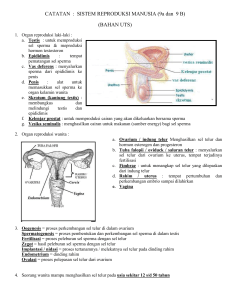
Variasi kesuburan sapi jantan dan strategi untuk meminimalkan
advertisement

Variasi kesuburan sapi jantan dan strategi untuk meminimalkan konsekuensi dari infertilitas dan / atau subfertilitas sapi jantan di tingkat kawanan. Flowers [40] baru-baru ini membandingkan perkiraan fertilitas dari kawanan dan individu sapi jantan dan menyimpulkan bahwa sapi jantan memiliki efek lebih besar pada kesuburan kawanan daripada sapi betina, mungkin karena meluasnya penggunaan alami dan kurangnya evaluasi yang tepat dari sapi jantan sebelum digunakan sebagai indukan kawanan. Kesuburan biasanya meningkat dengan dosis sperma, sampai ambang batas tercapai [41], berdasarkan konsep dari ciri sperma yang bersifat kompensantorik dan non-kompensantorik [36,41-45]. Ciri dari fertilitas jantan yang dapat dikompensasikan adalah dengan meningkatkan dosis inseminasi [36,40], berdasarkan asumsi bahwa tingkat ambang sperma yang kompeten (dengan tingkat motilitas yang dapat diterima, morfologi normal, dan kemampuan untuk menjalani kapasitasi dan memulai sebuah akrosom reaksi, misalnya, 70% sperma morfologi normal) adalah diperlukan untuk mencapai pembuahan normal. Oleh karena itu, bahkan jika inseminasi tersebut mengandung sperma yang rusak dengan kecacatan yang bersifat kompensatorik, kekurangan tersebut bisa diatasi dengan peningkatan jumlah sperma yang kompeten. Namun, infertilitas karena sifat non-kompensatorik tidak dapat diatasi dengan peningkatan jumlah sperma, karena sperma tersebut biasanya tidak dapt menyelesaikan pembuahan dan tidak dapat mempertahankan embrio [40]. Sebagai contoh, jika inseminasi tersebut mengandung proporsi sperma dengan DNA yang abnormal yang lebih tinggi dari, sperma mungkin bisa mencapai tempat fertilisasi dan memulai melakukan blok terhadap polispermia. Menyuntikkan dengan lebih banyak sperma juga tidak akan bermanfaat, karena proporsi sperma yang abnormal juga meningkat, dan peluang untuk sperma yang abnormal untuk berinteraksi dengan oosit dan memulai blok polispermia tetap sama. Industri AI komersial mengerahkan tindakan pengendalian mutu yang ketat dengan mengevaluasi ciri kompensatorik dan non-kompensatorik tertentu dan membuang sampel suboptimal [36]. Pendekatan diagnostik yang sudah maju dijelaskan dalam bagian berikut diharapkan dapat meningkatkan kemampuan kita untuk mengevaluasi sifat kompensatorik dan non-kompensatorik pada semen yang terlihat normal yang dapat mempengaruhi fertilitas pada berbagai tahap fertilisasi dan perkembangan embrio awal 8. Teknik yang lebih maju dalam memprediksi fertilitas 8.1. Analisis sperma dibantu komputer Sistem ini memungkinkan evaluasi obyektif dari berbagai karakteristik kinematic sperma. Sewaktu sperma bergerak maju di dalam trajektori motlitas, velositas, liniaritas dan penempatan lateral dari kepala sperma dapat ditentukan. Pentingnya motilitas total dan progresif dan hubungannya dengan fertilitas di lapangan dapat ditentukan [46,47]. Membandingkan parameter kinematik sperma sapi jantan dari kelompok dengan fertilitas rendah dan fertilitas tinggi, kami mendalilkan bahwa sperma beku yang dicairkan dari sap jantan dengan fertilitas tinggi lebih efisien dalam melalui hiperaktivasi dibandingkan dengan sapi jantan dengan fertilitas rendah dan mampu melintasi hambatan dalam saluran reproduksi betina dan mencapai tempat fertilisasi [48]. Selain itu, analisis sperma yang dibantu komputer dapat mengidentifikasi subpopulasi sperma berdasarkan karekteristk dari pergerakan sperma [49], yang mungkin lebih baik dalam memprediksi fertilitas. 8.2. DNA sperma Kualitas DNA sperma merupakan penentu utama dalam keberhasilan fertilitasi dan perkembangan yang kompeten. DNA sperma matang sangat kental (karena protamines). Penyimpangan dalam DNA (misalnya, single atau double-stranded break atau cacat protein yang menggangu konversi histone atau protamine atau mengubah pemadatan DNA dan konfigurasikromatin) bisa merusak dikondensasi nuklir dan mempengaruhi pengembangan [50]. DNA induk yang rusak bisa terinkoporasi ke dalam genom embrio (efek trans-generasi [51]). DNA sperma dinilai dengan sifat metakromatik akridin oranye. Dalam beberapa spesies (termasuk sapi), struktur kromatin sperma assay sangat berhubungan dengan fertilitas [52,53]. 