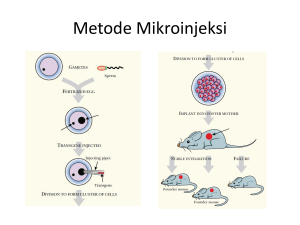
Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari
advertisement

Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana IRAWAN TAN SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2005 SURAT PERNYATAAN Dengan ini saya menyatakan bahwa disertasi yang berjudul: Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana Adalah benar merupakan hasil karya sendiri dan belum pernah dipublikasikan orang lain. Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya. Bogor, Mei 2005 IRAWAN TAN NIM 995205 ABSTRAK IRAWAN TAN. Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana. Dibimbing oleh ANTONIUS SUWANTO (Ketua Komisi Pembimbing), MAGGY THENAWIDJAYA SUHARTONO, MAGDALENA IRENE JOSEPHINE UMBOH (Anggota Komisi Pembimbing) Sintesis prekursor senyawa tetrapirol asam δ-aminolevulinat (ALA) di tanaman dimulai dari glutamat dan merupakan biosintesis tergantung pada tRNA yang terdiri atas tiga tahapan enzimatik yang terjadi di plastida. Pada hewan, khamir dan sejumlah bakteri, ALA dibentuk melalui satu tahapan dari suksinilCoA dan glisin oleh δ-aminolevulinate sintase (ALA-S). Gen yang menyandikan ALA-S (hemA) dari Rhodobacter sphaeroides telah berhasil dikonstruksi di bawah promotor 35S promoter dan diintroduksikan ke dalam genom Arabidopsis thaliana melalui perantaraan Agrobacterium tumefaciens secara in planta. Lima kandidat tanaman transgenik yang dianalisis dengan PCR ternyata empat diantaranya membawa gen hemA dan kanamisin. Hasil analisis transkrip dengan Reverse Transcriptase-PCR menunjukkan keempat tanaman transgenik tersebut terekspresi pada tarap mRNA. Pengukuran berat basah dan berat kering menunjukkan kenaikan tertinggi sebesar 16.8% dan 16.6%. Namun terdapat dua tanaman yang mengalami kelainan yaitu satu mengalami penurunan berat basah dan berat kering akibat perkembangan akar yang jelek (transgenik no 3) sedangkan satu lagi mengalami penurunan jumlah biji dan keterlambatan pembungaan (transgenik no 4). Hasil analisis aktivitas ALAS, total kandungan ALA dan klorofil pada tanaman transgenik no 4 menunjukkan aktivitas sebesar 40.5 nmol.mg-1.h-1, total ALA meningkat 153.6% dan total klorofil meningkat 43% Selain itu juga mengalami peningkatan toleransi terhadap salinitas sampai dengan konsentrasi 200 mM NaCl. Pada penelitian ini juga telah dilakukan kloning gen Chlorophyll A Oxygenase dari Arabidopsis thaliana. ABSTRACT IRAWAN TAN. Expression of δ-aminolevulinate Sintase from Rhodobacter sphaeroides in Arabidopsis thaliana. Supervised by ANTONIUS SUWANTO (Major Advisor), MAGGY THENAWIDJAYA SUHARTONO, MAGDALENA IRENE JOSEPHINE UMBOH (Coadvisors) Synthesis of the tetrapyrrole precursor δ-Aminolevulinate (ALA) in plants starts with glutamate and is a tRNA dependent pathway consisting of three enzymatic steps localized in plastids. In animals, yeast and some of the bacteria, ALA is formed in a single step from succinyl CoA and glycine by aminolevulinate synthase (ALA-S) inside mithochondria. A gene encoding ALA-S from Rhodobacter sphaeroides under 35S promoter was introduced into the genome of Arabidopsis thaliana employing Agrobacterium tumefaciens-mediated transformation in planta. Five putative transgenic lines were obtained and four among them were positive carrying hemA dan kanamycin. All the transgenic lines expressed the transgene at the level of RNA confirmed by RT-PCR analysis. The fresh and dry weigth was increased up to 16.8% and 16.6%, respectively. But, two lines was abnormal, one was reduced in fresh and dry weight because of poor root development (line no 3) and the other reduced in seed production for about one quarter and exhibited late flowering (line no 4). The ALAS activity in line no 4 was 40.5 nmol.mg-1.h-1 and the capacity to synthesize ALA and chlorophyll was increased 153.6% and 43%, respectively. In addition it could also improve tolerance of salinity stress when exposed to 200 mM NaCl. In this study, the gene for Chlorophyll A Oxygenase has been cloned from Arabidopsis thaliana. Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana IRAWAN TAN Disertasi Sebagai salah satu syarat untuk memperoleh gelar Doktor pada Program Studi Biologi SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2005 Judul Disertasi : Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana Nama : IRAWAN TAN NIM : 995205 Menyetujui Komisi Pembimbing Prof. Dr. Antonius Suwanto Ketua (Prof. Dr. Maggy T. Suhartono) Anggota (Prof. Dr. M. Irene J. Umboh) Anggota Mengetahui Ketua Program Studi Biologi Dekan Sekolah Pascasarjana Dr. Dedy Duryadi Solihin Prof. Dr. Syafrida Manuwoto Tanggal Ujian: 11 Juli 2005 Tanggal Lulus: 24 Oktober 2005 PRAKATA Puji syukur penulis panjatkan kehadirat Tuhan YME, atas berkah yang melimpah sehingga penulis dapat menyelesaikan penulisan disertasi dengan judul: Ekspresi Gen Penyandi Asam δ-aminolevulinat Sintase dari Rhodobacter sphaeroides pada Arabidopsis thaliana Pada kesempatan ini penulis mengucapkan terima kasih dan penghargaan yang sebesar-besarnya kepada Prof. Dr. Antonius Suwanto selaku pembimbing utama, yang telah membimbing penulis selama hampir 10 tahun sejak S1. Banyak contoh, pengalaman dan kesempatan serta kepercayaan yang penulis peroleh selama dibimbing Beliau. Penulis juga menyampaikan rasa terima kasih kepada Prof Dr. Maggy T. Suhartono dan Prof. Dr. M. Irene J. Umboh selaku anggota komisi pembimbing, atas semua bimbingan dan arahan sejak awal penelitian sampai penulisan disertasi ini. Penulis juga menyampaikan rasa terima kasih yang sebesar-besarnya kepada Almahumah Dr. Puspa Dewi Tjondronegoro, yang kepada Beliau lah penulis menyampaikan rencana penelitian ini pada akhir tahun 1996 dan sejak saat itu sampai akhir hayatnya, beliau selalu mendukung penulis untuk dapat mengerjakan penelitian ini. Penelitian ini merupakan janji penulis kepada Beliau dan baru saat ini janji tersebut penulis penuhi. Penulis mengucapkan terima kasih kepada Dr. Robert Harling dan The British Council atas kesempatan dan dana yang diberikan untuk melakukan shortterm research di Scottish Agricultural College, University of Edinburgh. Ucapan terima kasih penulis sampaikan kepada The Habibie Center dan Research Center for Microbial Diversity atas bantuan beasiswa dan dana penelitian selama studi S3 ini berlangsung. Kepada semua pihak yang tidak dapat disebutkan satu persatu selama penulis melakukan penelitian baik di Laboratorium Biokimia dan Mikrobiologi, PAU Biotek IPB dan Laboratorium Biologi Molekuler, SEAMEO-BIOTROP, penulis mengucapkan terima kasih atas semua bantuan dan kerjasamanya. Jakarta, Mei 2005 IRAWAN TAN RIWAYAT HIDUP Penulis dilahirkan di Jambi pada tanggal 8 Oktober 1972 dari ayah Eddie Sinatra dan ibu Kastina Tjandra. Pendidikan sarjana di tempuh di Program Studi Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam IPB, lulus pada tahun 1997. Pada tahun 1997, penulis diterima di Program Studi Biologi pada Program Pascasarjana IPB dan menamatkannya pada tahun 1999. Kesempatan untuk melanjutkan ke program doktor pada program studi dan pada perguruan tinggi yang sama diperoleh pada awal tahun 2000. Selama studi program doktor, penulis mendapat beasiswa selama setahun dari The Habibie Center pada tahun 2000 dan Research Center for Microbial Diversity untuk beasiswa dan dana penelitian (2001-2004). Selama mengikuti program S3, penulis pernah menjadi instruktur laboratorium pada workshop “International Training Course on Advances in Molecular Biology Techniques to Assess Microbial Biodiversity I-IV” dan “Lokakarya Teknologi DNA dan Deteksi Produk Rekayasa Genetika pada Bahan Pangan” yang diselenggarakan oleh SEAMEO-BIOTROP dari tahun 2000-2003. Instruktur laboratorium pada workshop “Advances in Molecular Biology Techniques to Assess Microbial Biodiversity, Taxonomy and Detection” tahun 2001 dan “ Pulsed-Field Gel Electrophoresis (PFGE) for DNA Fingerprinting” tahun 2002 yang diselenggarakan oleh Office of Biotechnology Research and Development, Department of Agriculture The Government of Thailand. Sebagai instruktur laboratorium pada “Lokakarya Pengajaran Bioteknologi Modern untuk Guru SMU” tahun 2003 dan 2004 yang diselenggarakan oleh Fakultas Teknobiologi, Unika Atma Jaya. Penulis juga mendapatkan kesempatan melakukan Short-term research di Scottish Agricultural College dari tanggal Agustus – September 2003 yang dibiayai oleh British Council. Penulis juga menjadi instruktur laboratorium pada workshop “Regional Training Course on Utilization of Molecular Marker Techniques in Plant Breeding” pada bulan Mei 2005 yang diselenggarakan oleh SEAMEO-BIOTROP. DAFTAR ISI Halaman DAFTAR TABEL ……………………………………………………. vii DAFTAR GAMBAR ………………………………………………… viii PENDAHULUAN ……………………………………………………. 1 Latar Belakang …………………………………………………… Tujuan ……………………………………………………………. Manfaat Penelitian ………………………………………………. 1 3 3 TINJAUAN PUSTAKA …………………………………………….. 4 Biosintesis asam δ-aminolevulinat (ALA) ………………………. Aplikasi ALA untuk pertanian ………………………………….. Ketahanan terhadap salinitas …………………………………….. Chlorophyll A oxygenase ………………………………………... Transformasi yang diperantarai Agrobacterium …………………. 4 5 8 18 24 BAHAN DAN METODE ……………………………………………. 29 Galur bakteri dan plasmid yang digunakan ………………………. Pembuatan E. coli kompeten……………………………………… Transformasi E. coli ……………………………………………… Purifikasi DNA dari gel agarose …………………………………. Isolasi DNA plasmid ……………………………………………... Konstruksi vektor ekspresi ALAS ……………………………….. Transformasi rekombinan binary vector ke A. tumefaciens EHA 105………………………………………………………………… Transformasi rekombinan binary vector ke Arabidopsis thaliana .. Analisis kandidat tanaman transgenik dengan PCR ……..………. Analisis transkrip dengan RT-PCR ………………………………. Pengukuran kandungan klorofil ………………………………….. Esei aktivitas ALA sintase dan total kandungan ALA ………….. Pengukuran berat basah dan berat kering ………………………... Pengujian ketahanan terhadap salinitas …...……………………… Kloning gen Chlorophyll A Oxygenase dari Arabidopsis thaliana…………………………………………………………… Tempat dan waktu penelitian …………………………………….. 28 29 29 30 30 31 HASIL DAN PEMBAHASAN ……………………………………… 45 Konstruksi vektor ekspresi ALAS ……………………………….. 45 36 37 39 40 42 42 43 43 43 44 Transformasi rekombinan binary vector ke Arabidopsis thaliana . Analisis tanaman Arabidopsis transgenik ……………………….. Ekspresi ALAS meningkatkan kandungan klorofil ……………… Analisis penurunan sifat monogenik Mendel …………………….. Pengujian ketahanan terhadap salinitas …………………………... Kloning gen Chlorophyll A Oxygenase (CAO) ………………….. 47 48 56 57 57 59 SIMPULAN DAN SARAN ………………………………………….. 62 Simpulan …………………………………………………………. Saran ……………………………………………………………… 62 63 DAFTAR PUSTAKA ……………………………………………… 64 DAFTAR TABEL Teks 1. 2. 3. Galur bakteri dan plasmid yang digunakan ……………………. Hasil analisis berat basah dan berat kering Arabidopsis thaliana transgenik dan non transgenik …………………………………. Hasil analisis kandungan klorofil, aktivitas ALAS dan ALA….. Halaman 28 54 56 DAFTAR GAMBAR Teks Halaman 1. Biosintesis asam δ-aminolevulinat (ALA) dan senyawa tetrapirol 4 2. Sensitivitas perkecambahan biji Arabidopsis terhadap NaCl …………………………………………………………………… 3. Sensitivitas tanaman Arabidopsis terhapa NaCl selama fase vegetatif ………………………………………………………….. 4. SOS pathway berperanan dalam homeostasis ion pada saat cekaman salinitas ………………………………..……………… 5. Cekaman salinitas menghambat pengambilan ion K pada Arabidopsis ...……………………………………………………. 6.. Model bagi induksi reactive oxygen species (radikal bebas superoksida, hydrogen peroksida dan hidroksil) yang terjadi pada saat perlakuan dengan NaCl dan peranan enzim antioksidatif superoksida dismutase (SOD), ascorbat peroksidase (APX) dan glutathione peroksidase (GPX) dalam menghancukan radikal bebas superoksida, hydrogen peroksida dan hidroksil ………….. 7. Halliwell-Asada pathway (Siklus Askobat-glutathione) 8. Struktur klorofil A dan klorofil B ……………………………….. 