Daya tahan otot - Universitas Sumatera Utara
advertisement

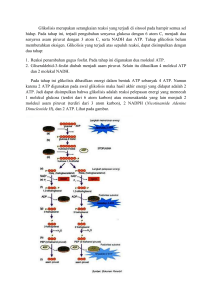
BAB 2 TINJAUAN PUSTAKA 2.1. MUSCULAR ENDURANCE (Daya tahan otot) Kemampuan otot merupakan penentu kesehatan dan penyakit. Daya tahan otot (Muscular endurance), kekuatan (strength), kelelahan ditentukan oleh berbagai factor, termasuk transportasi substrat untuk produksi ATP, kemampuan mitokondria terhadap pembakaran nutrien dan komposisi penyebab kontraktilitas. Walaupun faktor genetik juga menentukan fenotip otot, faktor fisiologi seperti; aktivitas fisik dan olahraga juga berkontribusi selama periode postnatal yang berdampak pada komposisi tipe serabut otot, biogenesis mitokondria dan jalur metabolik energi (Gan et al, 2013). Muscular endurance adalah kemampuan otot untuk melakukan atau mempertahankan kontraksi secara berulang dalam waktu tertentu. Muscular endurance berhubungan erat dengan kekuatan otot (Prentice, 2012). Muscular endurance akan mengalami penurunan jika dalam waktu dua minggu tidak melakukan aktivitas. Pada saat ini belum ada penelitian yang menyatakan apakah penurunan performa ini disebabkan oleh perubahan pada otot atau perubahan kemampuan kardiovaskuler. Adaptasi otot lokal terjadi selama periode tidak beraktivitas. Kita ketahui pada kasus postoperasi setelah satu atau dua minggu mengalami immobilisasi, aktivitas enzim oksidatif, seperti; succinate dehydrogenase (SDH) dan cytochrome oxidase menurun 40% hingga 60%. Penurunan aktivitas enzim oksidatif seharusnya diharapkan untuk memperbaiki Muscular endurance, dan hal ini sebagian besar berhubungan dengan kemampuan enduransi submaksimal dibandingkan dengan kemampuan maksimal aerobik atau VO2max. Pada saat atlit berhenti latihan, enzim glikolitik otot, seperti phosphorylase dan phosphofructokinase mengalami perubahan sedikit hingga total yang berlangsung selama 4 minggu. Perubahan lain pada otot selama tidak latihan adalah kandungan glikogen. Otot enduransi cenderung meningkatkan penyimpanan glikogen. Tetapi tidak latihan selama 4 minggu menyebabkan penurunan glikogen otot 40%. Muscular endurance ditentukan oleh berbagai faktor, termasuk diantaranya adalah kemamampuan oksidasi mitokondria, sintesis ATP, tipe serabut otot dan vaskularisasi (Yan et al, 2011; Gan et al, 2013). Serabut otot tipe I (slow-twich) kaya akan mitokondria sehingga 5 Universitas Sumatera Utara lebih dominan oksidasi asam lemak pada mitokondria untuk produksi ATP. Sedangkan serabut otot tipe II (fast-twich) lebih dominan pada glukosa sebagai substrat energi dan disubklasifikasi menjadi tipe IIa, IIx atau IIb pada hewan didasarkan pada tipe isoform myosin heavy chain (MHC). Sensitifitas insulin dan transportasi glukosa oleh insulin berhubungan positif dengan proporsi serabut otot tipe I. Training enduransi meningkatkan proporsi serabut oksidatif secara parallel dengan respon biogenik mitokondria berhubungan dengan peningkatan latihan fisik, oksidasi asam lemak dan glukosa dan pemakaian energi (Gan et al, 2013) Permainan sepak bola merupakan salah satu permainan yang membutuhkan daya tahan dalam jangka waktu yang cukup lama. Daya tahan penting dalam permainan sepak bola sebab dalam jangka waktu 90 menit bahkan lebih, seorang pemain melakukan kegiatan fisik yang terus menerus dengan berbagai bentuk gerakan seperti berlari, melompat, meluncur (sliding), body charge dan sebagainya yang jelas memerlukan daya tahan yang tinggi. 2.1.1. Tipe serabut otot Tidak semua serabut otot sama. Suatu otot skeletal terdiri dari serabut serabut yang berbeda kecepatan memendeknya dan kemampuan untuk menghasilkan kekuatan maksimal; serabut tipe I (slow-twich) dan serabut tipe II (fast-twich). Serabut tipe I kira-kira 110 ms untuk mencapai puncak tegangan ketika dirangsang. Serabut tipe II dapat mencapai puncak tegangan sekitar 50 ms. Walaupun hanya satu bentuk serabut tipe I yang telah diidentifikasi, serabut tipe II dapat diklasifikasikan lagi. Pada manusia, terdapat dua bentuk utama serabut tipe II yaitu fast-twich tipe a (tipe IIa) dan fast-twich type x (tipe IIx). Serabut tipe IIx Serabut tipe I bernoktah hitam; serabut tipe IIa tidak bernoktah dan tampak putih dan serabut tipe IIx tampak abu-abu (gray) (Kenny et al,2012). 2.1.2. Karakteristik serabut otot tipe I dan tipe II a. ATPase Serabut tipe I dan tipe II berbeda kecepatan kontraksinya. Perbedaan ini disebabkan bentuk myosin ATPase. Myosin ATPase merupakan enzim pengurai ATP untuk melepaskan energi yang digunakan untuk berkontraksi. Serabut tipe I memiliki bentuk myosin ATPase yang lambat (slow), sedangkan serabut tipe II memiliki yang lebih cepat. Sebagai respon terhadap rangsangan neural, ATP diuraikan lebih cepat pada serabut otot tipe II dibandingkan pada serabut tipe I. sehingga cross-bridge cycle lebih cepat pada serabut tipe II. 6 Universitas Sumatera Utara b. Reticulum sarkoplasmik Serabut tipe II memiliki perkembangan reticulum sarcoplasmic yang lebih tinggi dibandingkan dengan serabut tipe I. Serabut tipe II lebih cepat mengirimkan kalsium kedalam sel otot yang dirangsang. Kemampuan ini menyebabkan kecepatan kontraksi (Vo) lebih cepat pada serabut tipe II. Secara rata-rata, serabut tipe II pada manusia memiliki Vo 5-6 kali lebih cepat dibandingkan pada serabut tipe I. Walaupun jumlah kekuatan (Po) yang dihasilkan oleh serabut tipe II dan serabut tipe I dengan diameter yang sama adalah sama, jumlah tenaga yang dihasilkan serabut tipe II adalah tiga sampai lima kali lebih besar dibandingkan serabut tipe I karena kecepatan pemendekan yang lebih cepat. Hai ini menjelaskan mengapa seseorang dengan serabut tipe II lebih dominan terdapat pada otot kaki cenderung sprinter lebih baik dibandingkan seseorang dengan persentase serabut tipe I. c. Unit motor Motor unit disusun oleh single α-motor neuron dan innervasi serabut otot. Single α-motor neuron menentukan apakah serabut tipe I atau serabut tipe II. Single α-motor neuron pada motor unit tipe I memiliki lebih kecil sel bodi dan secara khusus diinervasi ≤300 serabut otot. Sementara α-motor neuron pada motor unit tipe II memiliki sel bodi lebih besar dan diinervasi ≥300 serabut otot. Perbedaan ukuran unit motor menyatakan bahwa single tipe I α-motor neuron perangsangan serabut, jauh lebih sedikit serabut otot berkontraksi dibandingkan single tipe II α-motor neuron merangsang serabutnya. Sehingga, serabut tipe II untuk mencapai tegangan puncak lebih cepat dan secara kolektif menghasilkan tenaga yang lebih dibandingkan serabut tipe I. 2.1.3. Distribusi serabut otot Persentase serabut tipe I dan tipe II tidak sama diseluruh otot tubuh. Umumnya, otot pada lengan dan kaki memiliki serabut hampir sama komposisinya pada setiap orang. Pada atlit enduransi terdapat dominan serabut tipe I pada otot kaki dan juga dengan persentase yang tinggi pada otot lengannya. Hubungan yang mirip juga terdapat pada serabut tipe II. Tetapi terdapat beberapa pengecualian, otot soleus (dibawah otot gastrocnemius di betis), terdapat persentase serabut tipe I yang cukup tinggi pada setiap orang. 7 Universitas Sumatera Utara 2.1.4. Serabut otot dan latihan fisik Karena terdapat perbedaan serabut tipe I dan tipe II, juga terdapat fungsi yang berbeda pada tipe serabut otot ketika seseorang beraktivitas fisik secara aktif. a. Serabut tipe I Umumnya, serabut tipe I berhubungan dengan tingginya kemampuan aerobic enduransi. Aerobic berarti “tersedianya oksigen” sehingga oksidasi adalah proses aerobic. Serabut tipe I sangat efisien dalam memproduksi ATP berasal dari oksidasi karbohidrat dan lemak. ATP dibutuhkan untuk menyediakan energi yang dibutuhkan serabut otot berkontraksi dan relaksasi. Pada saat oksidasi berlangsung, serabut tipe I secara terus menerus memproduksi ATP, menyebabkan serabut tetap aktif. Kemampuan untuk mempertahankan aktivitas muscular berlangsung dalam waktu yang lebih lama yang disebut muscular endurance, sehingga serabut tipe I memiliki enduransi aerobic lebih tinggi. Akibat kondisi tersebut, mereka sering direkrut selama pertandingan endurasi dengan intensitas rendah (seperti; marathon) dan pada saat aktivitas harian yang membutuhkan kekuatan otot yang rendah (seperti; berjalan) b. Serabut tipe II Serabut tipe II memliki enduransi aerobic yang sangat kurang dibandingkan serabut tipe I. Tipe serabut tersebut terjadi secara anaerobik (tanpa oksigen). Hal ini berarti tidak tersedianya oksigen, ATP dihasilkan melalui jalur anaerobik. Motor unit tipe IIa menghasilkan