8.3. RNA sperma DNA sperma dianggap tidak aktif, sperma matang melakukan fungsi mereka tanpa sintesis protein tambahan. Namun, sperma yang diejakulasi telah menyimpan RNA messenger (ditranskrip selama spermatogenesis), dengan peran potensial dalam pembuahan dan awal embriogenesis (Ulasan di Jodar et al. [54]). Ada berbagai jenis RNA sperma: RNAs ribosom dan RNAs mitokondria RNA chondrial; RNAs coding dalam fragmentasi atau keadaan atipikal, berbagai kelas RNA kecil tanpa kode (snc RNA), dan lain-lain (elemen dengan unsur intronik, RNA panjang tanpa kode, daerah pendek, unsur berpindah, dan RNA beranotasi tanpa kode [54]. RNA heterogen terkandung dalam sperma yang diejakulasi dapat digunakan sebagai ciri khas analisis genomik untuk menilai kualitas semen (integritas spermatogenesis dan fertilitas) [55,56]. Oleh karena itu, metode yang dapat diandalkan untuk menilai RNA sperma bisa menjadi metode noninvasif baru untuk memprediksi kontribusi sperma dalama fertilisasi dan perkembangan embrio awal. 8.4 Epigenom sperma Epigenom merupakan modifikasi dalam DNA (tanpa mengubah urutan DNA) dan struktur histon yang dapat mempengaruhi ekspresi gen [57]. Penyebab lingkungan (misalnya, racun, nutrisi, dan temperatur) tidak mengubah urutan DNA, tetapi menginduksi modifkasi kimia DNA dan mempengaruhi pemadatan DNA sehingga mempengaruhi akses mesin transkripsi DNA, dan mengubah ekspresi gen. Modifikasi kimia ini termasuk modifikasi histon (asetilasi, fosforilasi, dan metilasi) dan metilasi DNA (penambahan kelompok metil untuk sitosin); yang kemudian dianggap sebagai tanda epigenetik permanen. Mengevaluasi perubahan tanda epigenetik dan perubahan terkait di fenotip mungkin memiliki implikasi untuk mengidentifikasi kontribusi epigenetik terhadap regulasi fertlitas. Sebagai contoh, kami melaporkan bahwa suhu testis yang dinaikkan mengubah ekspresi kohort dari protein di sperma sapi jantan [58], mungkin karena perubahan dalam ekspresi gen melalui modifikasi epigenetik dari DNA sel germinal selama spermatogenesis. Kecacatan metilasi gen dapat mempengaruhi fertilitas sapi jantan [59]. Meningkatnya usia induk berkaitan dengan peningkatan metilisasi secara keseluruhan yang disertai penurunan metilisasi regional yang signifikan dan mempengaruhi kesehatan anak [57]. Oleh karena itu, sapi jantan tua yang digunakan untuk produksi semen komersial harus lebih berhati-hati. Sebuah platform baru untuk studi genom metil pada sapi [60] mungkin sangat berharga untuk menyaring epigenom sperma sapi jantan ang terkena kontaminasi lingkungan, sebelum produksi semen dalam skala besar. 8.5 Sperma proteomik Penelitian sedang berlangsung untuk mengidentifikasi protein sperma yang pentinguntuk meregulasi fertilitas sapi jantan, yang berpotensi sebagai penanda fertilitas. Dengan berfokus pada analisis ekspresi profil, proteomik bisa mengidentifikasi penanda alami fertilitas . Oleh karena protein menentukan fenotip sel, perubahan di tingkat proteome dapat menyebabkan perbedaan fenotipe yang mempengaruhi sifat yang secara ekonomis penting. Massa spektrometri memiliki diaktifkan identifikasi global dan kuantifikasi protein;2D-PAGE dan 2D-DIGE ditambah dengan spektrometri massa telah memfasilitasi identifikasi protein individu dan pasca modifikasi translasi. Meskipun kemajuan dalam resolusi teknologi proteomic, tidak ada ekstraksi protein atau identifikasi metode tunggal dapat mengidentifikasi semua protein seluler, karena perbedaan kisaran dinamika dan kelarutan. Menguraikan sel ke bagian konstituen akan meningkatkan cakupan proteoma dan melokalisasi protein subselular [61]. Permukaan proteome sperma berubah selama pematangan epididimis, kapasitasi, dan fertilisasi. Analisis proteomik pada membran menghasilkan protein W190, ditambah protein permukaan sperma nonraft [62]. Sebuah analisis proteomik komprehensif terhadap membrane yang tahan deterjen mengidentifikasi protein terlibat dalam zona pelusida mengikat dan / atau fusi olemma[63]. Ekor sperma proteome telah dipelajari secara ekstensif; sekumpulan data yang besar (1049 protein di ekor sperma manusia), termasuk beberapa protein peroksimal (organel dianggap sebagian besar absen di sperma matang [64]), sangat menarik. Lebih lanjut, 403 protein yang diidentifikasi dalam inti sperma, dimana lebih dari 50% tidak pernah dilaporkan pada sperma manusia [65]. Jumlah yang tidak memadai dan lokalisasi nuklir protein protamine 1 dikaitkan dengan gangguan struktur kromatin dan tingkat fertilitas yang berkuran pada sapi jantan perah [66].