9. Struktur fotosistem II pada keadaan cahaya redup (atas) atau cahaya terang (tengah) pada tanaman tingkat tinggi tipe liar dan mutan tanpa klorofil b (bawah) …………...................................... 10. Sebuah model yang diusulkan untuk siklus klorofil …………….. 11. Model hipotetik proses transfer T-DNA ………………………… 12. Kloning hemA ke plasmid pOK12 menghasilkan plasmid pOKhemA …………………………………………………….……… 13. Konstruksi gen hemA dibawah promoter 35S dengan orientasi sense …………………………………..…………………………. 14. Konstruksi binary vektor yang membawa gen hemA dibawah promoter 35S dengan orientasi sense ………................................. 15. Konstruksi gen hemA dibawah promoter 35S dengan orientasi antisense ……………………………………................................. 16. Konstruksi binary vektor yang membawa gen hemA dibawah promoter 35S dengan orientasi antisense ………………………... 17. Peta plasmid pSOUP …………………………………………….. 18. Tanaman Arabidopsis thaliana yang ditumbuhkan pada tabung PVC (A) dan siap diinfeksi (B) ………………………………….. 19. Tahapan infeksi dengan suspensi Agrobacterium (A) dan pasca infeksi (B) ……………………………………………………….. 20. Hasil analisis restriksi plasmid rekombinan pOK12-hemA, p35S2hemA dan pGII0029-hemA ……………………………………. 21. Hasil analisis restriksi plasmid rekombinan pGII0029-hemA (A) dan pGII0029-AShemA (B) yang didigesti dengan BamHI (3 & 5), EcoRI (1&6) dan HindIII (2&7) ……………………………... 10 11 14 15 17 18 19 19 23 28 32 33 33 34 35 35 38 38 46 46 22. Peta plasmid rekombinan pGII0029-hemA dan pGII0029AShemA …………………………………………………………. 23. Kecambah Arabidopsis yang akan (A) dan setelah (B, C dan D) diseleksi dengan penyemprotan antibiotic kanamisin (in solium selection) ………………………………………………………… 24. Analisis kandidat tanaman transgenik menggunakan PCR dengan primer spesifik gen hemA dan kanamisin resisten ………………. 25. Hasil analisis RT-PCR pada tanaman transgenik T1 ……………. 26. Profil tanaman transgenik no 4 (tanda panah) …………………... 27. Kondisi perakaran tanaman transgenik no 4 (A) dan 3 (B) …… 28. Hasil analisis PCR turunan pertama tanaman transgenik No. 4 … 29. Pengujian ketahanan terhadap cekaman salinitas ………………. 30. Peta plasmid rekombinan pAS900-CAO1 yang membawa gen CAO ……………………………………………………………... 31. Urutan nukleotida gen CAO dari Arabidopsis thaliana ecotype Columbia ………………………………………………………… 47 48 49 50 52 55 57 59 60 60 PENDAHULUAN Latar Belakang Populasi penduduk dunia telah mencapai lebih dari 6 milyar jiwa pada tahun 2000 dan diperkirakan dapat mencapai sekitar 8.5 milyar jiwa pada tahun 2025. Untuk memenuhi kebutuhan pangan dunia dibutuhkan peningkatan hasil yang signifikan dari tanaman pangan utama yang dibudidayakan di negara berkembang. Padi, sebagai contoh, diperkirakan butuh peningkatan hasil sekitar 50% pada tahun 2030 dari yang ada sekarang. Peningkatan potensi hasil akan melibatkan peningkatan biomassa tanaman pangan itu sendiri yang berarti peningkatan produksi asimilat yang berasal dari fotosintesis (Horton 2000). Proses fotosintesis merupakan proses yang sangat penting dalam produksi bahan pangan dan serat. Rata-rata hasil per luas lahan secara global dari tanaman pangan utama seperti gandum, padi dan jagung mengalami peningkatan lebih dari dua kali lipat dalam periode antara 1940 dan 1980 dan kecendrungan ini terus berlanjut. Peningkatan hasil biji-bijian ini sejalan dengan periode dimana pengertian kita tentang fotosintesis telah meningkat secara luar biasa. Kemajuan dalam penelitian fotosintesis ini berlanjut dengan terobosan baru yang dibawa oleh kemajuan dalam bidang biologi molekuler (Richards 2000). Sejumlah kondisi lingkungan memperlihatkan pengaruhnya terhadap pertumbuhan dan perkembangan serta produktivitas tanaman. Tanaman menghadapi berbagai tekanan baik secara biotik (hama dan penyakit) dan abiotik (salinitas, kekeringan, suhu yang ekstrem dll). Dalam sistem pertanian, tekanan abiotik, salinitas, suhu dingin dan kekeringan merupakan faktor yang berperanan penting dalam menurunnya potensial hasil dari berbagai tanaman pertanian (Boyer, 1982). Asam δ-aminolevulinat (ALA) merupakan prekursor kunci dalam biosintesis porfirin seperti klorofil dan heme. Sejauh ini, ALA diketahui mempunyai banyak kegunaan bukan saja sebagai herbisida (Rebeiz et al. 1984), insektisida (Rebeiz et al. 1988) dan faktor pemacu pertumbuhan dengan meningkatkan fiksasi CO2 dalam keadaan terang dan menurunkan pelepasan CO2 pada keadaan gelap (Hotta et al. 1997a, b), tetapi juga kemampuannya untuk meningkatkan toleransi tanaman terhadap stres oleh salinitas (Watanabe et al. 2000) dan temperatur dingin (Hotta and Watanabe 1999, Wang et al. 2004). Yoshida et al. (2004) melaporkan bahwa ALA pada konsentrasi 30-100 ppm dapat meningkatkan pertumbuhan dan penyerapan nitrogen pada tanaman Komatsuna yang ditanam di tanah alkalin yang apabila tidak diberi perlakuan ALA dapat menurunkan hasil sampai 50%. ALA juga diketahui mempunyai aktivitas hormonal seperti auksin dan sitokinin dalam induksi kalus dan rhizogenesis serta pertunasan (Bindu and Vivekanandan 1998a). Biosintesis ALA secara biologis dilakukan melalui dua jalur yang berbeda: (1) melalui suksinil-CoA dan glisin (Jalur Shemin atau Jalur C-4), dan (2) dari glutamat (Jalur C-5). Tumbuhan tingkat tinggi, briofita, sianobakter dan sebagian eubakteria menggunakan jalur C-5 untuk biosintesis ALA yang melibatkan tiga macam enzim yaitu Glutamil-tRNA sintase, Glutamil-tRNA reduktase dan Glutamat semialdehida aminotransferase. Sedangkan hewan, khamir dan sejumlah bakteri kelompok α-proteobacteria seperti Rhodobacter, Rhodospirillum dan Rhizobium menggunakan jalur biosintesis C-4 yang melibatkan hanya satu enzim yaitu ALA sintase (von Wettstein, Gough, and Kannangara 1995). Introduksi gen ALA sintase dari jalur C-4 ke tanaman (C-5) diharapkan dapat memperpendek jalur biosintesis selain efek lain yang diinginkan seperti telah disebutkan diatas. Dalam melakukan penelitian pada bidang biologi molekular dan rekayasa genetika tanaman dibutuhkan suatu sistem atau tanaman model untuk mempelajari ekspresi suatu gen dalam tanaman. Untuk keperluan ini Arabidopsis thaliana telah menjadi tanaman model yang secara luas digunakan dalam studi biologi tanaman (Meinke et al., 1998), termasuk didalamnya transformasi genetik. Hal ini dikarenakan Arabidopsis thaliana mempunyai ukuran genom yang kecil (125 Mb), siklus hidup yang pendek (sekitar 6 minggu sejak dikecambahkan) dan menghasilkan biji yang relatif banyak dari satu siklus Satu hal penting adalah telah berhasil diselesaikannya sekuen total genom tanaman ini pada tahun 2000 dan pengembangan metode transformasi yang efisien sehingga memungkinkan studi ekspresi gen secara lebih lengkap (TAIR, 2003). Tujuan Penelitian ini bertujuan untuk mempelajari ekspresi gen penyandi asam δaminolevulinat sintase (hemA) pada Arabidopsis thaliana. Manfaat Penelitian Penelitian ini diharapkan dapat meningkatkan performa pertumbuhan tanaman melalui peningkatan laju fotosintesis, ketahanan cekaman salinitas dan ketahanan tanaman terhadap naungan. Informasi yang diperoleh dari penelitian ini diharapkan dapat diterapkan pada tanaman pangan utama. TINJAUAN PUSTAKA Biosintesis asam δ-aminolevulinat Cincin porfirin merupakan komponen dari berbagai molekul biologis penting termasuk diantaranya haemoglobin, sitokrom, vitamin B12 dan klorofil. Prekursor kunci dari cincin porfirin ini adalah asam δ-aminolevulinat (ALA). Biosintesis ALA dapat dilakukan melalui dua jalur: (1) dari suksinil-CoA dan glisin (Shemin pathway, C-4 pathway), dan (2) dari glutamat (C-5 pathway) seperti yang tersaji dalam Gambar 1. Gambar 1. Biosintesis asam δ-aminolevulinat (ALA) dan senyawa tetrapirol (Sasaki et al. 2002). Pada jalur Shemin, suksinil-CoA disediakan melalui siklus TCA. Suksinil-CoA dan glisin dikondensasi dengan bantuan enzim ALA sintase (ALAS). Jalur biosintesis ini digunakan oleh hewan, khamir dan sejumlah bakteri dalam kelompok α-proteobacteria seperti Rhodobacter, Rhodospirillum dan Rhizobium. Di sejumlah bakteri, suksinil-CoA disintesis dari propionil-CoA melalui jalur biosintesis metilmalonil-CoA (Sasaki et al. 1990). ALAS merupakan enzim pembatas kunci untuk biosintesis senyawa tetrapirol, dan sintesis enzim ini sendiri diatur secara ketat (Sasikala and Ramana 1995) melalui pengaturan secara umpan balik pada gen penyandi HemA atau HemT (Neidle and Kaplan 1993). Jalur biosintesis C-5 merupakan karakteristik dari tumbuhan tingkat tinggi, Briofita, Sianobakteri dan sejumlah Eubakteria. Jalur biosintesis ini melibatkan tiga buah enzim yaitu Glutamat-tRNA sintase, Glutamil-tRNA reduktase (gtr A/hemA), dan Glutamat 1-semialdehida aminotransferase (hemL). Pada tumbuhan tingkat tinggi, gen penyandi tRNAGlu terdapat di DNA kloroplas, sedangkan ketiga macam enzim yang terlibat dalam biosintensis ALA disandikan oleh DNA inti dan diimpor ke dalam stroma kloroplas setelah disintesis oleh ribosom sitoplasma. Pada fitoflagelata Euglena gracilis ditemukan adanya dua jalur biosintesis (C-4 dan C-5) yang dioperasikan di dalam organel yang berbeda. Jalur C-5 beroperasi di kloroplas dan secara eksklusif bertanggung-jawab untuk sintesis klorofil, sementara di mitokondria, ALA sintase bertanggung-jawab untuk sintesis heme a dari sitokrom c oksidase (Weinstein and Beale 1983). Aplikasi ALA untuk pertanian Aplikasi praktis ALA sebagai bioherbisida pertama kali di demonstrasikan oleh Rebeiz et al. (1984). Hasil yang dicapai sangatlah berarti karena ALA dapat bekerja secara selektif, tidak berbahaya dan merupakan bahan yang dapat diuraikan secara biologis. Mekanisme ALA sebagai “photodynamic herbicide” seperti diutarakan oleh Rebeiz et al. (1984) sebagai berikut: tanaman yang diberi perlakuan dengan ALA dengan jumlah tinggi akan mengakumulasi secara berlebihan molekul protoporfirin IX (PPIX) pada tahap biosintesis klorofilnya. Ketika tanaman terkena cahaya, PPIX yang berlebih akan menghasilkan radikal bebas (O2-), yang akan mengoksidasi asam lemak tak jenuh pada membran sel dan pada akhirnya merusak tanaman. Oleh karena itu ALA dapat digunakan sebagai bahan pengganti yang aman untuk herbisida yang sangat toksik seperti Paraquat. Mekanisme yang sama juga ditunjukkan oleh ALA jika digunakan sebagai bioinsektisida untuk membasmi Trichopusia ni (Rebeiz et al. 1988). Namun dilain pihak, telah banyak penelitian yang melaporkan efek yang menguntungkan terhadap pertumbuhan berbagai jenis tanaman jika ALA diaplikasikan dalam jumlah rendah (30-100 ppm), seperti meningkatkan berat kering tanaman, peningkatan aktivitas fotosintesis dan efek penghambatan atas respirasi serta mempunyai aktivitas hormonal dalam induksi kallus dan mikropropagasi (Tanaka et al. 1992; Yoshida et al. 1996a; 1996b; Bindu and Vivekanandan 1998a; 1998b; Yoshida et al. 2004). Watanabe et al. (2000) menemukan bahwa aplikasi ALA pada konsentrasi rendah (100 ppm) meningkatkan toleransi terhadap salinitas pada tanaman kapas muda. Hal yang sama juga ditemukan pada tanaman bayam yang diberi perlakuan 0.6 dan 1.8 mM ALA pada media yang mengandung 50 dan 100 mM NaCl, ternyata bukan saja dapat tumbuh dengan baik, yang berarti tanaman menjadi toleran terhadap NaCl, tetapi juga menyebabkan meningkatnya laju fotosintesis (Nishihara et al. 2003). Pada barley, pemberian ALA 30-100 ppm dapat menyebabkan peningkatan hasil sampai 40% yang disebabkan oleh meningkatnya jumlah biji yang dihasilkan tanpa mempengaruhi berat biji. Hal yang sama juga diperlihatkan oleh tanaman gandum dan padi. Tanaman kacang merah yang diberi perlakuan ALA pada fase awal seperti fase daun primer atau daun pertama, meningkat hasilnya sampai 2030%, sedangkan perlakuan pada fase pembungaan menurunkan hasil sekitar 10%. Dalam hal ini hasil yang dicapai sangat tergantung pada fase pertumbuhan mana tanaman diberi perlakuan dengan ALA. Pada tanaman bawang dan kentang pemberian ALA pada fase pembentukan umbi dapat meningkatkan