tenaga lebih besar dibandingkan motor unit tipe I, tetapi motor unit tipe IIa juga mengalami kelelahan lebih mudah karena keterbatasan enduransi. Sehingga serabut tipe IIa secara primer digunakan dalam waktu singkat, pertandingan enduransi dengan intensitas lebih tinggi, seperti lari atau renang 400 m. (Katch et al, 2011) 2.1.5. Metabolisme pada otot Energi dilepaskan ketika elemen ikatan kimia secara bersama-sama membentuk molekul diuraikan. Substrat secara utama disusun oleh karbon, hydrogen, oksigen dan pada protein nitrogen. Ikatan molecular sebagai tempat elemen secara relative lemah dan selanjutnya menghasilkan sedikit energi ketika diuraikan. Akibatnya, nutrient tidak digunakan secara langsung untuk aktivitas selluler. Energi pada ikatan molecular nutrient secara kimia 8 Universitas Sumatera Utara dilepaskan didalam sel dan kemudian disimpan dalam bentuk komponen energi tingkat tinggi yang disebut Adenosine triphosphate (ATP). Pada saat istirahat tubuh membutuhkan perolehan yang dari pemecahan karbohidrat dan lemak. Protein memiliki fungsi yang sangat penting sebagai enzim yang membantu reaksi kimia dan sebagai pembangun struktur tetapi biasanya menghasilkan energy yang sedikit dari metabolisme. Selama kondisi berat, kontraksi otot dalam waktu singkat, karbohidrat lebih digunakan dengan lebih sedikit menggunakan lemak untuk menghasilkan ATP. Dalam waktu yang lama, latihan fisik intensitas rendah menggunakan karbohidrat dan lemak untuk mempertahankan produksi energi. a. Karbohidrat Jumlah karbohidrat yang digunakan dalam aktivitas fisik dihubungkan dengan ketersediaan karbohidrat dan metabolisme karbohidrat pada otot. Karbohidrat secara utama diubah menjadi enam karbon gula sederhana, glucosa, sebuah monosaccharide (satu unit gula) dan ditransportasikan kedalam darah menuju seluruh jaringan tubuh. Saat kondisi istirahat, karbohidrat yang dimakan disimpan didalam otot dan hati dalam bentuk yang lebih kompleks yaitu polysaccharide (molekul gula kompleks), glycogen. Glycogen disimpan didalam sitoplasma sel otot hingga sel menggunakan untuk membentuk ATP. Glycogen juga disimpan di hati dan diubah kembali menjadi glukosa jika dibutuhkan dan kemudian ditransportasikan oleh darah menuju jaringan yang aktif untuk dimetabolisme. Hati dan otot menyimpan glycogen secara terbatas dan dapat habis dalam waktu yang berkepanjangan, latihan fisik berat, khususnya jika diet jumlah karbohidrat tidak mencukupi. Selanjutnya akan diharapkan sangat besar pada sumber diet karbohidrat dan gula untuk selanjutnya mengisi tempat cadangan karbohidrat. Tanpa pasokan karbohidrat yang cukup, otot dapat mengalami kehilangan sumber energi utama. Selanjutnya, karbohidrat adalah hanya sumber energi yang digunakan oleh jaringan otak, sehingga penurunan karbohidrat menyebabkan dampak negative pada kognitif. 9 Universitas Sumatera Utara Gambar 2.1 Metabolisme Seluler b. Lemak Lemak menghasilkan sangat besar energi yang digunakan pada saat latihan fisik intensitas lama dan rendah. Tubuh menyimpan potensial energi dalam bentuk lemak dengan substansial lebih besar dibandingkan cadangan karbohidrat. Pada usia dewasa pertengahan dengan lemak tubuh yang lebih (jaringan adiposa), penyimpanan lemak dua kali lebih besar dibandingkan dengan karbohidrat. Tetapi penggunaan lemak untuk metabolisme lebih sedikit karena harus terlebih dahulu dirubah bentuknya kompleksnya, triglyceride, menjadi komponen dasarnya yaitu glycerol dan asam lemak bebas (FFAs). Hanya FFAs yang digunakan untuk membentuk ATP Pada hakekatnya energi yang dihasilkan dari pemecahan satu gram lemak (9.4 kcal/g) dibandingkan dengan jumlah karbohidrat satu gram (4.1 kcal/g). Meskipun demikian jumlah energi yang dihasilkan dari lemak lebih lambat untuk memenuhi kebutuhan energi pada aktivitas otot berat. Jenis lemak lainnya ditemukan pada tubuh yang berfungsi sebagai nonenergi. Phospholipids adalah komponen struktur pada membrane sel dan melindungi lapisan pada beberapa sel saraf besar. Steroid terdapat pada membrane sel dan berfungsi sebagai hormone atau sebagai pembangun hormone seperti estrogen dan testosterone. 10 Universitas Sumatera Utara 2.1.6. Sistem dasar energi Sel dapat menyimpan sangat terbatas jumlah ATP dan secara tetap menghasilkan ATP baru untuk menyediakan kebutuhan energi pada semua metabolisme sellular termasuk kontraksi otot. Sel menghasilkan ATP melalui salah satu atau kombinasi dari tiga jalur metabolik. a. Sistem ATP-PCr Sistem energi yang paling dasar adalah sistem ATP-PCr. Untuk menyimpan energi dalam jumlah sangat sedikit, sel harus terdapat molekul fosfat energi tingkat tinggi yang disebut phosphocreatine atau PCr (Creatine phosphate). Jalur sederhana ini melibatkan donasi Pi dari PCr ke ADP untuk membentuk ATP. Tidak sama dengan ATP bebas yang terbatas pada sel, energi dilepaskan melalui pemecahan pada PCr yang tidak secara langsung digunakan untuk aktivitas selular. Malahan, proses tersebut membentuk kembali ATP untuk mempertahankan secara relative konstan suplai pada kondisi istirahat. Energi yang dilepaskan dari PCr dikatalisasi oleh enzim creatine kinase, yang bekerja pada PCr untuk menguraikan Pi dari creatine. Energi yang dilepaskan dapat digunakan untuk menambah sebuah molekul Pi menjadi sebuah molekul ADP, membentuk ATP. Sebagai energi yang dilepaskan dari ATP melalui pemecahan sekelompok fosfat, sel dapat mencegah penurunan ATP melalui pemecahan PCr, menghasilkan energi dan Pi untuk membentuk kembali ATP dari ADP. Berdasarkan pada prinsip mekanisme umpan balik negative (negative feedback) dan terbatasnya jumlah enzim, aktivitas creatine kinase meningkat pada saat konsentrasi ADP atau Pi meningkat. Pada saat latihan fisik berat, jumlah ATP yang sedikit pada otot diuraikan dengan segera untuk persediaan energi, hasil yang diproduksi adalah ADP dan Pi. Peningkatan konsentrasi ADP meningkatkan aktivitas creatine kinase dan PCr dikatabolisme untuk membentuk ATP tambahan. Pada saat latihan fisik berlangsung dan ATP tambahan dihasilkan melalui dua sisyem energi, yaitu; sistem glikolitik dan sistem oxidative, sehingga aktivitas creatine kinase dihambat. Proses pemecahan PCr menyebabkan pembentukan ATP secara cepat dan dapat berhasil tanpa disetiap struktur didalam sel. Sistem ATP-PCr diklasifikasikan sebagai tingkat metabolisme substrat. Walaupun dapat terjadi dengan adanya oksigen, prosesnya tidak membutuhkan oksigen. Pada kondisi 11 Universitas Sumatera Utara kelelahan, kadar ATP dan PCr adalah rendah dan tidak dapat menghasilkan energi untuk lebih lanjut kontraksi dan relaksasi otot. Selanjutnya kemampuan untuk mempertahankan level ATP dengan energi dari PCr menjadi terbatas. Kombinasi penyimpanan ATP dan PCr dapat mempertahankan kebutuhan energi otot hanya selama 3-15 detik selama sprint. Pada saat itu, otot harus bergantung pada proses lainnya untuk pembentukan ATP, yaitu jalur glikolitik dan jalur oxidative. Gambar 2.2 Sistem ATP-PCr b. Sistem Glikolitik Sistem ATP-PCr memiliki keterbatasan untuk menghasilkan ATP bagi energi., hanya berlangsung beberapa detik. Metode kedua produksi ATP melibatkan pelepasan energi melalui pemecahan (lysis) pada glukosa. Sistem ini disebut sistem glikolitik karena memerlukan glikolisis, pemecahan glukosa melalui jalur yang melibatkan sekuensi enzim glikolitik. Glikolisis merupakan jalur yang lebih kompleks dibandingkan sistem ATP-PCr. Jumlah glukosa sekita 99% dari total gula bersirkulasi didalam darah. Glukosa darah berasal dari digesti karbohidrat dan pemecahan glikogen hati. Glikogen disintesis dari glukosa melalui proses disebut glycogenesis dan disimpan didalam hati atau didalam otot hingga dibutuhkan. Pada suatu saat, glikogen dipecah menjadi glucose-1-phosphate, kemudian memasuki jalur glikolisis, proses ini disebut glycogenolysis. 