsung ditentukan oleh genotipe gametofit (polen) saja tanpa dipengaruhi oleh genotipe sporofit. Pengguguran polen biasanya terjadi di fase akhir perkembangan mikrospora. GMJ dengan 14 sitoplasma Boro-type termasuk dalam tipe ini; (2) berdasarkan pola pelestarianpemulih kesuburan dari GMJ, terdapat tiga tipe galur mandul jantan yaitu WA, Honglian dan Boro type (BT); dan (3) berdasarkan morfologi polen, GMJ digolongkan ke dalam tipe typical abortion, spherical abortion, dan stained abortion. GMJ tipe typical abortion mempunyai bentuk polen tidak beraturan dan pengguguran polen biasanya terjadi pada fase uninukleat, sedangkan tipe spherical abortion mempunyai polen berbentuk agak lonjong dan polen gugur kira-kira saat fase binukleat. Terakhir, bentuk polen tipe stained abortion juga agak lonjong tetapi agak lebih kecil dibanding polen normal dan polen gugur saat fase trinukleat (Yuan et al. 2003). Galur mandul jantan yang berkembang di Indonesia hampir seluruhnya mempunyai sumber sitoplasma yang sama, yaitu Wild Abortive (WA). Apabila galur ini digunakan secara terus-menerus, ada kekhawatiran akan mengakibatkan kerapuhan genetik padi hibrida terhadap hama dan penyakit padi yang kemudian akan menyebabkan ledakan populasi hama dan penyakit, sama seperti yang pernah terjadi pada jagung hibrida (kasus Texas CMS). IRRI telah mengembangkan beberapa galur mandul jantan dengan sumber sitoplasma yang berbeda antara lain Gambiaca, Dissi, ARC, Kalinga, Hong Lian, Indonesian Paddy (IP) dan beberapa sumber sitoplasma dari padi liar seperti Oryza rufipogon dan O. perennis. Namun di Indonesia sumber-sumber sitoplasma di atas belum digunakan dalam pembentukan dan produksi padi hibrida, karena belum teridentifikasi galur pemulih kesuburan yang cocok berkombinasi dengan GMJ dengan sumber-sumber sitoplasma di atas. Uji Daya Gabung Keberhasilan setiap program pemuliaan tanaman tergantung pada pemilihan genotipe tetua yang tepat untuk program persilangan, disesuaikan dengan target pemuliaan. Studi daya gabung tetua-tetua yang digunakan dalam program pemuliaan memberikan informasi yang dapat membantu pemulia melakukan seleksi terhadap tetua yang lebih baik untuk digunakan dalam program pemuliaan, sehingga program tersebut menjadi lebih efektif. Daya gabung umum (DGU) dan daya gabung khusus (DGK) dapat menjadi indikator penting dalam evaluasi nilai potensial masing-masing galur tetua dalam kombinasi hibrida. Daya gabung merupakan ukuran kemampuan suatu galur atau tetua yang bila disilangkan dengan galur lain akan menghasilkan hibrida 15 dengan penampilan superior. Nilai masing-masing galur terletak pada kemampuannya untuk menghasilkan keturunan unggul bila dikombinasikan dengan galur-galur lain (Allard 1960). Daya gabung umum (general combining ability) adalah nilai rata-rata galurgalur dalam seluruh kombinasi persilangan bila disilangkan dengan galur-galur lain. Daya gabung umum (DGU) menggambarkan peranan dan interaksi yang kuat antar efek genetik aditif pada populasi dasar, sedangkan efek daya gabung khusus memberi ilustrasi mengenai variasi non-aditif (Falconer 1983). Nilai DGU yang tinggi menunjukkan tetua atau galur yang bersangkutan mempunyai kemampuan berkombinasi dengan baik, sedangkan nilai DGU yang rendah menunjukkan bahwa tetua tersebut mempunyai kemampuan berkombinasi yang lebih buruk dibanding tetua lain. Nilai DGU dapat positif atau negatif, tergantung pada karakter yang diamati dan cara menilainya. Nilai daya gabung dapat memberikan informasi mengenai kombinasi-kombinasi yang dapat menghasilkan turunan yang berpotensi hasil tinggi, yang ditunjukkan oleh diperolehnya heterosis dan nilai daya gabung yang besar. Daya gabung khusus (specific combining abillity atau DGK) adalah penampilan kombinasi pasangan persilangan tertentu. Nilai daya gabung khusus baik apabila nilai pasangan persilangan tertentu lebih tinggi dibanding nilai ratarata keseluruhan persilangan yang terlibat (Poehlman & Sleper 1990). Persilangan-persilangan yang memiliki nilai daya gabung khusus tinggi sangat berpotensi untuk dieksploitasi efek heterosisnya dan dapat dikembangkan sebagai varietas hibrida. Evaluasi persilangan antar galur inbred merupakan langkah penting dalam pengembangan padi hibrida. Secara ideal, proses ini sebaiknya dilakukan melalui evaluasi terhadap semua kemungkinan persilangan yang terjadi, sehingga dapat diketahui kombinasi yang sangat sesuai untuk disilangkan agar dapat menghasilkan suatu varietas yang baik. Analisis daya gabung yang banyak digunakan adalah lini x tester dan diallel (Singh & Chaudary 1979). Analisis dialel umum dilakukan menggunakan populasi persilangan galur-galur murni, dan telah banyak diterapkan pada banyak tanaman. Analisis daya gabung umum dan khusus yang melibatkan galur mandul jantan pada padi lebih umum dilakukan menggunakan metode lini x tester (Swamy et al. 2003; Sao & Motiramani 2006; Tiwari et al. 2011).