jumlah maupun berat umbi yang terbentuk. Wang et al. (2004) melaporkan bahwa jika kecambah melon (Cucumis melo L. Ximiya No. 1) yang ditumbuhkan dengan intensitas penyinaran 150 µmol m-2 s-1 dan diberi aplikasi larutan ALA dapat meningkatkan secara signifikan laju fotosintesis netto (Pn), jumlah CO2 yang difiksasi per jumlah foton (AQY), Efisiensi karboksilasi (CE) dan konduktansi stomata (Gs). Setelah pemberian dengan 10 ml larutan ALA (10 mg l-1 atau 100 mg l-1) per wadah yang diisi dengan 250 g pasir bersih selama 3 hari, Pn daun sekitar 40-200% lebih tinggi dibandingkan kontrol, dan AQY, CE and Gs meningkat secara berturut-turut sekitar 21-271%, 55-210% dan 60-335%. Lebih lanjut, perlakuan ALA meningkatkan kandungan klorofil daun dan tingkat gula terlarut demikian juga dengan laju respirasi dalam keadaan gelap, tetapi menurunkan laju respirasi dalam keadaan terang. Dilain pihak, Setelah kecambah melon yang ditumbuhkan di dalam ruang pertumbuhan diberi perlakuan pendinginan pada suhu 8°C selama 4 jam dan dikembalikan lagi ke suhu 25-30°C selama 2 dan 20 jam, Pn tanaman yang diberi air (kontrol) hanya sekitar 12-18% dan 37-47%, dibandingkan dengan Pn awal sebelum diberi perlakuan pendinginan. Jika kecambah dengan diberi perlakuan sama tetapi diberi ALA (10 mg l-1), Pnnya berturut-turut sekitar 22-38% and 76-101%, dibandingkan dengan kontrol sebelum pemberian cekaman dingin. Jika lama pendinginan menjadi 6 jam, tanaman yang diberi perlakuan ALA hanya memperlihatkan sedikit gejala nekrosis pada daun sedangkan semua tanaman yang hanya diberi air akan mati, hal ini menyimpulkan bahwa ALA meningkatkan torelansi tanaman terhadap stres dingin dibawah kondisi cahaya rendah. Ketahanan terhadap salinitas Tanaman membutuhkan mineral nutrien esensial untuk tumbuh dan berkembang. Namun demikian, kelebihan garam-garam mineral yang terlarut di tanah dapat berbahaya bagi kebanyakan tanaman. Cekaman salinitas merupakan salah satu faktor lingkungan penting yang membatasi pertumbuhan dan produktivitas tanaman pertanian di seluruh dunia. Di perkirakan bahwa masalah salinitas mempengaruhi setidaknya 20% lahan yang dapat ditanami di seluruh dunia dan lebih dari 40% lahan beririgasi dengan berbagai tingkat kerusakan (Rhoades and Loveday 1990). Pada kasus yang ekstrem, produktivitas lahan pertanian tidak dapat lagi menunjang kelangsungan produksi pertanian dan harus ditinggalkan. Di daerah pantai invasi air laut secara periodik langsung menambahkan garam ke tanah. Tanah di daerah semi-arid atau arid, khususnya dengan sistem drainase yang jelek, akumulasi garam sebagai akibat evaporasi air irigasi, meninggalkan deposit garam garam terlarut. Berdasarkan kemampuannya untuk tumbuh di tempat berkadar garam tinggi, secara tradisional tanaman dikelompokkan atas glikofita dan halofita (Flowers et al. 1977). Halofita toleran terhadap konsentrasi NaCl yang tinggi; beberapa dapat bertahan pada kadar garam dua kali konsentrasi garam pada air laut. Sebagian besar tanaman, termasuk mayoritas spesies tanaman pertanian masuk ke dalam kelompok glikofita dan tidak dapat mentolerir salinitas yang tinggi. Untuk glikofita, salinitas menyebabkan cekaman ionik, osmotik dan cekaman lanjutan seperti penghambatan pengambilan ion dan cekaman radikal oksida (Zhu 2001a). Toksisitas oleh natrium merupakan cekaman ionik utama yang berhubungan dengan salinitas tinggi. Sebagai tambahan, sejumlah tanaman juga sensitif terhadap klorida, anion utama yang ditemukan pada tanah salin. Pada tanah salin tertentu, toksisitas ion diperparah oleh pH alkalin. Potensial osmotik yang rendah dari larutan salin menghambat pengambilan air dan menyebabkan kekeringan fisiologis. Untuk tanaman halofita yang toleran terhadap toksisitas natrium, cekaman osmotik merupakan penyebab utama terhambatnya pertumbuhan. Pemahaman atas mekanisme toleransi tanaman terhadap salinitas akan membantu secara efektif proses pemuliaan atau rekayasa genetika tanaman yang tahan terhadap salinitas. Beberapa aspek dalam respon tanaman terhadap cekaman salinitas berhubungan sangat erat dengan respon terhadap cekaman kekeringan dan dingin (Zhu 2001b). Studi tentang toleransi tanaman terhadap salinitas akan memberikan pemahaman tentang mekanisme toleransi silang pada cekaman abiotik. Arabidopsis thaliana merupakan tanaman glikofita yang sensitif terhadap cekaman salinitas dimana dapat terjadi penghambatan pertumbuhan dan kerusakan jaringan yang menuju kematian. Seperti kebanyakan glikofita, sensitivitas Arabidopsis thaliana terhadap cekaman salinitas diperlihatkan pada semua fase perkembangannya. Sebagai contoh, perlakuan selama 8 jam dengan 150 mM NaCl pada fase pembentukan biji menyebabkan terjadinya deposisi callosa dan perubahan yang abnormal pada struktur ovul dan embrio yang mengindikasikan kematian sel (Sun and Hauser 2001). Sensitivitas Arabidopsis thaliana terhadap cekaman salinitas sangat terlihat pada fase perkecambahan biji dan kecambahnya. Perkecambahan biji Arabidopsis thaliana sangat terganggu pada konsentrasi 75 mM NaCl atau lebih (Gambar 2). Pertumbuhan kecambah juga sangat sensitif terhadap NaCl. Bahkan pada konsentrasi NaCl yang lebih rendah (< 50 mM NaCl) dapat secara nyata mempengaruhi berat basah sedangkan pada konsentrasi yang lebih dari 50 mM NaCl akan menghambat pertumbuhan (Gambar 3) dan bahkan dapat menyebabkan kematian. Hari ke 2 Hari ke 4 Gambar 2. Sensitivitas perkecambahan biji Arabidopsis terhadap NaCl. Biji ecotype C24 (Salah satu ecotype yang sensitif NaCl) ditumbuhkan pada kertas filter yang telah dibasahi dengan larutan NaCl dan diinkubasi pada suhu 4°C selama 2 hari sebelum dipindahkan ke suhu ruang (23°C) dibawah penyinaran cahaya putih untuk perkecambahan (Xiong and Zhu 2002). Minggu ke-3 Minggu ke-4 Gambar 3. Sensitivitas tanaman Arabidopsis terhadap NaCl selama fase vegetatif. Kecambah berumur dua minggu (ecotype Columbia) ditumbuhkan ditanah yang di beri larutan 0, 50, 75, dan 100 mM NaCl. Dokumentasi diambil pada minggu ke tiga (Panel atas) dan minggu ke empat (Panel bawah) setelah perlakuan (Xiong and Zhu 2002) Tanaman yang diadaptasikan dengan konsentrasi garam rendah dapat meningkatkan toleransi tanaman terhadap cekaman salinitas menunjukkan bahwa tanaman glikofita memiliki perangkat untuk membuatnya toleran terhadap salinitas yang mungkin tidak berkerja secara efektif pada keadaan yang tidak teradaptasi. Oleh karena itu, perbedaan toleransi terhadap salinitas antara glikofita dan halofita lebih pada tingkat kuantitatif daripada kualitatif, dan mekanisme dasar toleransi terhadap salinitas kemungkinan bersifat konservatif pada semua spesies tanaman (Xiong and Zhu 2002). Sebagai contoh, gen vakuolar Na+/H+ antiporter AtNHX1 tidak indusibel dibandingkan dengan gen yang homolog pada halofita, dan tingkat ekspresi yang tinggi gen AtNHX1 menggunakan promoter kuat 35S CaMV dapat secara nyata meningkatkan toleransi Arabidopsis terhadap salinitas (Apse et al.1999; Hamada et al. 2001). Ion natrium sebenarnya tidak dibutuhkan dalam pertumbuhan oleh sebagian besar tumbuhan. Bahkan tumbuhan sepertinya tidak memiliki sistem transpor khusus untuk pengambilan Na+. Namun demikian, Na+ masih dapat memasuki sel tanaman melalui beberapa jalur. Karena konsentrasi Na+ di tanah biasanya lebih tinggi dibandingkan dengan yang ada di sitosol sel akar, pergerakan Na+ kedalam sel akar dapat terjadi secara pasif. Bukti yang didapat baru-baru ini menunjukkan bahwa sebagian besar Na+ memasuki sel akar melalui saluran kation tidak tergantung voltase (VIC) atau yang dikenal dengan nama lain saluran kation tidak selektif (NSCC) (Amtmann and Sanders 1999; Demidchik and Tester 2002). Selain itu, disebabkan oleh kemiripan antara ion Na+ and K+, transporter K+ HKT1 kemungkinan merupakan pintu masuknya ion Na+ ke dalam sel akar karena transporter ini mempunyai afinitas terhadap Na+ pada saat konsentrasi Na+ diluar sel tinggi (Rubio et al. 1995). Gen yang homolog pada Arabidopsis, AtHKT1, memperantarai pengambilan Na+ ketika diekspresikan di Saccharomyces cerevisiae atau oosit Xenopus (Uozumi et al. 2000). Mutan supresor hkt1 mempunyai kandungan Na+ yang lebih rendah, menunjukkan bahwa AtHKT1 memerantarai pengambilan Na+ kedalam tanaman. Studi ini juga menunjukkan bahwa SOS3 pada tipe liar, bekerja bersama-sama dengan SOS2, mungkin menekan aktivitas AtHKT1 dalam memungkinkan Na+ masuk kedalam sel akar (Gambar 4). Tanaman mempunyai sejumlah cara untuk mengatasi cekaman ionik yang diakibatkan oleh salinitas yang tinggi termasuk diantaranya membatasi pengambilan, meningkatkan pengeluaran dan lokalisasi Na+ dalam vakuola dan mengontrol transport jarak jauh ke daerah daun. Kelebihan Na+ pada permukaan akar akan menganggu proses pengambilan K+ yang sangat penting dalam menjaga turgor sel, potensial membran dan aktivitas sejumlah enzim (Lazof and Bernstein 1999). Karena kemiripan sifat fisiko-kimia Na+ dan K+, Na+ pada konsentrasi tinggi mempunyai efek inhibisi yang kuat pada proses pengambilan ion kalium oleh akar. Sebagai contoh, pengambilan K+ melalui KUP1 Arabidopsis dihambat oleh konsentrasi 5 mM NaCl atau lebih tinggi (Kim et al. 1998; Fu and Luan 1998). Tanaman mengunakan sistem pengambilan K+ baik yang berafinitas tinggi maupun rendah. Ion natrium mempunyai efek yang lebih merusak pada sistem yang berafinitas rendah yang memiliki selektivitas K+/N+ yang rendah. Pada keadaan cekaman salinitas, merupakan hal yang penting bagi tanaman untuk mengoperasikan sistem pengambilan K+ yang berafinitas tinggi dalam rangka menjaga ketersediaan K+ yang dibutuhkannya. Sudah merupakan fenomena yang umum bahwa perlakuan NaCl pada tanaman menyebabkan menurunnya kandungan K+ (Gambar 5), yang secara parsial bertanggung jawab terhadap menurunnya pertumbuhan dan vigor tanaman dalam keadaan cekaman salinitas. [Na+] Tinggi Vakuola Regulasi gen pada taraf transkripsi dan pasca transkripsi Gambar 4. SOS pathway berperanan dalam homeostasis ion pada saat stres oleh salinitas. Konsentrasi NaCl ekstraselular yang tinggi akan memicu meningkatnya konsentrasi Ca2+ di sitosol. Sensor Ca2+ SOS3 yang menanggapi signal Ca2+ ini akan berinteraksi dan mengaktifkan protein kinase SOS2. SOS2 yang teraktivasi kemudian akan mengatur aktivitas transporter ion atau aktivator transkripsional untuk menjaga kesetimbangan ion atau ekspresi gen. Target protein SOS2 termasuk Na+/H+ antiporter (SOS1), penukar Na+/H+ vakuola ( NHX), dan Na+ transporter (HKT1). Target potensial lainnya termasuk ATPase tonoplas dan pirofosfatase, saluran air, dan kalium transporter (Zhu 2003). .Pengeluaran Na+ dari sel merupakan cara cepat untuk menghindari akumulasi ion natrium di dalam sitosol. Di perkirakan bahwa pengeluaran ion natrium mungkin lebih penting pada sel tertentu, misalnya sel epidermal akar. Hal ini disebabkan karena sebagian besar sel lain di tanaman dikelilingi sel tetangganya dan pengeluaran Na+ akan menimbulkan masalah bagi sel tetangganya. Pada analisis ekspresi promoter SOS1-GUS menunjukkan bahwa gen Na+/ H+ antiporter terekspresi dengan baik hanya pada sel epidermal yang mengelilingi ujung akar dan di sel parenkima yang menyelubungi xylem diseluruh tanaman (Shi et al. 2002a). Pada Arabidopsis thaliana, ekstrusi Na+ terjadi melalui Na+/ H+ antiporter SOS1 yang terlokalisasi di membran plasma (Shi et al. 2000a; Shi et al. 2002b). Mutasi pada SOS1 menyebabkan tanaman mutan menjadi sangat sensitif terhadap ion natrium (Wu et al. 1996). [Ion] (%) NaCl (mM) Gambar 5. Stres oleh salinitas menghambat pengambilan ion K pada Arabidopsis. Dengan meningkatnya NaCl di dalam medium kultur, kandungan Na+ di dalam tanaman meningkat sedangkan kandungan K+ menurun. Kecambah Arabidopsis (ecotype Columbia) ditumbuhkan di larutan ½MS (pH 5.3) selama 2 minggu yang diberi perlakuan NaCl. Kecambah ditumbuhkan selama 3 hari sebelum dipanen dan dianalisa kandungan ionnya (berdasarkan berat kering) (Xiong