12 Universitas Sumatera Utara Gambar 2.3 Produksi energi pada proses Glikolisis Sebelum glukosa atau glikogen digunakan untuk menghasilkan energi, terlebih dahulu dirubah menjadi komponen yang disebut glucose-6-phosphate. Tujuan proses glikolisis adalah untuk melepaskan ATP, konversi sebuah molekul glukosa menjadi glucose-6-phosphate membutuhkan pasokan satu molekul ATP. Pada konversi glikogen, glucose-6-phosphate dibentuk dari glucose-1-phosphate tanpa membutuhkan energi. Proses glikolisis secara teknis dimulai pada saat glucose-6-phosphate dibentuk.Glikolisis membutuhkan 10-12 reaksi enzimatik untuk memecah glikogen menjadi asam piruvat, yang selanjutnya dikonversi 13 Universitas Sumatera Utara menjadi asam laktat. Pada semua langkah didalam jalur nya dan semua enzim dilibatkan operasinya didalam sitoplasma sel. Hasil bersih yang diperoleh dari proses ini adalah 3 mole (mol) ATP yang dibentuk dari setiap mole glikogen yang dipecah. Jika glukosa digunakan berasal dari glikogen, hasil bersih hanya 2 mol ATP karena I mol digunakan untuk konversi glukosa menjadi glucose-6phosphate. Sistem energi seyogianya tidak menghasilkan jumlah besar ATP. Meskipun keterbatasan ini, kombinasi aksi pada ATP-PCr dan sistem glikolisis menyediakan bagi otot tenaga ketika suplai oksigen terbatas. Kedua sistem ini dominan terjadi selama menit-menit awal latihan fisik tinggi. Keterbatasan utama lainnya pada glikolisis anaerobic adalah disebabkan akumulasi asam laktat pada otot dan cairan tubuh. Glikolisis menghasilkan asam piruvat. Proses ini tidak membutuhkan oksigen, tetapi adanya oksigen menentukan nasib asam piruvat. Tanpa adanya oksigen, asam piruvat dikonvensi secara langsung menjadi asam laktat, asam dengan formula kimia C 3H6O3. Glikolisis anaerobic menyebabkan asam laktat, tetapi secara cepat disosiasi dan laktat dibentuk. Pada pertandingan sprint sekitar 1 atau 2 menit, kebutuhan akan sistem glikolitik adalah tinggi, konsentrasi asam laktat dapat meningkat mulai dari kondisi istirahat sekita 1 mmol/kg pada otot menjadi lebih dari 25 mmol/kg. kondisi keasaman pada serabut otot menghambat pemecahan glikogen karena menganggung fungsi enzim glikolitik. Dampak lainnya adalah, asam menurunkan kemampuan ikatan serabut dengan kalsium dan mengakibatkan gangguan kontraksi otot. Terbatasnya enzim pada jalur glikolitik yaitu phosphofructokinase atau PFK, sama seperti sebagian besar keterbatasan enzim. PFK mengkatalis pada langkah awal jalur, konversi fructose-6-phosphate menjadi fructose-1,6-biphosphate. Peningkatan konsentrasi ADP dan Pi meningkatkan aktivitas PFK dan selanjutnya mempercepat glikolisis, saat konsentrasi ATP meningkat memperlambat glikolisis melalui penghambatan PFK. Pada situasi selanjutnya, jalur glikolisis akan memasuki siklus Krebs untuk menambah produksi energi ketika tersedia oksigen, produk siklus Krebs, khususnya adalah sitrat dan ion hydrogen, sepertinya menyebabkan penghambatan PFK. 14 Universitas Sumatera Utara Jumlah penggunaan energi pada serabut otot pada saat latihan fisik dapat mencapai 200 kali lebih besar dibandingkan pada saat istirahat. Sistem ATP-PCr dan sistem glikolitik secara tersendiri tidak dapat memenuhi seluruh kebutuhan energi. Lebih lanjutnya, kedua sisten tersebut tidak mampu memenuhi kebutuhan energi untuk seluruh aktivitas kurang dari 2 menit. Latihan fisik yang berlangsung lama membutuhkan sistem energi yang ketiga, sistem oxidatif. c. Sistem Oxidatif Sistem akhir pada produksi energi seluler adalah sistem oxidative. Proses ini paling kompleks dari ketiga sisyem energi. Proses ini melalui pemecahan substrat dengan menambahkan oksigen untuk menghasilkan energi yang disebut Cellular respiration. Karena oksigen dibutuhkan, proses ini disebut proses aerobic. Tidak seperti kondisi anaerobic dalam memproduksi ATP yang terjadi didalam sitoplasma sel, oksidasi produksi ATP terjadi didalam organel sel khusus, yaitu mitochondria. Didalam otot, berbatasan dengan myofibril dan juga banyak terdapat pada sarkoplasma. Otot membutuhkan suplai energi yang tetap untuk menghasilkan tenaga selama aktivitas berlangsung lama. Tidak seperti kondisi anaerobik dalam produksi ATP, sistem oksidatif lambat turun; tetapi memiliki kemampuan memproduksi energi dalam jumlah besar, sehingga metabolisme aerobik merupakan metode utama produksi energi selama aktivitas enduransi. Kondisi ini sangat bergantung kepada sistem kardiovaskuler dan sistem respirasi untuk mengirimkan oksigen ke otot yang aktif. Produksi energi secara oksidatif berasal dari karbohidrat (dimulai dengan glikolisis) atau lemak. 1. Oksidasi karbohidrat Produksi ATP pada oksidasi dari karbohidrat nelibatkan tiga proses, yaitu: a) Glikolisis Pada metabolisme karbohidrat, glikolisis berperan pada produksi ATP secara aerobik dan anaerobik. Proses glikolisis adalah tanpa menghiraukan apakah oksigen tersedia. Tersedianya oksigen menentukan nasib akhir produk, asam piruvat. Glikolisis anaerobik menghasilkan asam laktat dan hanya 3 mol ATP per mol glikogen, atau 2 mol ATP per mol glukosa. Tersedianya oksigen, 15 Universitas Sumatera Utara asam piruvat dirubah menjadi sebuah senyawa, disebut acetyl coenzyme A (acetyl CoA) Gambar 2.4 Produksi energi pada sistem metabolik b) Siklus Krebs Setelah dibentuk, acetyl CoA memasuki siklus Krebs (siklus asam sitrat atau siklus asam trisiklik), sebuah rangkaian kompleks pada reaksi kimia yang menyebabkan oksidasi komplit acetyl CoA. Setiap molekul glukosa memasuki jalur glikolitik, dua molekul asam piruvat akan dibentuk. Selanjutnya setiap molekul glukosa akam mulai proses produksi energi dengan tersedianya oksigen pada siklus Krebs. Konversi succinyl CoA menjadi succinate pada siklus Krebs menyebabkan pembentukan guanosine triphosphate atau GTP, senyawa energi tingkat tinggi mirip dengan ATP. GTP kemudian transfer sebuah Pi menjadi ADP untuk membentuk ATP. Dihasilkannya dua ATP (per molekul glukosa) dibentuk oleh level substrat posforilasi. Pada akhir siklus Krebs, tambahan dua mol ATP dihasilkan secara langsung, dan karbohidrat telah dipecah menjadi karbondioksida dan hydrogen. 16 Universitas Sumatera Utara Seperti jalur metabolic lainnya melibatkan metabolisme energi, enzim siklus Krebs diatur oleh umpan balik negative (Negative Feedback) pada beberapa langkah didalam siklus. Enzim pada siklus Krebs yang membatasi jumlahnya adalah isocitrate dehydrogenase, seperti PFK, yang dihambat oleh ATP dan diaktifkan oleh ADP dan Pi, sebagai rantai transport electron. Karena kontraksi otot bergantung kepada ketersediaan kalsium didalam sel, kalsium yang berlebihan juga merangsang enzim isocitrate dehydrogenase. Gambar 2.5 Siklus Kreb’s c) Rantai Transport Elektron (ETC) Selama glikolisis, ion hydrogen dilepaskan ketika glukosa dimetabolisme menjadi asam piruvat. Ion hydrogen juga dilepaskan sebagai konversi dari piruvat menjadi acetyl CoA dan pada beberapa langkah didalam silkus Krebs. Jika ion hydrogen 17 Universitas Sumatera Utara tetap berada didalam sistem, didalam sel akan menjadi sangat asam. Siklus Krebs merupakan pasangan rangkaian reaksi yang dikenal dengan rantai transport electron. Ion hydrogen dilepaskan selama glikolisis, selama konversi asam piruvat menjadi acetyl CoA, dan selama siklus Kreb’s berkombinasi dengan dua koenzim; nicotinamide adenine dinucleotide (NAD) dan Flavin adenine dinucleotide (FAD), mengkonversi untuk mengurangi bentuk (NADH dan FADH2, sebaliknya). Selama siklus Krebs, tiga molekul NADH dan satu molekul FADH2 dihasilkan. Selanjutnya membawa atom hydrogen (electron) ke rantai transport electron, sebuah kelompok kompleks protein mitokondria terdapat didalam membrane mitokondria. Kompleks protein tersebut terdiri dari rangkaian enzim dan protein yang mengandug zat besi yang disebut cytochromes. Sebagai electron energi tingkat tinggi melewati dari kompleks menuju kompleks disepanjang rantai tersebut, beberapa energi dilepaskan melalui reaksi tersebut yang menggunakan pompa H+ dari matriks mitokondria menuju komparmen luar mitokondria. Setelah ion hydrogen berpindah kembali melewati membrane bagian bawah konsentrasinya menjadi tinggi, energi ditransfer menjadi ADP, dan ATP dibentuk. Langkah akhir membutuhkan enzim yaitu ATP synthase. Gambar 2.6 Rantai Transport Elektron Pada akhir rantai, H+ berkombinasi dengan oksigen untuk membentuk air, selanjutnya mencegah keasaman pada sel. Karena 18 Universitas Sumatera Utara oksigen sebagai aseptor akhir electron dan H+ disebut sebagai oxidative phosphorylation. Untuk setiap pasang electron ditransportasikan ke rantai transport electron oleh NADH, tiga molekul ATP dihasilkan, saat electron melewati rantai transport electron FADH hanya dua molekul ATP dihasilkan. Tetapi, karena NADH dan FADH berada diluar membrane mitokondria, H+ dipompa keluar membrane, membutuhkan energi untuk penggunaannya. Sehingga secara realita, energi yang dihasilkan hanya 2.5 ATP per NADH dan 1.5 ATP per FADH. Oksidasi komplit glukosa dapat menghasilkan 32 molekul ATP, saat 33 ATP dihasilkan dari satu molekul glikogen otot. Produksi bersih ATP dari level substrat posforilasi pada jalur glikolitik hingga siklus Krebs hanya dua ATP (atau tiga dari glikogen). Total 10 molekul NADH hingga ke rantai transport electron – dua pada glikolisis, dua pada konversi asam piruvat menjadi acetyl CoA dan enam pada siklus Kreb’s – hasilnya 25 molekul ATP. Ketika 30 ATP secara actual dihasilkan, energi digunakan untuk transportasi melewati membrane menggunakan lima ATP. Dua molekul FAD pada siklus Krebs dilibatkan pada transport electron menghasilkan tiga tambahan ATP. Pada akhirnya, level substrat posforilasi didalam siklus Krebs melibatkan molekul GTP menambah dua molekul ATP. Jumlah energi yang digunakan untuk memompa electron melewati membrane mitokondria merupakan konsep baru pada pfisiologi olahraga, dna banyak teksbook tetap menyatakan produksi energi yang dihasilkan sekitar 36-39 ATP per molekul glukosa. 