and Zhu 2002) Toleransi terhadap cekaman salinitas merupakan suatu karakter yang komplek melibatkan berbagai respon selular terhadap cekaman osmotik dan ionik, demikian juga efek cekaman sekunder. Banyak penelitian yang mempelajari berbagai efek cekaman salinitas dan melindungi tanaman dari reactive oxygen species (ROS) sepertinya merupakan salah satu komponen penting dari karakter tanaman yang toleran. Konsentrasi NaCl yang tinggi biasanya akan mengganggu transpor elektron selular pada berbagai organel subselular dan menyebabkan terbentuknya ROS seperti singlet oxygen, superoksida, hidrogen peroksida dan radikal hidroksil (Noctor and Foyer 1998). Kelebihan ROS memicu reaksi fitotoksik seperti peroksidasi lipid, degradasi protein dan mutasi DNA. Sumber utama ROS adalah radikal superoksida, yang dibentuk diberbagai organel subselular seperti mitokondria, kloroplas dan sitoplasma melalui sejumlah jalur metabolik selama kondisi cekaman oksidatif (Noctor and Foyer 1998). Tingkat kerusakan sel akibat proses peroksidasi ditentukan oleh potensi sistem enzim antioksidatif (Gambar 6). Untuk mengontrol jumlah ROS dan melindungi sel dibawah kondisi cekaman, sel tanaman mempunyai sejumlah enzim untuk mendetoksikasi ROS (superoksida dismutase, catalase, peroksidase dan glutathion peroksidase), detoksikasi produk peroksidasi lipid (glutathion S-transferase, phospholipidhidroperoksida glutathion peroksidase dan askorbat peroksidase) dan antioksidan berberat molekul rendah (antosianin, α-tokoferol, askorbat, glutathion dan senyawa polifenol) serta seluruh enzim yang dibutuhkan untuk meregenerasi bentuk aktif dari antioksidan (monodehidroaskorbat reduktase, dehidroaskorbat reduktase dan glutathion reduktase, Gambar 7 (May et al. 1998; Blokhina et al. 2003)). Di dalam sel, superoksida dismutase (SOD) merupakan pertahanan pertama terhadap ROS yang mengkatalisis dismutasi anion superoksida menjadi oksigen dan hidrogen peroksida (H2O2). O2- dihasilkan disetiap tempat dimana terdapat rantai transfer elektron dan oleh karena itu aktivasi O2 dapat terjadi di organel yang berbeda (Elstner 1991), termasuk mitokondria, kloroplas, mikrosom, glikosisom, peroksisom, apoplas dan sitosol. Oleh karena itu SOD dapat ditemukan di semua lokasi subselular. Kloroplas, mitokondria dan peroksisom merupakan tempat utama bagi pembentuk ROS (Fridovich 1986). Berdasarkan penggunaan kofaktor logam oleh enzim, SOD dapat dikelompokkan menjadi tiga: Fe-SOD, Mn-SOD dan Cu-Zn SOD dan ketiga SOD ini terletak di kompartemen sel yang berbeda. Fe-SOD terdapat di kloroplas, Mn-SOD di mitokondria dan peroxisome, dan Cu-Zn SOD terdapat di kloropls, sitosol dan kemungkinan di ruang ekstraselular (Alscher et al. 2002). Untuk mendetoksikasi H2O2 yang terbentuk, tanaman mengembangkan enzim antioksidatif seperti peroksidase dan katalase. Peroksidase tanaman menggunakan substrat yang berbeda seperti guaikol, askorbat dan glutathion untuk mendetoksikasi H2O2 intraselular. Berdasarkan spesifisitas substrat peroksidase dikelompokkan menjadi guaikol peroksidase, askorbat peroksidase and glutathion peroksidase. H2O2 yang dihasilkan di glyoxysom dan peroxisom dalam proses βoksidasi didetoksikasi menjadi H2O oleh enzim katalase, sedangkan di organel subselular lain H2O2 dikonversi menjadi H2O oleh enzim askorbat peroksidase dan glutathion peroksidase (Halliwell and Gutteridge 1989; Sen-Gupta et al. 1993). . NaCl Radikal Superoksida SOD Hidrogen peroksida APX GPX H2O + O2 Gambar 6. Model bagi induksi reactive oxygen species (radikal superoksida, hydrogen peroksida dan hidroksil) yang terjadi pada saat perlakuan dengan NaCl dan peranan enzim antioksidatif superoksida dismutase (SOD), askorbat peroksidase (APX) dan glutathion peroksidase (GPX) dalam mendetoksikasi radikal bebas superoksida, hidrogen peroksida dan hidrosil (Xiong and Zhu 2002). Gambar 7. Halliwell-Asada pathway (Siklus Askorbat-glutathione). APX, ascorbat-peroksidase; MDHAR, monodehidroaskorbat reduktase; DHAR, dehidroaskorbat reduktase; GR, glutathion reduktase (May et al 1998) Chlorophyll A Oxygenase (CAO) Klorofil merupakan inti dari proses fotosintesis dalam pemanenan energi cahaya dan pengkonversiannya menjadi energi kimia. Tumbuhan darat, algae hijau dan proklorofita menghasilkan klorofil a dan klorofil b (Gambar 8). Klorofil a sendiri terdapat di pusat reaksi dari komplek fotosistem yang memiliki komposisi dan organisasi yang sangat konservatif. Di lain pihak, klorofil a dan klorofil b merupakan komponen antena penangkap cahaya periferal. Komplek antena penangkap cahaya memperlihatkan perubahan yang terkontrol dalam beradaptasi terhadap berbagai kondisi pertumbuhan, yang memungkinkan penggunaan yang optimal dari cahaya yang tersedia. Diketahui bahwa rasio klorofil a dan b tinggi pada kondisi pertumbuhan dengan cahaya terang dibandingkan pada kondisi pertumbuhan cahaya redup, dimana terjadi pembesaran ukuran komplek antena penangkap cahaya pada kondisi cahaya redup (Gambar 9) (Tanaka and Tanaka 2005). Oleh karena itu regulasi biosintesis klorofil b menjadi sangat penting dalam memahami mekanisme adaptasi tanaman terhadap berbagai intensitas cahaya. Klorofil a Klorofil b Gambar 8. Struktur klorofil A dan klorofil B. Perbedaan terletak pada gugus dalam kotak merah. Pada klorofil a berupa gugus metil sedangkan klorofil b aldehida A. Protein-protein pusat reaksi (Chl a) B. Protein-protein antena utama (Chl a) C. Protein-protein antena periferal minor (Chl a, Chl b) D. Protein-protein antena periferal mayor (Chl a, Chl b) Kondisi intensitas cahaya rendah Kondisi intensitas cahaya tinggi Mutan minus Chl b Gambar 9. Struktur fotosistem II pada keadaan cahaya redup (atas) atau cahaya terang (tengah) pada tanaman tingkat tinggi tipe liar dan mutan tanpa klorofil b (bawah). Terdapat dua hipotesis mengenai regulasi ukuran antena penangkap cahaya: regulasi oleh ekspresi gen-gen Lhc dan regulasi oleh biosintesis klorofil b. Hipotesis pertama berdasarkan penemuan bahwa pola ekspresi gen-gen tersebut berhubungan erat dengan ukuran antena (Maxwell et al. 1995; Escoubas et al. 1995). Namun demikian, terdapat sejumlah bukti yang berlawanan dengan hipotesis pertama dan cenderung mendukung hipotesis kedua. Pertama, pada sejumlah mutan chl b-less, lebih banyak protein LHC yang hilang ketika jumlah klorofil b menurun. Pada Arabidopsis thaliana mutan yang tidak bisa sama sekali membentuk klorofil b, tidak ada protein LHC yang terbentuk (Espineda 1999), sementara itu di barley mutan yang masih mengandung sejumlah kecil klorofil b, satu atau lebih protein masih dapat ditemui (Bossmann et al 1997). Proses transkripsi, translasi dan transpor protein LHC berjalan normal, menunjukkan bahwa stabilitas protein LHC yang mengalami gangguan (Bellemare, 1982). Kedua, prekursor protein LHC membutuhkan klorofil b untuk kestabilan insersi ke dalam membran (Kuttkat et al. 1997). Ketiga, telah dibuktikan bahwa mRNA Lhc dengan jumlah kurang dari 5% dari keadaan normalnya sudah cukup untuk menghasilkan ukuran antena yang terbesar sekalipun. Flachman and Kühlbrandt (1997) dan Flachman (1995) melakukan transformasi tanaman tembakau dengan antisense gen Lhc. Di tanaman ini ekspresi gen Lhc mengalami tekanan oleh ekspresi antisensenya. Uniknya, tingkat protein LHC tidak mengalami perubahan bahkan pada tanaman transgenik mengekspresikan 5% mRNA dengan orientasi sense. yang hanya Hasil ini menunjukkan bahwa klorofil b dibutuhkan untuk kestabilan dan/atau insersi protein LHC ke dalam membran dan transkripsi gen-gen Lhc bukanlah faktor utama yang mengatur ukuran antena penangkap cahaya. Transformasi Arabidopsis dengan gen CAO dibawah kontrol promotor 35S CaMV menunjukkan bahwa ukuran antena pada fotosistem II meningkat antara 10-20% jika dibandingkan dengan tanaman tipe liarnya, hal ini memperlihatkan bahwa biosintesis klorofil b dapat mengatur ukuran antena penangkap cahaya pada tanaman (Tanaka et al. 2001). Sebelum gen yang bertanggung-jawab dalam biosintesis klorofil b ditemukan, sangat sedikit yang diketahui tentang jalur biosintesis klorofil ini. Identifikasi gen untuk biosintesis klorofil b dilakukan dengan mutagenesis insersi pada Chlamydomonas reinhardtii yang menghasilkan sejumlah chl b-less mutan. Hasil analisis genom yang termutasi berhasil mengidentifikasi sebuah gen yang diperkirakan suatu monooxygenase yang memiliki [2Fe-2S] Rieske center motif dan mononuclear iron-binding motif (Tanaka et al. 1998). Kloning gen homolog yang dilakukan menggunakan PCR dan pencarian dari database pada alga laut prokariotik, Prochlorothrix dan Prochlorococcus, dan tanaman tingkat tinggi, Arabidopsis thaliana menunjukkan bahwa gen ini konservatif dari prokariot sampai tanaman tingkat tinggi (Tomitani et al. 1999). Protein dari gen yang diklon dari Arabidopsis thaliana dan diproduksi di Escherichia coli mengkatalisis dua tahapan reaksi dari chlorophyllide (Chlide) a menjadi Chlide b (Chlorophyllide merupakan prekursor tidak teresterifikasi dari klorofil). Berdasarkan aktivitas ini, enzim ini dinamakan CAO (Chlorophyllide a monooxygenase) (Tanaka and Tanaka 2005). Hal yang menarik adalah bahwa enzim ini hanya mengenali chlorophyllide a, artinya bahwa klorofil a pertama- tama harus di de-esterifikasi kembali menjadi clorophyllide a dan kemudian dikonversi menjadi clorophyllide b dan kemudian diesterifikasi kembali menjadi klorofil b (Oster et al. 2000). Penemuan yang tidak terduga lainnya adalah bahwa CAO mengkatalisis dua tahapan reaksi. Tahapan pertama adalah oksigenasi gugus metil menjadi gugus hidroksil menghasilkan 7-hydroxymethyl-chlorophyll. Pada tahap kedua gugus hidroksil dioksigenasi lagi menjadi gugus formil menghasilkan klorofil b. Kedua tahapan ini membutuhkan NADPH dan ferredoxin sebagai tenaga pereduksi (Tanaka and Tanaka 2005). Walaupun sudah secara jelas dibuktikan dari eksperimen menggunakan tanaman yang mengekspresikan secara berlebihan gen CAO bahwa biosintesis klorofil b memegang peranan penting dalam mengatur ukuran antena, namun jalur degradatif klorofil b tetap merupakan hal yang menarik untuk diketahui. Selama ini jalur degradatif klorofil hanya diketahui untuk klorofil a saja, karena produk degradatif klorofil a dapat teramati. Alasan mengapa tidak terdapat produk degradatif klorofil b baru dapat diketahui setelah ditemukannya jalur konversi klorofil b menjadi klorofil a (Ito et al. 1993). Sekarang telah diketahui bahwa klorofil b pertama-tama dikonversi menjadi 7-hydroxymethyl-chl dan kemudian direduksi menjadi klorofil a (Ito et al. 1994; Ito et al. 1996; Ito and Tanaka 1996; Scheumann et al. 1996a; 1996b; Scheumann et al, 1998). Tahap pertama dan kedua membutuhkan NADPH dan ferredoxin sebagai tenaga pereduksi (Scheumann et al. 1998). Aktivitas dari tahap pertama meningkat selama proses senescen pada daun (Scheumann et al. 1999). Penelitian yang dilakukan menunjukkan bahwa klorofil a yang dibentuk dari klorofil b dapat diinsersikan kembali ke dalam protein pengikat klorofil, yang berarti bahwa jalur ini juga berfungsi untuk menggunakan kembali klorofil b (Ohtsuka 1997). Mungkin saja konversi klorofil b menjadi klorofil a dan aktivitas CAO dikoordinasi untuk mengatur kesetimbangan antara klorofil a dan klorofil b. Oleh karena itu interkonversi klorofil a dan klorofil b disebut "chlorophyll cycle" dan diperkirakan siklus ini memainkan peranan penting dalam mengatur ukuran antena (Gambar 10) (Tanaka et al. 1998; Oster et al. 2000). Gambar 10. Sebuah model yang diusulkan untuk siklus klorofil. Klorofil a (atas kanan) di dephytilasi menjadi chlorophyllide (Chlide) a (atas kiri) chlorophyllase (E) dan kemudian dikonversi menjadi Chlide b (bawah kiri) melalui 7hydroxymethyl-chlorophyllide (tengah kiri) oleh CAO (A). Chlide b dapat di konversi kembali menjadi Chlide a melalui 7-hydroxymethyl-chlorophyllide oleh Chl b reductase (C) dan 7-hydroxymethyl-chlorophyllide reductase (D). Alternatif lain, Chlide b di phytilasi menjadi klorofil b (bawah kanan) oleh chlorophyll synthase (B) dan kemudian dikonversi menjadi klorofil a melalui 7hydroxymethyl-Chl (tengah kanan) oleh Chl b reduktase (C) and 7hydroxymethyl-chlorophyllide reduktase (D) (Tanaka and Tanaka 2005). Salah satu tantangan kedepan dalam penelitian tentang klorofil b adalah identifikasi protein atau gen yang terlibat dalam konversi klorofil b menjadi klorofil a. Dengan selesainya sekuensing genom Arabidopsis thaliana memungkinkan hal ini dapat dilakukan lebih mudah (The Arabidopsis Genome Initiative 2000). Dengan mengklon gen yang terlibat dalam konversi klorofil b menjadi klorofil a, memungkinkan dilakukannya pengujian hipotesis bahwa degradasi klorofil memicu terjadinya proses senescen (Vincentini et al. 1995; Matile et al. 1996). Jika hipotesis ini benar, memungkinkan ditundanya proses senescen tanaman melalui supresi aktivitas konversi klorofil b menjadi klorofil a dengan ekspresi berlebih antisense gen yang mengkode enzim pereduksi klorofil b. Transformasi yang diperantarai Agrobacterium: Agrobacterium tumefaciens mempunyai peranan penting dalam perkembangan rekayasa genetika tanaman dan penelitian dasar dalam biologi molekular. Hampir 80% tanaman transgenik dihasilkan melalui cara ini. Pada awalnya, orang percaya bahwa hanya tanaman dikotil, gymnosperma dan beberapa spesies monokotil saja yang dapat ditransformasi menggunakan bakteri ini; namun hasil yang dicapai baru-baru ini merubah secara total pandangan ini yang diperlihatkan oleh beberapa spesies “rekalsitran” yang tidak termasuk inang alami seperti monokotil dan fungi ternyata dapat ditransformasi (Chan et al. 1993;Bundock et al. 1995). Sebagai tambahan, sel yang tertransformasi biasanya membawa jumlah kopi T-DNA terintregrasi dalam jumlah rendah didalam genomnya dengan sedikit atau tanpa penantaan kembali, dan DNA berukuran sangat besar dapat ditransformasikan ke dalam tanaman. Dasar molekular transformasi genetik sel tanaman oleh Agrobacterium tumefaciens adalah transfer dari bakteri dan integrasi ke dalam genom tanaman suatu bagian dari plasmid Ti (tumor-inducing) atau Ri (rhizogenic-inducing) (Gambar 11). Transfer T-DNA diperantarai oleh produk yang disandikan oleh suatu daerah vir yang berukuran 30-40 kb pada plasmid Ti. Daerah ini terdiri sedikitnya 6 operon esensial (vir A, vir B, vir C, vir D, vir E, vir G) dan dua non esensial (vir F, vir H). Operon yang konstitutif hanya operon vir A dan vir G, yang menyandikan sistem dua komponen (VirA-VirG) yang akan mengaktifkan transkripsi gen-gen vir lainnya. Vir A merupakan suatu protein sensor dimer transmembran yang dapat menditeksi molekul signal, terutama senyawa fenolik kecil yang dilepaskan oleh tanaman yang terluka (Pan et al. 1993). Signal lain bagi aktivasi Vir A diantaranya pH asam, senyawa fenolik seperti acetosyringon (Winans et al. 1992) dan monosakarida jenis tertentu yang bekerja secara sinergis dengan senyawa fenolik (Ankenbauer et al. 1990; Cangelosi et al. 1990; Shimoda et al. 1990a; Doty et al. 1996). Protein VirA secara struktural dapat dibagi menjadi tiga domain: domain periplasma (input) dan dua domain transmembran (TM1 dan TM2). Domain TM1 dan TM2 bertindak sebagai suatu transmiter (signaling) dan penerima (sensor) (Parkinson 1993). Domain periplasma sangat penting untuk deteksi monosakarida (Chang and Winans 1992). TM2 merupakan suatu domain kinase dan mempunyai peranan yang penting dalam aktivasi VirA yaitu memfosforilasi dirinya sendiri pada residu His-474 (Huang et al. 1990; Jin et al. 1990a; 1990b) sebagai respon atas molekul signal dari tanaman yang terluka. Deteksi monosakarida oleh VirA merupakan sistem amplifikasi penting dan respon terhadap jumlah senyawa fenolik yang rendah. Induksi sistem ini hanya mungkin melalui protein pengikat gula (glukosa/galaktosa) periplasma ChvE (Ankenbauer and Nester 1990; Cangelosi et al. 1990), yang berinteraksi dengan VirA (Shimoda et al. 1990b; Chang and Winans 1992; Turk et al. 1993). VirA yang teraktivasi mempunyai kemampuan untuk mentransfer gugus fosfatnya ke residu aspartat dari suatu protein pengikat DNA sitoplasma VirG (Jin et al. 1990a; 1990b; Pan et al., 1993). VirG berfungsi sebagai faktor transkripsional yang mengatur ekspresi gen-gen vir lainnya ketika terfosforilasi oleh VirA (Jin et al. 1990a; 1990b). Daerah Cterminal bertanggung-jawab untuk aktivitas pengikatan DNA sedangkan daerah N-terminal adalah domain fosforilasi dan mempunyai kemiripan dengan domain penerima signal VirA (sensor). Aktivasi sistem vir juga tergantung pada faktor luar seperti suhu dan pH. Pada suhu yang melebihi 32°C, gen-gen vir tidak akan terekspresi karena perubahan konformasi dalam proses pelipatan protein VirA yang menyebabkan protein ini tidak aktif (Jin et al. 1993). VirD1 dan VirD2 bertanggung-jawab untuk pembentukan T-strand, TDNA utas tunggal, dengan mengenali dan memotong secara spesifik utas bagian bawah T-DNA pada kedua pembatas, yang mana batas kanan merupakan titik awal dan sangat penting dalam proses ini. Setelah pemotongan, VirD2 tetap terikat secara kovalen pada ujung 5’ utas T, membentuk suatu komplek dengan karakter polar dimana ujung 5’ akan bertindak sebagai bagian kepala dalam proses transfer. Komplek VirD2/utas T dan protein pengikat DNA utas tunggal lain yang disebut VirE2 dipercaya ditransferkan secara terpisah ke dalam sel tanaman melalui suatu struktur yang mirip pilus yang terdiri atas suatu komplek protein VirB (Fullner 1996) and VirE2 selama didalam sel bakteri tidak dapat menempel ke utas T karena adanya protein chaperonin yang disebut VirE1 (Deng et al. 1999; Sundberg and Ream 1999). Sekali memasuki sel tanaman, molekul VirE2 akan bersama-sama menempel ke utas T, membentuk komplek T (Sundberg et al. 1996), yang kemudian ditargetkan ke dalam nukleus oleh nuclear target signals (NLS) yang terdapat pada VirD2 dan VirE2, dimana TDNA diintegrasikan secara acak ke dalam genom tanaman dengan kopi tunggal atau ganda. Mekanisme yang terlibat dalam integrasi T-DNA belum terkarakterisasi, namun diperkirakan melibatkan proses rekombinasi yang tidak sah (Gheysen et al. 1991; Lehman et al. 1994; Puchta 1998). BAHAN DAN METODE Galur-galur bakteri dan plasmid Bakteri dan plasmid yang digunakan pada penelitian ini tercantum pada Tabel1. Tabel 1. Galur-galur Bakteri dan Plasmid yang Digunakan Galur dan Plasmid Galur Karakteristik Referensi SupE44 ∆lacU169 (Ǿ80 lacZ∆M15) hsdR17 recA1 endA1 gyrA96 thi-1 relA1 Sambrook et al. (1989) pTiBo542, C58, Hood et al. (1993) Apr, promotor Hellens et al. (2000) pGII0029 Kmr, T-DNA yang membawa gen nptII, lacZ Hellens et al. (2000) pSoup Tetr, pSa Rep Hellens et al. (2000) pOK12 KmR, lacZ- Vieira dan Messing (1991) pBBR1MCS2 KmR, lacZ Kovach et al (1994) E. coli DH5α A. tumefaciens EHA105 Plasmid p35S-2 pAS900 R Km , lacZ Suwanto (unpublished) pUI1014 pUC18 yang berukuran 2 kb pOK-hemA pOK12 yang disisipi fragmen hemA yang diklon pada situs HindIII dan EcoRI Penelitian ini pBBR1MCS2-AShemA Fragmen hemA dari pOK-hemA disubklon pada situs EcoRV dan EcoRI Penelitian ini p35S-hemA Plasmid p35S-2 yang disisipi fragmen hemA pada situs HindIII dan EcoRI Penelitian ini p35S-AshemA pGII0029-hemA pGII0029-AshemA pAS900-CAO1 membawa fragmen hemA Fragmen hemA dari pBBR1MCS2-AshemA yang diangkat dengan EcoRV dan SpeI disubklon ke p35S-2 pada situs XbaI dan SmaI pGII0029 yang disisipi hemA dibawah promotor 35 dari p35S-hemA pada situs EcoRV pGII0029 yang disisipi hemA pada posisi antisense dibawah promotor 35S dari p35SAshemA pada situs EcoRV pAS900 yang disisipi CAO pada situ SpeI dan SacI Neidle and Kaplan (1993) Penelitian ini Penelitian ini Penelitian ini Penelitian ini Pembuatan E. coli kompeten E. coli strain DH5α ditumbuhkan dalam botol kultur berisi 25 ml medium LB selama 16 jam dengan pengocokan 150 rpm pada suhu 37°C. Keesokkan harinya sebanyak 250 µl inokulum diinokulasikan ke dalam 25 ml medium LB segar dan diinkubasi pada suhu 37°C dengan pengocokan 150 rpm selama 3 jam atau sampai mencapai OD600 nm = 0.4. Kultur selanjutnya diinkubasikan diatas es selama 30 menit dan sebanyak 3 ml dipanen dalam tabung eppendorf dengan sentrifugasi pada kecepatan 5000 rpm selama 2 menit. Sel kemudian dicuci dengan 1 ml larutan NaCl (0.1M NaCl, 5 mM Tris-Cl, 5 mM MgCl2, pH 7). Sel yang telah dicuci ini kemudian diresuspensi dalam 1 ml larutan CaCl2 (0.1M CaCl2, 5 mM Tris-Cl, 5 mM MgCl2, pH 7) selanjutnya diinkubasi selama 20 menit diatas es. Selasai inkubasi, sel diendapkan dengan sentrifugasi pada kecepatan 5000 rpm 2 menit dan diresuspensikan kembali dalam 200 µl larutan CaCl2 dan diinkubasi kembali selama 1 jam. Transformasi E. coli Sel kompeten yang telah dibuat dengan cara di atas kemudian ditambahkan dengan 10µl hasil ligasi dan diinkubasi selama 45 menit diatas es. Selanjutnya dilakukan kejutan pada suhu 42ºC selama 1 menit dan diinkubasikan kembali diatas es selama 2 menit. Untuk recovery ditambahkan sebanyak 250 µl LB dan diinkubasi selama 1 jam dengan digoyang pada kecepatan 170 rpm. Setelah itu sel hasil transformasi disebar di atas media dengan antibiotik yang sesuai dan diinkubasi selama semalam. tumbuh digores di atas media baru. Keesokan harinya transforman yang Purifikasi DNA dari gel agarose Fragmen DNA yang didapat dari hasil digesti dipisahkan dengan elektroforesis gel agarose dan fragmen yang diinginkan dipotong keluar dari gel menggunakan pisau scalpel bersih. DNA kemudian dipurifikasi menggunakan QiaQuick spin columns (Qiagen, Studio City, CA) mengikuti protocol yang disarankan oleh pembuatnya. Volume bufer elusi yang digunakan adalah 30µl dan sebanyak 1 µl hasil elusi diukur konsentrasinya dengan spektrofotometer. Isolasi DNA plasmid Koloni bakteri yang didapat dari hasil transformasi ditumbuhkan di medium LB dengan antibiotik yang sesuai selama semalam pada suhu 37°C. Sebanyak 3 ml kultur dipelet dalam tabung eppendorf dengan sentrifugasi pada kecepatan 13.000 rpm selama 2 menit. Supernatan kemudian dibuang dan diresuspensikan dalam 200 µl bufer P1 (50 mM Tris-Cl pH 8, 10 mM EDTA pH 8, 10 mg/ml RNAse). Selanjutnya ditambahkan 200µl bufer P2 (0.2 N NaOH, 1% SDS) dan dibolak-balik beberapa kali sampai lisis sempurna. Kemudian ditambahkan 200 µl bufer P3 (3 M Kalium asetat bufer pH 5) dan divortex selama 10 detik. Selanjutnya sentrifugasi pada kecepatan 10.000 rpm selama 10 menit. Supernatan kemudian pindahkan pada tabung baru dan diekstrak dengan 350µl fenol:kloroform:isoamilalkohol (25:24:1) dan divortex selama 20 detik dan disentrifugasi pada kecepatan 10.000 rpm selama 10 menit. Fase air yang terbentuk dipindahkan ke tabung baru dan dipresipitasi dengan 0.7 volume isopropanol dingin dan diinkubasi di -20°C selama 10 menit. Setelah itu disentrifugasi pada kecepatan 10.000 rpm selama 1 menit. Pelet yang terbentuk kemudian dicuci dengan 1 ml 80% etanol dan dikeringkan dengan vakum. DNA kemudian dilarutkan dengan 50 µl dH2O. Sebanyak 5µl DNA digunakan untuk analisis restriksi. Konstruksi vektor ekspresi ALAS Gen hemA diamplifikasi dari plasmid pUI1014 (Neidle and Kaplan 1993) menggunakan primer IrawanHF (5’-CCCAAGCTTATGGACTACAATCTG-3’) IrawanER (5’-ACCGGAATTCTCAGGCAACGACCTC) yang telah diberi tambahan situs restriksi HindIII dan EcoRI menggunakan enzim DyNAzymeTM EXT DNA Polymerase (FinnZymes, Singapore), kemudian diklon ke dalam plasmid pOK12 (Vieira and Messing 1991) untuk menghasilkan plasmid rekombinan pOK-hemA (Gambar 12). Fragmen hemA ini lalu disisipkan dibawah promoter 35S pada plasmid p35S-2 (Hellens et al. 2000) yang telah didigesti dengan enzim HindIII dan EcoRI (Gambar 13). Gen hemA yang telah diletakkan dibawah promoter 35S dengan orientasi sense ini kemudian diklon ke dalam plasmid pGII0029 (Hellens et al. 2000) pada situs EcoRV menghasilkan plasmid rekombinan pGII0029-hemA (Gambar 14). Untuk mengkonstruksi gen hemA dengan