2. Oksidasi lemak Lemak juga berkontribusi penting untuk kebutuhan energi otot. Penyimpanan glikogen otot dan hati dapat memberikan sekitar 2.500 kcal energi, tetapi lemak yang disimpan didalam serabut otot dan lemak didalam sel dapat mensuplai sedikitnya 70.000 hingga 75.000 kcal pada dewasa. Walaupun berbagai rangkaian kimia (seperti; trigliserida, 19 Universitas Sumatera Utara posfolipid, dan kolesterol) diklasifikasikan sebagai lemak, hanya trigliserida sebagai sumber utama energi. Trigliserida disimpan didalam sel lemak dan diantara dan didalam serabut otot. Untuk menjadi sumber energi, trigliserida harus dipecah menjadi unit dasar; satu molekul gliserol dan tiga molekul asam lemak bebas. Proses ini disebut lipolysis, dan dibawa oleh enzim yaitu lipase. Asam lemak bebas merupakan sumber energi utama untuk metabolisme lemak. Setelah bebas dari gliserol, asam lemak memasuki darah dan ditransportkan keseluruh tubuh, masuk ke serabut otot bergantung pada konsentrasi gradien. Peningkatan konsentrasi asam lemak bebas didalam darah meningkatkan jumlah ditransportasikan kedalam serabut otot. a) Beta oksidasi (β-Oxidation) Lemak disimpan didalam tubuh pada dua tempat, didalam serabut otot dan didalam sel jaringan adiposa disebut adiposit. Bentuk simpanan lemak adalah trigliserida, dipecah menjadi asam lemak bebas dan gliserol untuk metabolisme energi. Sebelum asam lemak bebas digunakan untuk produksi energi, asam lemak bebas harus dikonversi menjadi acetyl CoA didalam mitokondria, proses ini disebut β-Oxidation. Acetyl CoA dengan segera memasuki siklus Kreb’s untuk metabolisme oksidasi. β-Oxidation adalah rangkaian langkah dua unit karbon acyl dipotong pada rantai karobon asam lemak bebas. Sejumlah langkah bergantung pada jumlah karbon didalam asam lemak bebas, biasanya antara 12 dan 24 karbon. Sebagai contoh, jika sebuah asam lemak bebas mulanya terdiri dari 16 rantai karbon, βOxidation menghasilkan delapan molekul acetyl CoA. Unit acyl menjadi acetyl CoA, sehingga memasuki siklus Kreb’s untuk pembentukan ATP. Setelah memasuki serabut otot, asam lemak bebas diaktivasi secara enzimatik menggunakan energi berasal dari ATP, menyebabkan katabolisme (pemecahan) didalam mitokondria. Seperti glikolisis, β-Oxidation membutuhkan pasokan energi dua 20 Universitas Sumatera Utara ATP untuk aktivasi, tetapi tidak seperti glikolisis, tidak menghasilkan ATP secara langsung. b) Siklus Kreb’s dan Rantai Transport Elektron Setelah β-Oxidation, metabolisme lemak mengikuti jalur yang sama dengan metabolisme karbohidrat oksidatif. Acetyl CoA dibentuk dari β-Oxidation memasuki siklus Kreb’s. siklus Kreb’s menghasilkan hydrogen, selanjutnya ditransportasikan ke rantai transport electron selama electron dihasilkan oleh β-Oxidation untuk mengalami posforilasi oksidatif. Sama seperti metabolisme glukosa, melalui produk oksidasi asam lemak bebas adalah ATP, H2O, CO2. Tetapi, pembakaran sempurna molekul asam lemak bebas membutuhkan oksigen lebih banyak karena molekul asam lemak bebas mengandung lebih banyak karbon dibandingkan molekul glukosa. Gambar 2.7 Produksi energi pada otot skeletal Karbon yang lebih banyak karbon pada asam lemak bebas dibandingkan pada glukosa menyebabkan lebih banyak acetyl CoA yang dibentuk dari metabolisme yang dihasilkan dari asam lemak, sehingga lebih banyak acetyl CoA yang memasuki siklus Kreb’s dan lebih banyak electron yang dikirim ke rantai transport electron 21 Universitas Sumatera Utara sehingga metabolisme lemak dapat menghasilkan lebih banyak energi dibandingkan metabolisme glukosa. Tidak seperti glukosa atau glikogen, asam lemak adalah heterogen dan jumlah ATP yang dihasilkan bergantung kepada oksidasi lemak yang spesifik. Berdasarkan contoh pada asam palmitate, sebuah asam lemak dengan lebih banyak dari 16 karbon. Kombinasi reaksi oksidasi, siklus Kreb’s dan rantai transport electron menghasilkan 129 molekul ATP dari satu molekul asam palmitate, dibandingkan dengan hanya 32 molekul ATP dari satu glukosa dan 33 dari glikogen. (Kenney et al, 2012) 2.1.7. Kontraksi otot skeletal Otot merupakan jaringan dengan kemampuan untuk berkontraksi. Terdapat 3 jenis utama otot; otot skeletal, otot jantung dan otot polos. Pada ketiga jenis otot tersebut, sebagian besar energy yang diproduksi digunakan untuk kebutuhan kontraksi otot yang digunakan oleh molekul aktin pada filament myosin. Gambar 2.8 Kontraksi otot Energi juga digunakan untuk pemompaan Ca 2+ dari sarkoplasma ke reticulum sarkoplasmik setelah otot berkontraksi. Pemompaan ion sodium dan potassium melalui membrane otot untuk mempertahankan gradien ionic yang juga membutuhkan energi. Energi kaya akan komponen adenosine triphosphate (ATP) sebagai bahan bakar di dalam otot. Tetapi suplai ATP otot dapat bertahan sekitar 1-2 detik. Creatine phosphate (CP), yang terdiri dari 22 Universitas Sumatera Utara ikatan posfat energi tingkat tinggi merupakan sumber energy cepat untuk pembentukan ATP. Penyimpanan CP juga terbatas dan mampu memberikan energi untuk kontraksi otot selama 58 detik. Sumber utama energi pada otot berasal dari glukosa dan asam lemak, konsumsi bergantung kepada beban dan latihan subjek dan juga ketersediaan oksigen. Produksi ATP berasal dari glikolisis sitosolik, oksidasi asam lemak beta oksidasi dan siklus asam sitrat yang diatur secara ketat dan berespon cepat terhadap kebutuhan otot untuk memperoleh ATP. Ketika kebutuhan energi melebihi kemampuan otot skeletal menyediakan ATP melalui siklus asam sitrat pada kondisi oksidatif, glikolisis akan dirangsang dan asam laktat diproduksi, menyebabkan ATP secara anaerobik yaitu tanpa menggunakan oksigen. Gambar 2.9 Mekanisme Kontraksi-Relaksasi otot Pada otot skeletal, kontraksi terjadi secara normal pada saat depolarisasi pada plasma membrane, yang didahului oleh pelepasan ion calsium dari penyimpanan di intraselluler didalam reticulum sarkoplasmik. Ion kalsium berikatan dengan troponin C, pengatur protein berhubungan dengan filament tipis, menyebabkan perubahan bentuk protein. Perubahan bentuk tersebut ditransmisikan ke komponen filament tipis lainnya (Troponin T, Troponin I, Tropomyosin dan Actin) sehingga subunit actin pada filament tipis dapat berinteraksi dengan molekul myosin yang terdapat filament tebal. Kontraksi berhenti 23 Universitas Sumatera Utara ketika ion kalsium diambil oleh reticulum sarkoplasmik melalui kerja ATP-driven pump (Ca2+ ATPase). 2.1.8. Pemeriksaan tes muscular endurance (daya tahan otot) Daya tahan otot (muscular endurance) adalah kemampuan sistem neuro-muskular melakukan satu atau dua jenis aksi; (1) kontraksi berulang selama periode waktu hingga terjadinya kelelahan dengan latihan fisik dinamik dan (2) mempertahankan persentase spesifik sebuah kontraksi volunteer maksimal selama periode waktu dengan latihan fisik static. Umumnya daya tahan otot dipresentasikan melalui total jumlah pengulangan yang dapat dilakukan atlit/ non-atlit dengan persentase pada 1RM (One-Repetition Maximum). Metode untuk mengevaluasi daya tahan atlit/ non-atlit termasuk YMCA (Young Men’s Christian Association) bench press test, push-up test dan curl-up test. a. YMCA Bench Press Protocol YMCA bench press test merupakan protocol klasik pengukuran daya tahan otot. Tes ini mengkaji daya tahan otot pada bagian tubuh atas, khususnya otot pektoralis mayor, anterior deltoid dan trisep. Tes ini secara luas digunakan mengkaji daya tahan otot membutuhkan sandaran pada punggung dan mendorong beban secara tetap pada irama 30 repetisi permenit hingga mengalami kelelahan atau tidak mampu mengikuti irama yang diinstruksikan. Kim et al melaporkan YMCA bench press dapat juga digunakan untuk memperkirakan kekuatan maximal bench press dengan prediksi penjumlahan; Pria 1RM (kg) = 1.55 x (repetisi yang dilakukan pada 30 reps.min-1) + 37.9 Perempuan 1RM (kg) = 0.31 x (repetisi yang dilakukan pada 30 reps.min -1) + 19.2 Hanya jumlah maksimum repetisi yang akan ditentukan, kemudian kita membandingkan data dengan hasil normal pada table untuk mengklasifikasikan daya tahan otot bagian atas tubuh (Haff et al, 2012) Tabel 2.1 Nilai Normal daya tahan otot untuk Bench press Usia 18-25 26-35 36-45 46-55 56-65 >65 (Thn) % M F M F M F M F M F M F rank 90 49 49 48 46 41 41 33 33 28 29 22 22 80 34 30 30 29 26 26 21 20 17 17 12 12 70 26 21 22 21 20 17 13 12 10 9 8 6 24 Universitas Sumatera Utara 60 17 13 16 16 12 10 8 6 4 4 3 2 50 5 2 4 2 2 2 1 0 0 0 0 0 Skor didasarkan pada jumlah repetisi komplit dalam 1 menit dengan 80 lb (36.3 kg) pada laki-laki dan 35 lb (15.9 kg) pada perempuan. M = male dan F = female Diambil dari V. Heyward,2010, Advanced fitness assessment and exercise prescription, 6th ed. b. Tes Push-up Tes daya tahan otot pada bagian dada dan lengan atas (pektoralis mayor, anterior deltoid dan trisep), tidak ada batas waktu dan membutuhkan sedikit peralatan. Tes ini dilakukan secara berkelanjutan dan membutuhkan atlit untuk mempertahankan punggung yang sejajar setiap waktu. Tes dihentikan ketika teknik yang tepat tidak dapat dilakukan selama dua kali pengulangan berturutan. Jumlah total repetisi (pengulangan) kemudian digunakan untuk mengtabulasi skor, dibandingkan dengan data sesuai usia dan jenis kelamin pada table normal (Haff et al, 2012). Tabel 2.2 Nilai normal tes Push-up berdasarkan usia dan jenis kelamin Usia (Thn) Klasifikasi 15-19 20-29 30-39 40-49 50-59 60-69 M F M F M F M F M F M F ≥39 ≥33 ≥36 ≥30 ≥30 ≥27 ≥25 ≥24 ≥21 ≥21 ≥18 ≥17 Baik 29-38 25-32 29-35 21-29 22-29 20-26 17-24 15-23 12-20 11-20 11-17 12-16 Cukup 23-28 18-24 22-28 15-20 17-21 13-19 13-16 11-14 10-12 7-10 8-10 5-11 Kurang 18-22 12-17 17-21 10-14 12-16 8-12 10-12 5-10 7-9 2-6 5-7 2-4 Sangat ≤17 ≤11 ≤16 ≤9 ≤11 ≤7 ≤9 ≤4 ≤6 ≤1 ≤4 ≤1 Sangat Baik Kurang Sumber: the Canadian Physical Activity, Fitness & Approach:CSEP-Health & Fitness program’s Health-Related Appraisal and Counselling Strategy, 3rd Edition © 2003. c. Tes Curl-up Tes ini mengkaji daya tahan otot abdominal, umunya berhubungan dengan laboratorium fisiologi olahraga (exercise physiology). Beberapa variasi jarak durasi dari 60 – 120 detik dan membutuhkan sebuah irama pada 20-25 rep.min-1. ACSM merekomendasikan menggunakan waktu 1 menit tes curl-up dengan irama 25 rep.min-1. Repetisi curl-up dihitung hanya jika teknik yang tepat dilakukan, dan jumlah komplit repetisi digunakan untuk skor tes. Hasilnya kemudian dibandingkan dengan table normal berdasarkan usia dan jenis kelamin.. (Haff et al, 2012). 25 Universitas Sumatera Utara Tabel 2.3 Nilai normal tes Curl-up berdasarkan usia dan jenis kelamin Persentil 90 80 70 60 50 40 30 20 10 6 23 20 15 12 10 9 8 7 5 7 27 23 20 16 13 12 10 9 7 8 31 27 25 20 17 15 13 11 9 9 41 33 27 23 20 18 15 14 11 10 38 35 29 27 24 20 19 14 10 Usia (Tahun) 11 12 49 100 40 58 35 48 29 36 26 32 22 31 21 27 18 24 13 18 13 60 55 48 42 39 35 31 30 21 14 77 58 52 48 40 33 30 28 24 15 100 70 60 50 45 40 32 29 22 16 79 61 48 40 37 34 30 28 23 17+ 82 63 50 47 42 39 31 28 24 Sumber: Adapted by permission from Presidents Council for Physical Fitness, Presidents Challenge Normative Data Spreadsheet (Online). Availabel: www.presidentschallenge.org 2.2. MUSCULAR POWER (DAYA LEDAK OTOT) Salah satu komponen yang menunjang dalam pelaksanaan aktivitas olahraga adalah daya ledak (power). Daya ledak adalah kemampuan mengatasi hambatan dalam kecepatan kontraksi otot yang tinggi (Harre,2008). Daya ledak otot (power) merupakan bagian penting pada kemampuan muscular dan diukur dalam satuan watts. Walaupun terdapat hubungan yang erat antara kekuatan (strength) dan kecepatan (velocity) yang menyebabkan pertambahan kekuatan, kemampuan menghasilkan kekuatan pada kecepatan pergerakan yang cepat menjadi karakteristik pada atlit elit pada berbagai cabang olahraga. Kekuatan dihasilkan selama latihan fisik yang sangat bergantung kepada jenis latihan fisik yang digunakan. Pelari jarak jauh menghasilkan hanya 50 W pada setiap siklus langkah selama lari jarak jauh. Secara jelas, atlit angkat beban (atlit angkat besi) menghasilkan 7.000 W pada fase kedua “pull” selama “clean” dan “jerk” (Garhammer,1993; Kraemer dan Loney,2012). Berdasarkan kondisi fisiologi, kekuatan berperan penting dalam membantu tenaga dan kondisi khusus begaimana pertambahan kekuatan. Berdasarkan spesifikasinya, daya ledak dibagi menjadi empat, yaitu; daya ledak eksplosif (explosive power), daya ledak cepat (speed power), daya ledak kuat (strength power) dan daya ledak tahan lama (endurance power) (Nala, 2011). Daya ledak juga merupakan ukuran dari kemampuan otot, yang berkaitan dengan kekuatan dan kecepatan gerak dan dapat didefenisikan sebagai kerja per unit waktu (gaya x jarak/waktu). Gaya dikali kecepatan gerak adalah defenisi yang ekuivalen. Bertambahnya ukuran otot saat berkontraksi dan berkembangnya gaya pada seluruh ROM serta hubungannya dengan kecepatan dan gaya merupakan factor-faktor yang mempengaruhi daya ledak. Gerakan dari daya ledak otot dapat 26 Universitas Sumatera Utara dilihat pada gerakan vertical jump, long jump, angkat besi dan gerakan lain yang melibatkan kontraksi otot. 2.2.1. Komposisi tipe serabut otot Daya ledak otot dihasilkan akibat pemendekan pada sebuah otot hanya sekitar 1% dari panjangnya waktu istirahat, kondisi ini terjadi berkelanjutan setiap waktu pada otot yang berkontraksi. Untuk menhasilkan kekuatan dari area yang aktif, kepala myosin harus berikatan dengan sebuah molekul ATP. Myosin ATPase, sebuah enzim penyusun utama pada kepala myosin, menghindrolisis molekul ATP menjadi ADP dan Pi, selanjutnya kepala myosin akan kembali ke posisi awalnya posisi istirahat. Sumber awal kontraksi pada semua otot adalah adanya ATP yang bebas didalam serabut otot (Brooks et al, 1996; Kraemer et al, 2012). Tetapi penyimpanan ATP pada intramuscular terbatas dan harus diisi ulang melalui satu atau lebih sistem energi tubuh. Daya ledak maksimal bergantung terhadap ketersediaan segera ATP dan sumber energi utama yang diperoleh dari ATP-PCr system yang terdiri dari ATP dan phosphocreatine (PCr) yang tersimpan didalam otot dan reaksi kimia untuk menghasilkan ATP. Walaupun ketersediaan ATP berperan terhadap pencegahan kelelahan, yaitu jumlah myosin ATPase yang memecah ATP membatasi komponen kecepatan kontraksi (Kamen et al, 2000: Kraemer et al, 2012). Hai ini berarti walaupun ketidakadekuatan ATP dapat menghalangi kemampuan, jumlah ATP yang berlebihan tidak menyebabkan manfaat secara fisiologis terhadap keluaran kekuatan. Myosin ATPase yang ada bervariasi jumlahnya atau isoform nya, memiliki karakteristik fungsional yang berbeda. Perbedaan isoform pada myosin ATPase memecah ATP berbeda jumlahnya dan dampaknya pada kecepatan kontraksi dan daya ledak yang dihasilkan. Jumlah myosin ATPase pada tipe serabut otot yang memecah ATP secara terbalik proporsionalnya dengan kemampuan oksidatifnya (Kraemer et al, 2012). Hal ini berarti walaupun serabut otot tipe II menyebabkan output daya ledak lebih besar potensialnnya, lebih mudah mengalami kelelahan dan kemampuannya lebih rendah menetralkan kondisi asam disekitar intraselluler. Tetapi, serabut otot tipe II menyebabkan penyimpanan ATP dan PCr lebih besar dan meningkatkan aktivitas enzim glikolitik, segingga menyebabkan kemampuan lebih baik pada latihan fisik dengan intensitas tinggi dalam waktu yang singkat. Persentase tipe serabut membentuk unit fungsional pada setiap unit motor bervariasi melalui peran sebuah otot pada pergerakan manusia. Sebagai contoh, sebuah otot pada area abdominal terlibat secara utama mendukung postur yang dibentuk secara utama oleh serabut tipe I, berperan utama pada pergerakan otot atau pada otot lokomotif, seperti vastus lateralis yang salah satu tipe serabut otot khusus sekitar 40-60% tipe II ke tipe II (Staron et al, 2000; 27 Universitas Sumatera Utara Kraemer et al, 2012). Pada atlit elit lari marathon memiliki 80-90% serabut otot tipe I, hal ini akan menyebabkan kemampuan berlari sub-2:10.00 maraton. Pad kebutuhan diatas 80% di unit motornya, persentasi tinggi pada serabut otot tipe II akan metabolik dan secara mekanik tidak akan efisien sehingga menyebabkan lebih cepat merasa kelelahan, misalnya pada