orientasi antisense, gen hemA yang telah diklon ke plasmid pOK12 (Vieira and Messing 1991) diangkat lagi menggunakan enzim EcoRI dan EcoRV untuk disubklon ke plasmid pBBR1MCS2 (Kovach et al 1994) menghasilkan plasmid rekombinan pBBR1MCS2-AShemA (Gambar 15). Kemudian fragmen hemA diangkat kembali dengan enzim EcoRV dan SpeI untuk selanjutnya disubklon ke plasmid p35S-2 yang telah didigesti dengan enzim XbaI dan SmaI menghasilkan plasmid rekombinan p35S-AShemA (Gambar 16). Selanjutnya diklon ke plasmid pGII0029 (Hellens et al. 2000) yang telah didigesti dengan enzim yang sama (Gambar 17). lacZ Ap pUI1014 3910 bp EcoRI hemA HindIII lacZ Ori rep pOK12 Ori 2135 bp PCR HindIII EcoRI hemA Km 1.2 kb Digesti dengan HindIII + EcoRI EcoRI Ori rep lacZ pOK-hemA 3335 bp hemA Km lacZ HindIII Gambar 12. Kloning hemA ke plasmid pOK12 menghasilkan rekombinan plasmid pOK-hemA EcoRI lacZ Ori rep pOK-hemA 3335 bp BamHI hemA SmaI SacI XbaI HindIII Km EcoRI EcoRV EcoRV 35S promoter lacZ HindIII p35S-2 CaMV polyA Digesti dengan HindIII + EcoRI EcoRV EcoRV hemA p35S-2hemA 35S promoter CaMV polyA Gambar 13. Konstruksi gen hemA dibawah promoter 35S dengan orientasi sense BspHI BglII HpaI pSa Ori PvuI LB nptI pGII0029 4.7 kb BspHI nos-kan ColEI ori RB BglII StuI EcoRV hemA 35S promoter lacZ p35S- 2hemA EcoRV CaMV polyA Digesti dengan EcoRV EcoRV BglII HpaI BspHI PvuI pSa Ori nptI BspHI LB nos-kan ColEI ori pGII0029-hemA 6.6 kb BglII StuI RB EcoRV polyA 35S EcoRV hemA Gambar 14. Konstruksi binary vektor yang membawa gen hemA dibawah promoter 35S dengan orientasi sense EcoRI lacZ Ori rep Mob Km KpnI XhoI pOK-hemA 3335 bp pBBR1MCS-2 5144 bp SalI HindIII hemA Km EcoRV EcoRI SmaI BamHI SpeI XbaI SacI lacZ Digesti dengan EcoRI + EcoRV HindIII EcoRV MCS Rep Digesti dengan EcoRI + EcoRV Mob Km BamHI SmaI S acI XbaI HindIII EcoRI EcoRV E coRV 35S promoter p35S-2 CaMV polyA KpnI XhoI SalI HindIII EcoRV pBBR1MCS-2AShemA 6.4 kb Rep hemA EcoRI SmaI BamHI SpeI XbaI SacI Digesti dengan XbaI + SmaI EcoRV 35S promoter Digesti dengan EcoRV + SpeI hemA p35S-2AShemA EcoRV CaMV polyA Gambar 15. Konstruksi gen hemA dibawah promoter 35S dengan orientasi antisense BspHI BglII HpaI pSa Ori PvuI LB nptI pGII0029 4.7 kb BspHI nos-kan hemA EcoRV ColEI ori RB BglII StuI 35S promoter lacZ EcoRV p35S-2AShemA EcoRV CaMV polyA Digesti dengan EcoRV BglII HpaI BspHI PvuI pSa Ori nptI BspHI LB nos-kan ColEI ori pGII0029-AShemA 6.6 kb BglII StuI RB EcoRV polyA 35S EcoRV hemA Gambar 16. Konstruksi binary vektor yang membawa gen hemA dibawah promoter 35S dengan orientasi antisense Gambar 17. Peta plasmid pSOUP Transformasi plasmid rekombinan ke Agrobacterium tumefaciens EHA 105 Transformasi plasmid dilakukan menggunakan metode freeze-thaw (An et al. 1988) ke dalam Agrobacterium tumefaciens EHA 105 (Hood et al. 1993). Plasmid rekombinan ditransformasi secara bersamaan dengan plasmid pSOUP (Gambar 17) (Hellens et al. 2000). Agrobacterium thumefaciens EHA105 (pGII0029-hemA/AShemA) ditumbuhkan selama semalam pada suhu 28°C dalam 25 ml medium TYNG (10g/l Tripton, 5g/l ekstrak khamir, 5g/l NaCl, 1g/l glukosa, 0.2 g/l MgSO4, pH 7.5). Keesokkan harinya dilakukan subkultur sebanyak 1% ke medium yang baru dan diinkubasikan pada kondisi yang sama selama semalam. Selanjutnya kultur didiamkan diatas es selama 10 menit kemudian di panen dengan disentrifugasi pada kecepatan 5000 rpm 4°C selama 10 menit. Setelah supernatant dibuang, sel dicuci dengan 1 ml 20mM CaCl dingin dan disentrifugasi kembali dengan kondisi yang sama. Sel kembali diresuspensi dalam 1 ml 20 mM CaCl2 dingin. Kedalam 150 µl sel Agrobacterium ditambahkan sebanyak 1µg plasmid dan dicampur rata kemudian dibekukan dengan nitrogen cair selama 5 menit. Setelah itu dicairkan kembali dengan meletakkannya diatas meja pada suhu ruang dan ditambahkan 1 ml LB (10g/l Tripton, 5g/l ekstrak khamir, 5g/l NaCl) untuk kemudian diinkubasi selama semalam pada suhu 28°C dengan kecepatan 200 rpm. Setelah kurang lebih 16 jam, kultur kemudian disebar diatas medium TYNG yang diberi antibiotic kanamisin 50µg/ml dan tetrasiklin 2µg/ml diinkubasikan kembali sampai ada koloni yang tumbuh (sekitar 3-4 hari) Transformasi rekombinan binary vector ke Arabidopsis thaliana Transformasi dilakukan dengan mencelupkan bunga tanaman Arabidopsis thaliana ecotype Columbia ke dalam suspensi Agrobacterium tumefaciens EHA105 (pGII0029-hemA) (Floral dip transformation) (Clough and Bent 1998) sedangkan seleksi transforman dilakukan secara in solium (Xiang et al. 1999). Arabidopsis thaliana ditumbuhkan sampai fase pembungaan di dalam walk-in incubator dengan suhu 22°C dan penyinaran ~ 80 µE.m-2.s-1 selama 24 jam. Tanaman ditanam pada tabung PVC yang berdiameter 4.5 cm dengan tanah (Trubus) yang dipasteurisasi. Untuk mencegah terjatuhnya tanah pada saat infeksi bakteri, tabung ditutupi dengan kain kassa yang diikat dengan karet gelang (Gambar 18a). Tanaman ditumbuhkan selama 32-34 hari atau sampai tangkai bunga mencapai panjang 2-9 cm dengan beberapa bunga yang telah mekar (Gambar 18b). Agrobacterium tumefaciens strain EHA105 (pGII029hemA) ditumbuhkan selama semalam (~ 18 jam) pada medium cair YEP (10 g/l yeast extract, 10 g/l peptone, 5 g/l NaCl) yang diberi antibiotik kanamisin (50 mg/l) dan diinkubasi pada suhu 28°C dengan kecepatan 180 rpm. Sel bakteri kemudian dipanen dengan sentrifugasi pada kecepatan 6000 g pada suhu 4°C selama 10 menit dan diresuspensikan pada medium infeksi (5% sukrosa + 0.05% Agristick® (Bayer CropScience, Jakarta) sehingga mencapai OD600 sekitar 0.80 (Clough and Bent 1998). A B Gambar 18. Tanaman Arabidopsis thaliana yang ditumbuhkan pada tabung PVC (A) dan siap diinfeksi (B) Tanaman dicelupkan kedalam suspensi bakteri selama 3-5 detik dengan digoyang secara perlahan (Gambar 19a) dan selanjutnya diberi selubung dengan ujung yang ditutup untuk menjaga kelembaban dan mencegah perkawinan silang (Gambar 19b). Tanaman yang telah diinfeksi diinkubasi di ruang gelap selama semalam sebelum dikembalikan ke kondisi lingkungan yang telah disiapkan. Plastik penutup pada ujung selubung dibuka setelah 24 jam pasca infeksi. Tanaman ditumbuhkan sampai menghasilkan biji untuk selanjutnya dilakukan seleksi untuk mendapatkan kandidat tanaman yang tertransformasi. A B Gambar 19. Tahapan infeksi dengan suspensi Agrobacterium (A) dan pasca infeksi (B) Seleksi kandidat tanaman transgenik dilakukan secara In-solium mengikuti metode yang dikembangkan oleh Xiang et al. (1999). Biji yang telah dikeringkan dikecambahkan secara langsung ditanah (1,000-3,000 biji pada wadah plastik berdiameter 18). Setelah melewati tahap stratifikasi selama 3 hari pada suhu 4°C, wadah dipindahkan ke walk-in incubator dengan penyinaran ~ 80 µE.m-2.s-1 selama 24 jam dan suhu 22°C. Setelah kecambah tumbuh dan dua daun pertamanya telah muncul, dilakukan penyemprotan dengan kanamisin yang telah dicampur dengan 0.1% Agristick® (Bayer CropScience, Jakarta). Penyemprotan kanamisin dilakukan setiap hari dengan jumlah yang cukup untuk membasahi seluruh permukaan daun. Tanaman disemprot dengan 100 mg/l kanamisin selama 2 hari, dilanjutkan dengan 200 mg/l kanamisin selama 2 hari, dan akhirnya dengan 500 mg/l kanamisin (Xiang et al. 1999). Selama tahap seleksi kecambah ditutupi dengan plastic untuk mencegah efek terbakarnya daun dan dehidrasi yang berlebihan akibat kombinasi perlakuan dengan antibiotik dan surfaktan. Analisis kandidat tanaman transgenik dengan PCR Kandidat tanaman transgenik diisolasi total DNA genomnya menggunakan metode dari Lassner et al. (1989): Sebanyak 0.2 g bahan tanaman segar digerus dengan menggunakan nitrogen cair dan dimasukkan ke tabung 1.5 ml. Sebanyak 0.5 ml bufer isolasi (2% (w/v) CTAB, 1.4 M NaCl, 20 mM EDTA, 100 mM TrisHCl, pH 8.0, 0.2% ß-merkaptoetanol) ditambahkan kedalam tabung dan dicampur dengan cara dibolak-balik sampai homogen. Selanjutnya dilakukan inkubasi pada suhu 65˚C selama satu jam dengan setiap 10 menit dibolak-balik agar homogen. Sebanyak satu kali volume kloroform-isoamil alkohol (24:1 v/v) ditambahkan sample dengan dibolak-balik sampai homogen selama 10 menit. Selanjutnya dilakukan sentrifugasi pada kecepatan 8000 rpm selama 15 menit. Fase cair yang terbentuk kemudian dipindahkan ke tabung baru dan ditambahkan 1 µl RNase A (10 mg/ml dalam 10 mM Tris-HCl, 15 mM NaCl, pH 7.5) dan diinkubasi pada suhu 37ºC selama 1 jam. Presipitasi DNA dilakukan dengan penambahan 1/30 volume 3 M sodium asetat (pH 5.2) dan 0.6 volume isopropanol dingin dan dicampur dengan membolak-balikkan tabung beberapa kali. Pelet DNA didapat setelah dilakukan sentrifugasi pada kecepatan 6.500 rpm selama 10 menit. Setelah supernatan dibuang, pelet dicuci dengan penambahan 0.5 ml 80% ethanol dan disentrifus kembali dengan kecepatan 8.000 rpm selama 3 menit di suhu ruang. Pelet selanjutnya dikering-udarakan dan dilarutkan dibufer TE. DNA kandidat tanaman transgenik kemudian di PCR menggunakan GeneAmp PCR System 2400 (Perkin Elmer) untuk mengetahui keberadaan gen hemA dan kanamisin menggunakan primer NewhemAF (CTACAATCTGGCACTCGATA C) dan NewhemAR (GTCCGAGATCATCTTGCAGT) dan kanF (GTT CTTTTT GTCAAGACCGACCT) dan kanR (GCTCAGAAGAACTCGTCAAGAAG) dengan kondisi PCR (sama untuk keduanya) : 94°C, 30”; 50°C, 1’; 72°C, 1’ sebanyak 25 siklus. . Analisis transkrip dengan Reverse Transcriptase (RT)-PCR Analisa transkrip dilakukan dengan mengisolasi total RNA dan pembuatan cDNA menggunakan Trizol® Reagent (Invitrogen, California) dan ProtoScriptTM First strand cDNA Synthesis Kit (New England Biolab, Singapore). Sebanyak 100 mg sample daun dihancurkan 1 ml TRIzol Reagent menggunakan batang pengerus sampai homogen. Material tanaman yang tidak larut dipisahkan dengan sentrifugasi 12.000x g selama 10 menit pada suhu 4°C. Homogenat kemudian dipindahkan ke tabung yang baru dan diinkubasi selama 4 menit pada suhu ruang dan selanjutnya ditambahkan 0.2 ml kloroform. Tabung dikocok dengan tangan selama 15 menit dan diinkubasi kembali selama 2-3 menit pada suhu ruang. Setelah itu dilakukan sentrifugasi pada kecepatan 12.000 x g selama 15 menit pada suhu 4°C. Fase cair yang dihasilkan kemudian dipindahkan ke tabung yang baru dan RNA dipresipitasi dengan penambahan 0.5 ml isopropanol. Inkubasi pada suhu ruang selama 10 menit dan disentrifugasi kembali pada kecepatan 12.000 x g selama 10 menit pada suhu 4°C. Pelet RNA kemudian dicuci dengan penambahan 1 ml etanol 75% dan divortex dan kemudian disentrifus kembali pada kecepatan 7.500 x g selama 5 menit pada suhu 4°C. Pelet RNA kemudian dikering udarakan dan selanjutnnya dilarutkan dengan RNase-free water. Sebanyak 5µl ekstrak RNA ditambahkan dengan 2 µl primer d(T)23 VN (50 µM) dan 4 µl dNTP (10 mM) serta air bebas nuklease sampai mencapai volume 16 µl. Kemudian dipanaskan pada suhu 70°C selama 5 menit dan disentrifugasi secara cepat dan diletakkan diatas es. Selanjutnya ditambahkan 2 µl 10X bufer RT, 1 µl inhibitor RNase dan 1 µl M-MuLV Reverse Transcriptase(New England Biolab, Singapore) untuk kemudian diinkubasi pada suhu 42°C selama satu jam. Inaktivasi enzim dilakukan dengan pemanasan pada suhu 95°C selama 5 menit. Untuk menghilangkan RNA ditambahkan 1µl RNase H (2 unit) dan diinkubasi pada 37°C selama 20 menit dan diinaktivasi kembali dengan pemanasan pada 95°C selama 5 menit. Hasilnya kemudian diencerkan menjadi 50µl dengan dH2O untuk selanjutnya sebanyak 5 µl digunakan untuk reaksi PCR. Pengukuran kandungan klorofil Kandungan klorofil diukur menggunakan metode Arnon (1959). Potongan daun segar digerus dalam nitrogen cair kemudian ditambahkan aseton 80% dan diaduk sampai homogen. Ekstrak klorofil disaring dengan kertas saring Whatman no 42 dan dimasukkan ke dalam labu ukur 100 ml. Sisa jaringan diekstrak sekali lagi dengan aseton 80% dan hasilnya dicampur dengan ekstrak yang pertama kemudian ditambahkan aseton sampai mencapai volume 100 ml. Sebanyak 5 ml larutan dimasukkan ke dalam labu ukur 