lari jarak jauh. Sebaliknya, tingkat kekuatan yang tinggi tidak dapat dihasilkan, kecuali seorang atlit serabut otot tipe II pada unit motor persentasenya lebih tinggi, misalnya persentase ekstrim tipe II serabut otot (>70%). Walaupun persentase serabut otot pada laki-laki dan perempuan mirip, area crosssectional pada serabut otot pada laki-laki secara khusus lebih besar dibandingkan perempuan. Serabut otot tipe II paling banyak pada laki-laki dan lebih dominan setelah melakukan program latihan. Pada perempuan yang tidak terlatih, menariknya, serabut otot tipe I lebih banyak dibandingkan serabut otot tipe II tetapi setelah latihan yang dikaitkan dengan hasil investigasi perubahan yang sama pada laki-laki. 2.2.2. ATP-PCr System (Sistem ATP-PCr) Kontraksi otot yang sangat kuat terjadi dalam waktu yang singkat, jumlah ATP yang dihasilkan tinggi, isi ulang harus dengan segera. Walaupun ATP dapat diperoleh dari pemecahan glukosa secara anaerobik (anaerobic glycolysis) dan jalur aerobik (Kreb’s Cycle), hal ini tidak dapat mengatasi tingginya kebutuhan aksi daya ledak pada latihan fisik. Akibatnya, ATP disediakan oleh penyimpanan PCr didalam serabut otot itu sendiri melalui sistem ATPPCr. Pada sistem ATP-PCr, ADP dihasilkan selama kontraksi otot yang dapat diubah kembali menjadi ATP melalui enzim creatine kinase karena menyebabkan reaksi didalam PCr intramuscular. Proses ini berperan untuk menghasilkan ATP yang akan digunakan selama latihan fisik sedikitnya 2 sampai 10 detik. Walaupun efektif dalam waktu yang terbatas, penyimpanan PCr sangat cepat habis selama 10 detik pertama pada latihan fisik maksimal. Pada otot tertentu, PCr yang disimpan secara total dapat habis dalam waktu kira-kira 4 detik. Secara umum, PCr yang disimpan didalam otot adalah 50% diisi ulang dari total yang habis setelah 1 menit istirahat dan penuh selama 5-6 detik saat istirahat. Pada kondisi tersebut penambahan creatine terlibat pada daya ledak dan enduransi daya ledak meningkat melalui pengumpulan creatine pada intramuscular. (Kraemer et al,2012). 28 Universitas Sumatera Utara 2.2.3. Pemeriksaan daya ledak otot (muscular power) a. Standing Broad Jump Merupakan jenis pemeriksaan daya ledak otot pada tungkai yang mudah dinstruksikan dan dilakukan. Standing broad jump sering digunakan selain vertical jump untuk mengukur daya ledak otot tungkai. Subjek melakukan lompatan kedepan sejauh mungkin dengan kedua kaki melompat dan mendarat secara bersamaan. Jauh lompatan diukur menggunakan pita pengukur dari garis start sampai ke titik pendaratan tumit. Sama dengan vertical jump, kemampuan lompatan paling jauh dicatat dan bergantung pada protocol. Nilai jauh lompatan terbaik dua atau tiga kali dicatat. Hubungan signifikan antara standing broad jump dengan pemeriksaan daya ledak (r = 0.61) juga dilaporkan sebagai hasil penelitian. Penelitian yang dilakukan oleh Larson menyatakan standing broad jump dan vertical jump memiliki korelasi sangat tinggi (r = 0.79) dengan pengukuran daya ledak otot. Pengukuran standing broad jump dapat dilakukan pada anak perempuan dan anak laki-laki minimal berusia 6 tahun. (Amstrong dan Mechelen, 2008) Tabel 2.3 Nilai Normal Standing Broad Jump test berdasarkan usia Persentil 99 95 90 80 75 70 60 50 40 30 25 20 10 5 1 Mean 7 169 157 149 140 137 134 128 123 118 112 109 105 96 88 74 123 8 176 162 155 145 142 139 134 128 123 118 115 111 102 85 81 128 9 184 170 162 153 149 146 141 135 130 125 121 118 109 102 88 139 Anak laki-laki Usia (tahun) 10 11 12 13 14 15 195 208 221 235 248 258 180 191 204 217 230 240 171 183 195 208 220 231 162 173 185 197 209 220 158 169 181 193 205 215 155 165 177 189 201 211 149 159 171 182 194 204 144 154 165 176 187 197 139 148 159 170 181 191 133 142 153 163 174 183 130 139 149 159 170 179 126 135 145 155 165 175 117 126 135 144 154 163 110 118 127 135 144 153 95 103 119 118 126 134 144 154 165 176 187 197 16 266 249 239 228 224 220 213 206 200 192 188 184 171 161 142 206 17 272 255 246 235 231 227 220 213 207 200 196 191 179 170 150 213 18 277 260 250 240 236 232 225 219 213 206 202 198 187 178 158 219 b. Vertical Jump Kemampuan Vertical power atau jumping merupakan salah satu tes popular yang digunakan untuk mengkaji daya ledak anaerobik pada populasi atlit. Kemampuan untuk 29 Universitas Sumatera Utara mengekspresikan tingginya output vertical power atau tinggi lompatan berhubungan dengan berbagai aktivitas olahraga, termasuk kompetisi angkat beban (angkat besi), balap sepeda, kemampuan berlari. Kemampuan melompat juga menunjukkan perbedaan diantara tingkat permainan dan kemampuan bermain pada sepakbola Amerika dan sepakbola Terdapat berbagai macam tes daya ledak,salah satunya adalah vertical jump test.. Vertical jump test dikenal juga dengan nama sarget test. Tes ini dikembangkan oleh Dr.Dudley Allen Sargent yang bertujuan untuk mengukur daya ledak (power) otot-otot tungkai dengan menggunakan dinding berskala centimetre (Quinn, 2013). Vertical jump test didukung oleh peran utama dari otot penggerak tubuh, yaitu kelompok otot quadriceps femoris. Karena itu peningkatan vertical jump harus bertahap dan diperlukan adaptasi dari otot quadriceps femoris sebagai penggerak utama. Dalam meningkatkan kekuatan otot apabila serabut otot banyak, maka kekuatan otot akan besar sehingga kekuatan otot yang besar akan mendukung terciptanya vertical jump test yang baik. Loncat tegak (vertical jump) adalah suatu gerakan mengangkat tubuh dari suatu titik ke titik yang lebih jauh atau lebih tinggi dengan ancang-ancang lari cepat atau lambut dengan menumpu dua kaki dan mendarat dengan kaki atau anggota tubuh lainnya dengan keseimbangan yang baik. Loncat adalah suatu menolak tubuh atau melompat ke atas dalam upaya membawa titik berat badan selama mungkin di udara (melayang di uadara) yang dilakukan dengan cepat dengan jalan melakukan tolakan pada dua kaki untuk menolak tubuh setinggi mungkin. Loncat adalah lompat dengan kedua atau keempat kaki secara bersamasama. Pada pengukuran yang pertama dan kedua menggunakan vertical jump test. Tes ini dilakukan sebanyak tiga kali, dan hasil atau pencapaian terbaik akan dimasukkan kedalam data. Tabel 2.5 Nilai normal Vertical Jump test berdasarkan usia dan jenis kelamin Usia/ klasifikasi Sangat baik Baik Cukup Kurang Sangat Kurang 15-19 tahun M F ≥56 cm ≥40 cm 51-55 cm 36-39 cm 46-50 cm 32-35 cm 42-45 cm 28-31 cm ≤41 cm ≤27 cm Sumber: the Canadian Physical Activity, Fitness & Approach:CSEP-Health & Fitness program’s Health-Related Appraisal and Counselling Strategy, 3rd Edition © 2003. Selanjutnya kita dapat memprediksi atau menghitung jumlah peak anaerobic power output dari sistem energi dalam satuan watt (PAPw) dari tinggi vertical jump dalam satuan 30 Universitas Sumatera Utara sentimeter (VJcm) dan berat badan dalam satuan kilogram (BWkg). Penghitungan dapat digunakan pada laki-laki maupun perempuan: PAPw = (60.7 x VJcm) + (45.3 x BWkg) – 2055 Contoh: seorang laki-laki berusia 21 tahun dengan berat badan 78 kg, catatan vertical jump 43 cm ( tinggi jangkauan saat berdiri = 85 cm; tinggi vertical jump = 228), maka prediksi peak anaerobic dalam satuan watt : PAPw = (60.7 x VJcm) + (45.3 x BWkg) – 2055 = (60.7 x 43 cm) + (45.3 x 78 kg) = 4088.5 W Selanjutnya hasil perhitungan bandingkan dengan tabel normal peak anaerobic power berdasarkan usia dan jenis kelamin. Tabel 2.6 Nilai normal peak anaerobic power output berdasarkan usia dan jenis kelamin Usia/ klasifikasi Sangat baik Baik Cukup Kurang Sangat Kurang 15-19 tahun M ≥4,644 W 4,185-4,643 W 3,858-4,643 W 3,323-3,857 W ≤3,322 W F ≥3,167 W 2,795-3,166 W 2,399-2,794 W 2,156-2,398 W ≤2,155 W Sumber: the Canadian Physical Activity, Fitness & Approach:CSEP-Health & Fitness program’s Health-Related Appraisal and Counselling Strategy, 3rd Edition © 2003. 