50 ml dan diencerkan dengan aseton 80% sampai mencapai volume 50 ml. Ekstrak klorofil selanjutnya diukur pada panjang gelombang 663, 645 dan 652 nm. Jumlah tanaman yang digunakan untuk analisis klorofil adalah 3 untuk tanaman kontrol dan 8 untuk tanaman trangenik. Esei aktivitas ALA sintase dan total kandungan ALA Sebanyak satu gram potongan daun digerus dalam nitrogen cair kemudian ditambahkan 1 ml bufer (50 mM Tris-Cl pH 7.6, 5 mM EDTA pH 8, 0.2% ßmercaptoethanol). Sebanyak 100 µl ekstrak dicampurkan dengan 100µl bufer esei yang mengandung : Tris-Cl pH 7.6 (100 mM); Glisin (200 mM), Piridoksal fosfat (0.2 mM); Suksinil-CoA (0.32 mM); DTT (2 mM) dan Asam levulinat (20 mM). Campuran esei ini kemudian diinkubasi selama 4 jam pada suhu 37°C. Setelah diinkubasi dilakukan pengukuran kandungan ALA menggunakan reagen Ehrlich (Urata dan Granick 1963) sebagai berikut: sebanyak 100 µl 10% TCA ditambahkan kedalam campuran esei kemudian disentrifugasi pada kecepatan 13.000 rpm selama 5 menit. Sebanyak 300 µl supernatant ditambahkan dengan 400µl 1 M Sodium asetat pH 4.6 dan 35 µl asetilaseton dan selanjutnya dididihkan selama 20 menit. Setelah didinginkan sampai suhu ruang, sebanyak 700 µl reagen Ehrlich ditambahkan dan didiamkan selama 20 menit sebelum diukur absorbansinya pada panjang gelombang 556. Pengukuran berat basah dan berat kering Arabidopsis thaliana ditumbuhkan sampai berumur 6 minggu kemudian ditimbang berat basahnya. Berat kering ditimbang setelah dikeringkan dalam oven selama 2 hari pada suhu 60°C Pengujian ketahanan terhadap salinitas Tanaman Arabidopsis yang berumur 3 minggu didalam pot yang berisi tanah disiram dengan larutan yang mengandung 0, 200 mM dan 400 mM NaCl setiap hari selama 2 minggu. Setelah 2 minggu dilakukan dokumentasi terhadap efek perlakuan NaCl. Kloning gen Chlorophyll A Oxygenase dari Arabidopsis thaliana Kloning gen Chlorophyll A Oxygenase dari Arabidopsis thaliana dilakukan dengan menggunakan teknik PCR berdasarkan sekuen yang ada di GeneBank dengan accession number NM-103548. Total RNA diekstrak menggunakan Trizol® Reagent (Invitrogen, California) dan cDNA disintensis menggunakan ProtoScriptTM First strand cDNA Synthesis Kit (New England Biolab, Singapore). Setelah itu dilakukan amplifikasi menggunakan primer CAOF (5’-TTTCCGGACTAGTCATGAACGC-3’) dan CAOR (5’-AGATTTCTTTGAGCTCAGTTAGCC-3’) yang membawa situs restriksi bagi enzim SpeI dan SacI menggunakan enzim DeepVentR Polymerase (New England Biolab, Singapore) dengan kondisi PCR : 94°C, 30”; 50°C, 1’; 72°C, 1’ . Hasil amplifikasi kemudian elektroforesis dan pita DNA berukuran sekitar 1.6 kb dipotong dan dipurifikasi menggunakan QiaQuick spin columns (Qiagen, Studio City, CA). Fragmen yang diduga gen CAO ini kemudian didigesti dengan enzim SpeI + SacI dan sisipkan ke plasmid pAS900 pada situs restriksi yang sama. Fragmen yang telah menyisip diplasmid pAS900 (Suwanto, unpublished) ini kemudian ditentukan urutan nukleotidanya menggunakan automatic DNA Sequencer ABI3100 untuk menentukan apakah memang benar gen ini adalah gen Chlorophyll A Oxygenase dan tidak mengalami mutasi selama proses PCR. Tempat dan waktu penelitian Penelitian ini dilakukan di laboratorium Biologi Molekuler SEAMEOBIOTROP, Bogor, Scottish Agricultural College, Scotland dan Research Center for Microbial Diversity IPB, Bogor dari bulan Januari 2002 sampai Desember 2004. HASIL DAN PEMBAHASAN Konstruksi vektor ekspresi ALAS Pada penelitian ini telah dilakukan konstruksi rekombinan binary vector yang membawa gen hemA dibawah promoter 35S baik dengan orientasi sense maupun antisense. Seri plasmid pGreen (Hellens et al. 2000) dipakai dalam penelitian ini karena ukurannya yang kecil sehingga memudahkan dalam proses kloning, fleksibilitas dalam pemilihan penanda seleksi dan banyaknya situs restriksi untuk kloning (MCS) serta jumlah kopi yang banyak. Plasmid binary vektor yang berukuran kecil sangat berguna jika DNA yang ingin kita masukkan ke dalam sel tanaman berukuran besar. Selama ini binary vector yang ada berukuran lebih dari 10 kb. Seri plasmid pGreen hanya bisa bereplikasi di dalam sel Agrobacterium jika berada bersama plasmid pSOUP yang membawa gen pSa replicase yang berkerja in trans terhadap pSa ori yang terdapat pada pGreen. Analisis restriksi atas plasmid rekombinan yang dihasilkan dalam proses konstruksi vektor ekspresi asam δ-aminolevulinat sintase (ALAS) dapat dilihat pada Gambar 20 dan 21. Peta plasmid baik yang membawa gen hemA dengan orientasi sense maupun antisense terhadap promoter 35S dapat dilihat pada Gambar 22. Karena plasmid pGreen tidak dapat dimobilisasi dengan cara konjugasi, introduksinya ke dalam Agrobacterium dilakukan dengan metode freeze-thaw (An et al. 1988) bersama-sama dengan plasmid pSOUP. Bila diinginkan untuk membuat tanaman transgenik yang marker-free, kedua plasmid ini bisa dipakai untuk membawa T-DNA yang terpisah antara gen yang diinginkan dengan marker antibiotik untuk seleksi transforman. M 1 2 3 4 5 6 7 8 9 10 3.0 kb 1.5 kb 1.0 kb Gambar 20. Hasil analisis restriksi plasmid rekombinan pOK12-hemA, p35S2hemA dan pGII0029-hemA. M: molekular marker, 1: fragmen hemA berukuran ± 1.2 kb, 2: pOK12, 3: pOK12-hemA didigesti dengan HindIII + EcoRI, 4: pOK12-hemA didigesti dengan HindIII + EcoRI+BamHI, 5:p35S-2, 6: p35ShemA yang didigesti dengan HindIII+EcoRI, 7: p35S-hemA yang didigesti dengan HindIII+EcoRI+BamHI, 8:p35S-hemA yang didigesti dengan EcoRV, 9:pGII0029, 10:pGII0029-hemA yang didigesti dengan EcoRV M 1 2 3 M 5 6 6.0 3.0 2.0 1.5 1.0 0.5 A B Gambar 21. Hasil analisis restriksi plasmid rekombinan pGII0029-hemA (A) dan pGII0029-AShemA (B) yang didigesti dengan BamHI (3 & 5), EcoRI (1 & 6) dan HindIII (2 & 7) Gambar 22. Peta plasmid rekombinan pGII0029-hemA dan pGII0029-AShemA Transformasi rekombinan binary plasmid ke Arabidopsis thaliana Introduksi gen penyandi ALAS dilakukan pada tanaman Arabidopsis thaliana karena tanaman ini relatif cepat pertumbuhan dan siklus hidupnya pendek serta cara transformasi melalui infeksi Agrobacterium pada bagian bunga (Floral dip transformation) yang langsung dapat menghasilkan biji transgenik (Clough and Bent 1998). Dengan cara ini tidak diperlukan proses kultur jaringan karena benih yang dihasilkan dapat langsung diseleksi pada saat ditumbuhkan di tanah dan disemprot dengan antibiotik kanamisin (in solium selection) (Gambar 23). Dengan tersedianya cara transformasi yang sederhana dan cepat ditambah dengan selesainya sekuen total genom Arabidopsis thaliana pada tahun 2000, membuat studi tentang ekspresi suatu gen di Arabidopsis thaliana menjadi sangat mengguntungkan. Berkembangnya teknologi microarray memungkinkan untuk mempelajari ekspresi suatu gen dan pengaruhnya terhadap gen-gen lain secara mendalam. Pada penelitian ini, binary vector yang membawa antisense hemA tidak jadi digunakan sebagai kontrol insersi karena faktor insersi T-DNA yang acak tidak mungkin mendapatkan insersi pada tempat yang sama antara konstruk sense dan antisense. A B C D Gambar 23. Kecambah Arabidopsis yang akan (A) dan setelah (B, C dan D) diseleksi dengan penyemprotan antibiotik kanamisin (in solium selection) Analisis tanaman Arabidopsis thaliana transgenik Dalam penelitian ini, berhasil didapatkan beberapa kandidat tanaman transgenik hasil seleksi secara in solium (Gambar 23D) dan berdasarkan analisis dengan PCR, dari lima tanaman, empat diantaranya positif membawa gen hemA dan kanamisin (Gambar 24). Satu tanaman tidak membawa kedua gen tersebut yang diduga non-transforman yang lolos dari seleksi (escape). Seleksi transforman secara in solium ini, menghendaki adanya keseragaman perkecambahan sehingga semua tanaman mengalami seleksi pada waktu yang sama. Penyemprotan kanamisin pada kecambah Arabidopsis thaliana yang memiliki satu pasang daun sangat efektif untuk menyeleksi tanaman yang nontransforman. Bila ada kecambah Arabidopsis thaliana yang baru muncul kemudian (terlihat dari jumlah daunnya yang lebih sedikit dari yang lain) biasanya 7-10 hari setelah penyemprotan kanamisin terakhir, kemungkinan bukan transforman (escape). M 1 2 3 4 5 6 7 hemA Kanr Gambar 24. Analisis kandidat tanaman transgenik menggunakan PCR dengan primer spesifik gen hemA dan kanamisin resisten. M: marker; 1-5: kandidat tanaman transgenik; 6: non-transforman; 7: kontrol Bukti bahwa gen yang diintroduksikan dapat terekspresi dengan baik pada tanaman Arabidopsis thaliana transgenik adalah dari hasil analisis transkrip menggunakan Reverse Transcriptase (RT)-PCR (Gambar 25). Pada analisis transkrip, tanaman Arabidopsis thaliana non-transforman tidak dijumpai adanya produk amplifikasi yang berarti secara alami Arabidopsis thaliana tidak mempunyai gen ini. Tanaman menggunakan jalur biosintesis C-5 (dari glutamat) (Beale et al. 1975) untuk menghasilkan prekursor senyawa tetrapirol sedangkan gen hemA dari Rhodobacter sphaeroides merupakan enzim biosintesis asam δaminolevulinat (ALA) jalur C-4 (kondensasi suksinil-CoA dan glisin) (Gibson et al. 1958). Amplifikasi gen hemA pada preparasi total RNA juga tidak didapati adanya produk amplifikasi, sehingga memastikan bahwa amplifikasi produk dengan primer spesifik hemA pada preparasi cDNA tanaman Arabidopsis thaliana transgenik merupakan hasil dari ekspresi gen hemA yang diintroduksikan. Hasil analisis transkrip ini diketahui bahwa keempat tanaman Arabidopsis thaliana transgenik dapat mengekspresikan gen hemA yang diintroduksikan. Untuk membuktikan bahwa ekspresi gen juga terjadi pada taraf translasi, dilakukan analisis aktivitas enzim ALA sintase. Pengujian aktivitas enzim merupakan cara langsung untuk mengetahui suatu gen terekspresi atau tidak. Karena analisis western blotting untuk mengetahui ada tidaknya ekspresi gen pada taraf protein tidak menjamin bahwa enzim itu aktif karena bisa saja protein enzim ada tapi tidak aktif karena ada masalah pelipatan protein. Tanaman Arabidopsis thaliana transgenik no 4 yang diesei aktivitas enzimnya mempunyai aktivitas ALAS sebesar 40.5 nmol.mg-1.h-1 (tabel 2). Dari keempat tanaman yang positif membawa gen hemA, hanya tanaman No. 4 yang sangat berbeda dari segi morfologinya yaitu tanaman tumbuh lebih besar (Gambar 26) akan tetapi mengalami keterlambatan dalam pembungaan (sekitar satu minggu) dan penurunan jumlah biji yang dihasilkan (± 25% dari tipe liar). Terjadinya perubahan morfologi tanaman menjadi lebih besar dan keterlambatan pembungaan, mungkin disebabkan oleh meningkatnya efisiensi pemanfaatan nitrogen oleh tanaman. Karena tanaman yang diberi pemupukan nitrogen yang berlebihan juga memperlihatkan hal yang sama yaitu tanaman menjadi tumbuh subur (lebih banyak daun) tetapi pembungaan menjadi terlambat. Hotta et al. (1997a) melaporkan bahwa tanaman yang disemprot dengan larutan ALA mengalami peningkatan laju fotosintesis, peningkatan fiksasi CO2 dan penurunan respirasi serta meningkatnya penyerapan hara khususnya N akibat peningkatan aktivitas nitrat reduktase. Yoshida et al. (2004) juga melaporkan adanya peningkatan pengambilan nitrogen dan rasio antara nitrogen dalam bentuk nitrat per total nitrogen pada daun dan petiol pada tanaman Komatsuna. Walaupun penelitian di atas berasal dari aplikasi ALA secara eksogenous, namun kemungkinan hal yang sama terjadi pada tanaman Arabidopsis thaliana transgenik yang membawa gen hemA. Karena gen hemA merupakan penyandi enzim ALAS, ekspresi gen ini menyebabkan terjadinya peningkatan produksi ALA didalam sel (tabel 2). M 1 1.0 kb 0.5 kb 2 3 4 5 6 7 8 9 10 11 hemA 18SR Gambar 25. Hasil analisis RT-PCR pada tanaman transgenik T1. M: molekular marker, 1-4: Tanaman transgenik, 5: Tanaman kontrol, 6: hemA, 7: Kontrol negatif PCR, 8: Internal kontrol 18SRNA, 9-11: kontrol cDNA Gambar 26. Profil tanaman transgenik no 4 (tanda panah). Tanaman mendapatkan nitrogen dari tanah biasanya dalam bentuk nitrat Enzim yang terlibat dalam pemanfaatan nitrat adalah nitrat