2.3. GEN PEROXISOME PROLIFERATOR-ACTIVATOR RECEPTOR ALPHA (PPARα) Metabolisme nutrient dan homeostasis energi diatur oleh endokrin, parakrin dan signal autokrin yang mengontrol ekspresi dan aktivitas enzim metabolik dan mentransport protein melalui mekanisme transkripsional dan posttranskripsional. Mediator lipid mempunyai peran penting pada pengontrolan metabolik. PPARs (NR1Cs/ Nuclear Receptor subfamily 1, group C, members) merupakan sebuah kelas ligand-activated transcription factor, berperan sebagai pengatur transkripsional metabolisme lipid dan karbohidrat. Lipid jenuh dan lipid tidak jenuh dan derivative eicosanoid nya merupakan aktivator alami pada subkelas nuclear receptors tersebut. Peningkatan peran PPARs pada pengaturan metabolik disebabkan oleh adanya fibrate hipolipidemia dan thiazolidinediones-sensitizer insulin yang mengsintesis ligand PPARα 31 Universitas Sumatera Utara (NR1C1) dan PPARγ (NR1C3), PPARδ atau yang lebih dikenal PPARβ yang merupakan isotipe PPAR (Lefebvre et al, 2006). PRARs merupakan nuclear reseptor pengaturan asam lemak membentuk pasangan heterodimer dengan RXRs (retinoid X recerptors) dan berikatan dengan peroxisome prolifetaror regulatory elements (PPREs) pada promotor yang memberi respon pada gen. semua sub tipe PPAR (⍺, β/δ,γ1 dan γ2) mengikat asam lemak (Xu et al, 1999; Jump; et al, 2006); ikatan asam lemak menstimulasi rekrutmen coactivator dan transkripsi gen. PPARs merupakan sensor lipid intraseluler, level intraseluler non-esterified fatty acid (NEFA) pada hepatosit tikus sebelum dan sesudah perlakuan. Hasil penelitian menyatakan bagaimana asam lemak mengontol fungsi PPAR⍺. Pertama, level NEFA intraseluler dipertahankan pada level sangat rendah dengan hasil <0.1% dari total lipid didalam sel. Kedua, perubahan pada sel terkait asam lemak, menyebabkan perubahan signifikan pada NEFA intraseluler dan menunjukkan respon yang kuat pada target gen PPAR⍺, seperti cytochrome P450-4A (CYP4A) dan cytosolic thioesterase (CTE-1). Ketiga, dampak asam lemak pada signalling PPAR⍺ adalah cepat, tetapi bersifat sementara. Pada dasarnya asam lemak merubah target gen PPAR⍺ secara parallel merubah NEFA intraseluler. Seperti pada konsep penelitian bahwa PPARs memonitor komposisi NEFA intraseluler dan berespon secara sesuai dengan jalur metabolic yang dirangsang untuk meminimalkan kerusakan akibat NEFA intraseluler yang berlebihan (Pawar et al, 2003; Jump et al, 2006). PPARα sebagian besar diekspresikan di hati, tetapi juga ditemukan pada myocite jantung, proximal tubular sel epitel ginjal, otot skeletal, epitel usus besar, endothelium dan sel otot polos dan juga pada sel kekebalan termasuk makrofag, limfosit dan granulosit (Kliewer et al.,1994; Varga et al., 2011). PPARα merupakan kunci pengaturan peroksisome dan β-oksidasi mitokondria asam lemak, sintesis badan-badan keton dan metabolisme sistemik lipid. Oksidasi lipid secara utama terjadi didalam hati dan untuk mencegah steatopsis (penumpukan lemah di hati). Selama pasokan asam lemak, merangsang transkripsi PPARα untuk mengatur gen dan sistem oksidasi (sistem oksidasi microsomal omega dan beta oksidasi mitokondrial dan beta oksidasi peroksisomal) diaktivasi. Aktivasi dan peningkatan sensitivitas didalam hati menyebabkan peningkatan pembakaran energi dan menurunkan penyimpanan lemak. Fungsi PPARα sebagai sonsoring lipd dan mengontrol pembarakan energi. Juga memiliki peran dalam pathogenesis fatty liver disease (FLD) dan ligan pada reseptor mungkin efektif didalam penurunan staetosis melalui meningkatkan penggunaan energi (Grygiel dan Górniak, 2014). 32 Universitas Sumatera Utara PPARα dapat diaktivasi oleh ligand secara natural ataupun sintetik, seperti Polyunsaturated Fatty Acid, eicosanoid dan obat-obatan hipolipidemik (fibrates) dan mengatur transkripsi DNA dengan cara berikatan dengan sekuensi nukleotida yang spesifik berlokasi pada region pengaturan pada target gen, seperti; peroxisome proliferator responsive elements (PPRES). Ikatan Ligand dengan DNA membutuhkan heterodimer, yaitu PPARα dan Retinoic X receptor ⍺ yang berinteraksi dengan PPRE yaitu konsensus sekuensi (AGGTCA N AGGTCA) terdiri dari 2 pengulangan dipisahkan oleh 1 nukleotida. Ketika ligand berikatan dengan PPARα, terjadi perubahan bentuk pada PPARα-retinoic X receptors sehingga mengaktivasi kompleks transkripsional untuk berpasangan dengan protein coactivator, salah satunya secara sekuensi atau menjadi pasangan modul subkomplek. Selanjutnya kompleks coactivator acetylate (steroid receptors coactivators,p300) meremodeling kromatin nukleosom dan komponen mediator PPARα untuk memfasilitasi perekrutan pada bagian transkripsi basal dengan RNA polymerase II untuk transkripsi pada target gen spesifik (Contreas, et al.,2012). Gambar 2.10 Peran PPARα Hepatik pada PUFA 2.3.1. Struktur dan fungsi PPARα Struktur 3 dimensi PPARs terdiri dari DNA binding domain pada N-terminus dan sebuah ligand binding domain (LBD) pada C-terminus. Setelah interaksi dengan agonis, PPARs ditranslokasikan ke nucleus dan heterodimerisasi dengan nuclear receptor lain, yaitu retinoid X receptor (RXR). RXR membentuk sebuah heterodimer dengan sejumlah reseptor lain (misalnya; vitamin D atau hormone tiroid). Area spesifik DNA sebagai target area yang telah berikatan dengan PPARs disebut Peroxisome proliferator hormone response elements (PPREs). PPREs ditemukan pada promotor gen respon PPAR, seperti fatty acid-binding protein (aP2). Pada sebagian besar kasus, proses ini mengaktivasi transkripsi pada berbagai gen melibatkan berbagai proses fisiologi dan patofisiologi. 33 Universitas Sumatera Utara Fungsi PPARs dimodifikasi oleh sejumlah coactivator dan core processor, dengan cara menstimulasi atau menghambat fungsi reseptor, atau sebaliknya. Ligand mengaktivasi PPARγ-RXR, sehingga menyebabkan pertukaran co-repressor menjadi co-activator. Pada sel manusia terdapat berbagai cofactor bergantung kepada jenis sel dan hubungan cofactor secara khusus dengan gen lainnya. Peroxisome proliferator-activated receptor dibagi menjadi 3 isoform; PPARα, PPARβ/δ dan PPARγ. Setiap isoform berbeda pendistribusiannya, spesifikasi litgand dan peran fisiologisnya. Setiap isoform berfungsi untuk mengaktivasi atau menghambat gen yang berbeda. Semua isoform berperan pada homeostasis lipid dan pengaturan glukosa (keseimbangan energi) dan kerjanya secara terbatas pada jenis jaringan tertentu. Gambar 2.11 target gen PPARα dan mekanisme transkripsi PPARα secara mayoritas diekspresikan pada jaringan dengan metabolik aktif, seperti hati, jantung, otot skeletal, mukosa intestinal dan jaringan adiposa cokelat. Reseptor ini berimplikasi pada metabolisme lipid dan aktif jika kadar lipid rendah. 34 Universitas Sumatera Utara 2.3.2. Aktivasi transkripsional oleh PPARα Pengaturan transkripsional pada gen metabolik pada otot dimulai dengan signal ekstriksik dari jaringan, seperti; peptida, hormone steroid dan katekolamin dan juga signal intriksik dari jaringan, seperti; peregangan mekanik, kalsium intraselluler dan ketersediaan nutrien. Myocyte enhancer factor 2 (MEFC2) merupakan mediator awal berbagai signal dan berperan penting pada metabolisme otot melalui perangsangan aktivitas transkripsi pada dua pengatur metabolik; peroxisome proliferator-activated receptor alpha (PPARα) dan peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC1α) (Czubryt et al, 2003; Baskin et al, 2015). Selama latihan fisik, berbagai jalur signalling diaktifkan sehingga menyebabkan perubahan transkripsional pada otot. Sebagai contoh, peningkatan kalsium intraselluler mengaktivasi calcium/calmodulin-dependent protein kinase II (CaMKII), peregangan mekanik mengaktivasi mitogen-activated protein (MAP) dan c-Jun N-terminal kinase dan perubahan rasio NAD/NADH dan AMP/ATP mengaktivasi sirtuin dan AMP-activated protein kinase (AMPK) (Egan dan Zierath, 2013; Baskin et al, 2015). Sehingga aktivitas transkripsional PPARα dan PGC1α meningkat, menstimulasi biogenesis mitokondrial dan oksidasi asam lemak meningkat. Pengaturan transkripsional pada gen metabolic merupakan hal penting terhadap otot lurik terkait dengan perubahan fisiologis dan secara utama diatur oleh Nuclear Receptors (NRs). Proses transaktivasi Nuclear Receptors (NRs) melalui 5 langkah utama; ikatan dengan ligand; ikatan yang stabil ligand NRs dengan DNA; pelepasan coreprosesor dan pengikatan coactivator; aktivasi transkripsi; dan disosiasi transkripsional kompleks; disertai dengan salah satunya shut-down atau reinisiasi transkripsi. Penelitian secara kristalografi menyatakan bahwa ikatan ligand dengan PPARα menyebabkan stabilisasi global pada bentuk reseptor (Xu et al, 2002), tanpa pengaturan kembali struktural, tidak seperti reseptor LBD prototipe Retinoic acid mengalami transisi struktural utama pada ikatan. Gambar 2.12 Mekanisme transkripsi gen oleh PPARs 35 Universitas Sumatera Utara Hal ini menyebabkan kestabilan ligand PPARα dikenal sebagai struktur yang dapat digunakan protein-protein berinteraksi dengan coaktivator (agonist-bound PPARα) atau coreprosessor (antagonist-bound PPARα) (Xu et al, 2002). Situasi ini umum terjadi pada PPARα, PPARγ dan PPARδ. Selanjutnya kompleks coactivator dan coreprossessor berfungsi sebagai enzimatik (misalnya; acetylase, deacetylase, methylase, demethylase dan kinase)dengan target kromatin, komponen pada dasar tempat transkripsional dan coactivator dan coreprosessor lainnya. Pelepasan coaktivator dan coreprosessor menyebabkan pembukaan kromatin dan preinisiasi kompleks pasangan pada promotor. DNA-binding domain (DBD) menyebabkan PPARα dapat berikatan dengan PPAR response element (PPREs) pada promotor target gen sebagai heterodimer dengan isotipe (Wan et al, 2003; Lefebvre et al, 2006 Retinoid E receptors (RXR). PPERs secara khusus di atur langsung secara berulang pada sekuensi inti nya AGGTCA yang dipisahkan oleh 1 atau 2 nukleotida (DR1 dan DR2), diapit kebagian awal oleh sekuensi A/T yang banyak. Ketika geometri PPERs dipastikan untuk heterodimer PPAR/RXR, DR1 dan DR2, PPERs juga dikenali oleh heterodimers PPERs, sehingga mendukung cross-talk dengan RARs dan RXrs yang dapat mempengaruhi pengontrolan metabolik. C-terminus pada PPARα, dimana struktur 3D telah di uraikan, yang terdiri dari ligand-regulated E domain atau AF-2 atau ligand-binding domain (LBD), sebagai tempat utama T-shaped ligand-binding pocket untuk sintesis ligand (Cronet et al, 2001; Ijpenberg et al, 2004; Lefebvre et al, 2006). Gambar 2.13 Mekanisme molekular aksi PPARα 36 Universitas Sumatera Utara 2.3.3. Polimorfisme gen PPARα PPAR⍺ berlokasi pada kromosom 22q13.3 dan terdiri dari delapan ekson. Single nucleotide polymorphisms (SNPs) pada gen tersebut menunjukkan hubungan dengan dislipidemia, resistensi insulin, diabetesdan cardiovascular disease (Yong et al,2008; Chen et al, 2010). L162V polymorphisms (rs1800206) menunjukkan substitusi “C” menjadi “G” menyebabkan asam amino leucine menjadi valine, pertukaran pada kodon 162 (Flavel et al, 2000; Sapone et al, 2000; Chen et al, 2010). Polimorfisme gen ini juga berhubungan dengan variasi pada konsentrasi plasma total kolesterol, LDL, HDL, TG, APOB dan APOA1, tetapi hubungannya masih kontroversi (Yong et al,2008; Chen et al, 2010). Polimorfisme intron 7 G>C polymorphisms menyebabkan substitusi “G” menjadi “C” pada intron 7 (rs4253778). Hubungan antara SNP ini dengan rendahnya nilai TG diobservasi pada pasien diabetik. Sebaliknya, alel “C” pada intron 7 G>C berhubungan dengan progress aterosklerosis (Flavel et al, 2002; Chen et al, 2010) dan perkembangan ventrikel kiri akibat olahraga (exercise) (Jamshidi et al, 2002; Chen et al, 2010). Gen PPAR⍺ sangat penting dalam metabolisme glukosa dan lipid (Roth et al, 2007; Baar et al, 2004; Liang et al, 2006; Russel et al, 2003; Ginevičinė et al, 2010) dan sering diekspresikan didalam serabut otot tipe I (slow-twich) dibandingkan dengan serabut otot tipe II (fast-twich) (Ahmetov II et al, 2009; Russel et al, 2003; Ahmetov II et al, 2006; Ginevičinė et al, 2010). PPAR⍺ alel G diasumsikan sebagai alel enduransi dan alel C sebagai alel kecepatan (speed)/ tenaga (power) (Bray et al, 2009; Ahmetov II et al, 2009; Ahmetov II et al, 2006; Ginevičinė et al, 2010). Hasil penelitian menyatakan bahwa PPAR⍺ alel C lebih banyak ditemukan kelompok atlit dibandingkan pada populasi umum Lituania. PPAR⍺ genotip GG lebih banyak ditemukan pada atlitenduransi dan tim olahraga dibandingkan pada kecepatan/ tenaga dan olahraga campuran. Pria dengan kategori olahraga yang mengutamakan kecepatan/ tenaga memiliki nilai single muscular contraction power (SMCP) dan anaerobic alactic muscular power (AAMP) yang tinggi sehingga mendukung alel C menentukan kecepatan dan tenaga (Ginevičinė et al, 2010). Beberapa hasil penelitian juga menyatakan dampak PPAR⍺ pada kemampuan atletik. Ahmetov et al menyatakan bahwa alel G secara signifikan lebih tinggi pada atlit Rusia yang berorientasi pada enduransi (80.3%) dibandingkan dengan atlit yang berorientasi pada power (50.6%) karena alel C memberikan manfaat terhadap aktivitas tipe power tenaga (power). Ahmetov juga menyatakan homozigot GG secara signifikan persentasenya lebih tinggi pada serabut otot tipe I (55.5%±2.0% versus 38.5%±2.3%) dibandingkan homozigot CC. 37 Universitas Sumatera Utara polimorfisme PPAR⍺ intron 7 G/C berhubungan dengan kemampuan fisik atlit Rusia dan hubungan antara genotip PPAR⍺ dengan komposisi serabut otot. Hasil penelitian lainnya menyatakan secara signifikan ditemukan alel G lebih dominan pada pemain sepakbola Italia (80% dibandingkan dengan 70% pada subjek control) (Proia et al, 2014). Polimorfisme PPAR⍺ alel G berhubungan dengan peningkatan oksidasi asam lemak pada otot skeletal dan peningkatan proporsi serabut otot tipe I (slow twich) (Ahmetov II et al, 2006), karena serabut otot tersebut lebih efisien menggunakan oksigen selama berlangsung kontraksi (Lopez-Leon et al, 2016). Hasil observasi penelitian menyatakan alel C pada polimorfisme PPAR⍺ intron 7 G/C berhubungan dengan penurunan transkripsi mRNA PPAR⍺ sehingga kadar PPAR⍺ lebih rendah yang berdampak pada aktivasi transkripsional PPAR⍺ pada target gen dan menyebabkan penurunan oksidasi asam lemak. Mekanisme proses ini masih kurang jelas diketahui. Beberapa peneliti mengasumsikan bahwa alel C intron 7 mungkin dihubungkan dengan ketidakseimbangan varian yang belum diketahui pada area pengaturan pada gen PPAR⍺ yang merubah kadar protein PPAR⍺ dan dampaknya adalah perubahan pada proses transkripsional pada target gen PPAR⍺ (Ahmetov II et al, 2006; Maciejewska-Karlowska, 2013), yaitu penurunan oksidasi asam lemak dan tingkat metabolisme oksidasi pada otot skeletal. Hasil temuan Ahmetov II et al mengasumsikan variasi pada intron 7 gen PPAR⍺ dihubungkan dengan fenotip performa. Hasil temuan, yaitu; homozigot GG lebih dominan ditemukan pada kelompok atlit yang berorientasi pada enduransi dan hasil observasi alel C lebih dominan ditemukan pada kelompok kemampuan fisik aerobic. Hasil penelitian tersebut mungkin menjelaskan hubungan antara genotip PPAR⍺ dengan komposisi serabut otot. Atlit yang berorientasi pada tenaga (power) dikarakteristikkan dengan frekuensi alel C yang lebih besar, yaitu otot skeletal cenderung hipertrofi dan merubah substrat energy menyebabkan penurunan oksidasi asam lemak sebagai respon terhadap latihan fisik anaerobic. Secara jelas, frekuensi genotip pada atlit enduransi mungkin dihubungkan dengan peningkatan oksidasi asam lemak didalam otot skeletal dna peningkatan proporsi serabut otot tipe I (slow-twich) pada individu dengan genotip GG. Hasil tim penelitian pada polimorfisme intron 7 G/C gen PPAR⍺ pada atlit Polandia menyatakan distribusi genotip GG dan alel G lebih tinggi pada kelompok atlit dayung dibandingkan dengan kelompok kontrol tidak terlatih. Hal ini didasarkan pada penelitian sebelumnya terkait intron 7 alel G berhubungan dengan kemampuan enduransi, sehingga diasumsikan alel G berhubungan dengan ekspresi normal gen PPAR⍺ dan mempertahankan 38 Universitas Sumatera Utara level normal protein PPAR⍺ sebagai komponen penting terhadap respon adaptasi terhadap latihan enduransi. Kemungkinan pada subjek sehat, ketika plasma lipid dalam rentang normal, aktivitas PPAR⍺ tidak sebagai factor penghambat dan selanjutnya (seperti, variasi pada intron 7 G/C) yang diasumsikan menyebabkan perubahan fungsi yang sedikit, tetapi tidak menunjukkan pengaruh. Tetapi pada atlit enduransi yang melakukan latihan jangka lama, plasma lipid sering menjadi melebihi normal yaitu merangsang cadangan lemak yang akan mengaktivasi PPAR⍺, meningkatkan fungsi yang berbeda yang menjelaskan relevansi alel G pada intron 7 pada atlit enduransi (Maciejewska-Karlowska, 2013). 2.4. Asas Hardy-Weinberg Equilibrium Asas Hardy-Weinberg menyatakan bahwa frekuensi alel dan frekuensi genotipe dalam suatu populasi akan tetap konstan, yakni berada dalam kesetimbangan dari satu generasi ke generasi lainnya kecuali apabila terdapat pengaruh-pengaruh tertentu yang mengganggu kesetimbangan tersebut. Pengaruh-pengaruh tersebut meliputi perkawinan tak acak, mutasi, seleksi, ukuran populasi terbatas, hanyutan genetik, dan aliran gen. Kesetimbangan genetik adalah suatu keadaan ideal yang dapat dijadikan sebagai garis dasar untuk mengukur perubahan genetik. Frekuensi alel yang statis dalam suatu populasi dari generasi ke generasi mengasumsikan adanya perkawinan acak, tidak adanya mutasi, tidak adanya migrasi ataupun emigrasi, populasi yang besarnya tak terhingga, dan ketiadaan tekanan seleksi terhadap sifatsifat tertentu. 39 Universitas Sumatera Utara