reduktase dan nitrit reduktase. Nitrat reduktase akan mereduksi nitrat menjadi nitrit dan oleh enzim nitrit reduktase akan direduksi menjadi ammonium. Nitrat dan nitrit reduktase merupakan enzim yang mengandung heme (Vance 1990). Oleh karena heme seperti juga klorofil dibentuk dari ALA, maka dapat dimengerti mengapa peningkatan sintesis ALA dapat meningkatkan aktivitas kedua enzim diatas. Terjadinya penurunan jumlah biji yang dihasilkan kemungkinan efek dari ekspresi ALAS di bawah promoter kuat secara konstitutif. Pada saat pembentukan biji biasanya akan terjadi peningkatan sintesis DNA dan protein yang terlibat dalam pembentukan biji. Pada saat ini aktivitas di organ vegetatif menjadi rendah bahkan sebagian besar N yang ada di biji berasal dari re-alokasi N yang terdapat pada organ vegetatif. Sedangkan 75% C yang ada di biji berasal dari fotosintat. (Peoples and Gifford 1990). Bila aktivitas sel di organ vegetatif masih tetap tinggi seperti dalam hal ini ekspresi ALAS secara berlebihan akan terjadi persaingan pemakaian sumber daya di tanaman yang berakibat biji kekurangan bahan baku yang diperlukan untuk perkembangannya. Selain itu mungkin ALA bersifat organ spesifik di mana pada organ generatif dapat menyebabkan proses fertilisasi menjadi terganggu sehingga berakibat menurunnya jumlah biji yang dihasilkan. Yoshida et al. (2004) melaporkan adanya hubungan erat fungsi fisiologis ALA dengan biosintesis H2O2. Aplikasi ALA tidak hanya menaikkan berat basah dan berat kering tanaman tetapi juga jumlah H2O2 yang ada dijaringan tanaman. H2O2 merupakan senyawa yang berperanan dalam proses programmed cell death (PCD). Hotta et al (1997b) melaporkan bahwa tanaman kacang merah yang diberi perlakuan ALA pada fase awal seperti fase daun primer atau daun pertama, meningkat hasilnya sampai 2030%, sedangkan jika perlakuan diberikan pada fase menurunkan hasil sekitar 10%. pembungaan malah Tabel 2. Hasil analisis berat basah dan berat kering Arabidopsis thaliana transgenik dan non transgenik Berat Basah (gr) Berat Kering (gr) Kontrol 0.95 (100%) 0.18 (100%) Transgenik 1 0.92 (96.8%) 0.21 (116.6%) Transgenik 2 1.05 (110%) 0.18 (100%) Transgenik 3 0.57 (60%) 0.10 (55.5%) Transgenik 4 1.11 (116.8%) 0.19 (105.5%) Hasil analisis berat kering dan berat basah memperlihatkan sejumlah data yang menarik (Table 2). Tanaman transgenik no 4 mempunyai berat basah tertinggi dan mengalami kenaikan 16.8% dibandingkan kontrol tetapi berat keringnya hanya naik 5.5%. Hal serupa terjadi pada tanaman transgenik no 2. Sedangkan tanaman transgenik no 1 yang berat basahnya turun 3.2% dibandingkan kontrol justru mengalami kenaikan berat kering tertinggi 16.6%. Kalau hal ini dihubungkan dengan analisis yang telah diuraikan sebelumnya, yaitu meningkatkanya aktivitas enzim nitrat reduktase, meningkatnya berat basah mungkin akibat meningkatnya efisiensi pemakaian nitrogen oleh tanaman. Proses asimilasi N anorganik dapat dianggap sebagai bagian dari proses fotosintesis. Asimilasi nitrat (NO3-) atau NH4+ membutuhkan tenaga pereduksi dan ATP yang mana keduanya disediakan oleh reaksi terang dari fotosintesis. Kerangka karbon yang dibutuhkan untuk asimilasi N anorganik didapat dari oksidasi senyawa karbon tereduksi yang juga dihasilkan dari proses fotosintesis. Oleh karena itu, meningkatnya asimilasi N akan menguras hasil fiksasi CO2 selama proses fotosintesis yang berakibat pada rendahnya pertambahan berat kering tanaman. Pada tanaman transgenik no 1, karena berat basahnya lebih rendah dari kontrol mungkin disebabkan oleh kemampuannya untuk asimilasi N tidak sebaik tanaman transgenik no 2 dan 4 sehingga tersedia tenaga pereduksi (NADPH) yang cukup untuk mereduksi CO2 yang terlihat dengan terjadinya kenaikan berat kering dibandingkan dengan kontrol. Tanaman transgenik no 3 mengalami penurunan berat basah maupun berat kering yang cukup besar. Hal ini mungkin terjadi karena adanya kelainan perakaran (Gambar 27), dimana tanaman ini buluh akarnya tidak berkembang seperti pada tanaman kontrol maupun transgenik lainnya. Kelainan perakaran ini menyebabkan penyerapan haranya menjadi menurun dan berakibat pada rendahnya pertumbuhan tanaman. Mungkin insersi T-DNA, bukan akibat ekspresi hemA karena jumlah mRNAnya mirip dengan tanaman transgenik no 1, menyebabkan mutasi pada gen perakaran sehingga akar tidak berkembangan dengan baik. A B Gambar 27. Kondisi perakaran tanaman transgenik no 4 (A) dan 3 (B) Ekspresi ALAS meningkatkan kandungan klorofil Tabel 3. Hasil analisis kandungan klorofil, aktivitas ALAS dan ALA Kontrol Trangenik Chl-a (mg/g) Chl-b (mg/g) Total Chl (mg/g) ALA-S (nmol.mg-1.h-1) ALA (µM) 3.577 (100%) 5.313 (148.5%) 1.369 (100%) 1.797 (131.2%) 4.946 (100%) 7.11 (143.7%) 0 0.535 40.5 1.357 Hasil analisis kandungan klorofil terhadap tanaman transgenik no 4 didapatkan bahwa terjadi peningkatan kandungan total klorofil dari 4.9 mg/g berat basah pada tanaman tipe liar menjadi 7.1 mg/g berat basah atau terjadi kenaikan sekitar 43%. Klorofil a dan b mengalami kenaikan sekitar 48% dan 31% (Tabel 3). Peningkatan kandungan klorofil sejalan dengan meningkatnya sintesis ALA yang mungkin sebagian dikonversi menjadi klorofil. Zavgorodnyaya et al. (1997) yang mengekspresikan ALAS khamir ditanaman tembakau dan ditargetkan ke dalam kloroplas mendapatkan bahwa ekspresi ALAS di tanaman yang sebelumnya telah membawa antisense terhadap GSA-AT (salah satu enzim pada jalur C-5) dapat mengembalikan fenotip seperti pada tipe liarnya. Ekspresi ALAS juga dapat meningkatkan kapasitas sintesis ALA (30% dibanding kontrol) dan klorofil (20% dibanding kontrol) serta membuat tanaman menjadi tahan terhadap gabaculin (suatu inhibitor kuat terhadap enzim transaminase yang tergantung vit B, termasuk GSA-AT). Aktivitas ALAS yang tertinggi mencapat 23.05 nmol.mg1 .h-1. Jika kemampuan untuk mensintesis klorofil baru dapat ditingkatkan dengan menyediakan prekursor klorofil yang lebih banyak mungkin dapat menunda terjadinya senescen. Karena pada proses senescen terjadi penghancuran klorofil (Matile et al. 1996). Proses senescen dapat menurunkan laju fotosintesis yang pada akhirnya akan menurunkan produktifitas tanaman pertanian. Analisis penurunan sifat monogenik Mendel Untuk mengetahui pola penurunan sifat dari tanaman transgenik dilakukan pendeteksian keberadaan gen hemA pada 21 tanaman T1 dengan PCR. Ternyata dari 21 tanaman T1, 16 diantaranya membawa gen hemA (76%) atau dengan perbandingan 3:1, sehingga dapat disimpulkan bahwa pola pewarisannya normal sesuai dengan penurunan sifat monogenik Mendel (Gambar 28). M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 K+ kb HemA 1.0 0.75 Kanr Gambar 28. Hasil analisis PCR turunan pertama tanaman transgenik No. 4 Pengujian ketahanan terhadap salinitas Pada tanaman Arabidopsis thaliana transgenik yang mengekspresikan ALAS terjadi peningkatan ketahanan terhadap cekaman salinitas. Hal ini ditunjukkan dengan kemampuannya bertahan hidup selama penyiraman dengan larutan NaCl sampai 200 mM setiap hari selama 2 minggu (Gambar 29). Penelitian yang dilakukan oleh Watanabe et al. (2000) pada kecambah kapas juga menunjukkan hal serupa dimana kecambah kapas dapat tumbuh pada tanah yang diberi perlakuan 1.5% NaCl dan ALA. Hasil analisis kandungan Na+ dijaringan akar tanaman menunjukkan jumlah yang lebih rendah dari tanaman kontrol. Oleh karena itu mereka menduga kehadiran ALA dapat mengurangi pengambilan Na+. Mungkin juga karena ALA meningkatkan pengeluaran Na+ dari sel melalui pengaktifan Na+/H+ antiporter sehingga Na+ di jaringan akan lebih rendah. Nishihara et al. (2003) melaporkan bahwa tanaman bayam yang ditumbuhkan pada dua konsentrasi NaCl 50 mM dan 100 mM dapat tumbuh dengan baik bahkan meningkat laju fotosintesisnya jika diberi perlakuan ALA 0.6 dan 1.8 mM. Daun bayam yang diberi perlakuan ALA memperlihatkan rasio asam askorbat teroksidasi/tereduksi yang rendah dan dan rasio glutathion tereduksi/teroksidasi yang tinggi jika dibandingkan kontrol dengan kondisi NaCl yang sama. Sedangkan aktivitas enzim antioksidan seperti askorbat peroksidase, katalase dan glutathion reduktase meningkat secara nyata, terutama setelah 3 hari setelah pemberian 0.6 dan 1.8 mM ALA. Data ini mengindikasikan bahwa perlindungan terhadap kerusakan oksidatif oleh jumlah antioksidan dan aktivitas enzim yang tinggi dan dengan lebih aktifnya siklus askorbat-glutathion mungkin terlibat dalam toleransi terhadap NaCl. Enzim yang terlibat pendetoksikasi senyawa radikal bebas (ROS) seperti superoksida dismutase, katalase, peroksidase merupakan protein yang mengandung heme. Oleh karena itu sintesis ALA yang lebih tinggi akan menyediakan senyawa heme yang lebih banyak bagi pembentukan enzim-enzim pendetoksikasi radikal bebas seperti superoksida, hydrogen peroksida dan radikal hidroksil. Hal ini adalah salah satu kunci penting dalam ketahanan terhadap cekaman salinitas. Pada penelitian lain menunjukkan bahwa pemberian ALA (tanpa cekaman) dapat meningkatkan jumlah fruktan (polifruktosilsukrosa) sebagai karbohidrat non-struktural pada tanaman rakkyo (Yoshida et al. 1996) dan bayam (Yoshida et al. 1995). Seperti diketahui bahwa fruktan merupakan gula alcohol yang berperanan sebagai osmoprotektan dalam menjaga turgor sel terutama pada cekaman kekeringan. Sehingga ekspresi gen ALAS mungkin dapat meningkatkan toleransi terhadap cekaman salinitas pada tanaman, karena pada cekaman salinitas air juga menjadi tidak tersedia karena tekanan osmotik diluar sel lebih tinggi. Gambar 29. Pengujian ketahanan terhadap cekaman salinitas. Tanda panah menunjukkan tanaman transgenik no. 4 yang tahan terhadap pemberian larutan NaCl 200 mM Kloning Gen Chlorophyll A Oxygenase (CAO) Kemampuan untuk hidup ditempat yang ternaungi ditentukan oleh kemampuan tanaman untuk memperbesar antena penangkap cahaya. Dalam keadaan cahaya redup, biasanya tanaman akan memperbesar antena sehingga jumlah cahaya yang diperlukan untuk proses fotosintesis dapat tercukupi. Oleh karena itu modifikasi genetika untuk mengatur ukuran antena mungkin sangat berguna dimasa depan dalam rangka meningkatkan produktifitas tanaman pertanian. Dalam penelitian ini telah dilakukan kloning gen CAO (1.6 kb) yang bertanggung jawab dalam biosintesis klorofil b. Klorofil b diketahui sangat berperan dalam menentukan ukuran antena. Gen CAO yang diklon ke dalam plasmid pAS900 (Gambar 30) ini telah dilakukan pengurutan nukleotidanya setelah dibandingkan dengan yang ada di GeneBank menunjukkan tidak adanya mutasi pada urutan nukleotidanya (Gambar 31). Gambar 30. Peta plasmid rekombinan pAS900-CAO1 yang membawa gen CAO ATGAACGCCG TTCTCTCCAG AGAGTGGATT AAAGGAAAGT GTGCTCGTCA TCGTGAGTAC CACCAACAGG TGGTTAATGA TTCTGCTGTC CCGCCTTATA CAATGGTACC TGTACGGAAT TGTCCGTACC AGGTGAAGAT ACCTGCACCT CTCCCGGTGG CCACTTTTGC ATACTGGGAT TCAAAACCCG GTTTACCTTC GAATCTTCCA GTTTTAGGAC TCGGAGTTCG ACTGAGCTC CCGTGTTTAG AAGAGTCAAT AGTTGAGAAG TTTTAGATGT AGATCTTCTT AAGTCCATCG TTCATATATC TAGGTTGTTA GAGTTAGATA GTCCACACTT AATTGAATGC ACATGTGCGC ATGGATGGGA CAAATCATTA ATACTTCCTT AACACGGTTT AAAAGGCTGG CCATATCCAA GGAAACTAGA TTCTAAAAAC TTCATGGAAC AACAAGAACG GTATAGACTA TCCTTCTGCT AAAGAAGGGC GGAGACCTTT TAATCAAGCT ACCATTATGA GTACAGTGAA TGAAGCAAGG CCTGGCAGAG GGGAAAAGAC GAAGAATTTT TTTGAACAAC ATAGAGCATG ATACTCAACC CCTTGTCTTG CTTTACAGCC ACTTCTAGAT AGTGTCCCAA TCGATATGGA AGGCAAAAGC AAGACAAGAC ATCTGTGGAG GATGTTAAAC TGGAGGAACG TTATCTCTCC GTGAAAGGAG GTTTGATGTG ATTGAAGTTG TTCTTCATGA GAAAGAACTA GTTTCGACTG TTGTAACGGA AAACACGGGT TGGTATCCCG CATGGGTTAT TCCTCTTGAT GATGGAGAAT AACAAGAAGG TCCATCAGGG AATCTCTTGG GTTTGGTGAA ATTTAAACCA ACACAGCAGT TTCTATACCG ACATTTCGCT GGAGCAAACA CAGTAGATCG CTATCTCCTT AATTTAGGGT GAGGATCCTA CTAGGTTTGA CAAGGTTGTT GCTGGATTGC CTTTAGACAA ATTAGATAAA GCGAAAAGCT TTGCTTTCAC CTTTAGGGGT CTTGGCACAG GTAAGAAGAT TATGATCTGG TTTTTAATTC ATCTTGCTCA GTTTTTAACA CCGTGTATTG GTGCAACACA AATGTCACTA GAACAGGTCT TATGGAATTT TGGCGATGAT CTCTAAAACC ATTTGCTGTG GATCAAAAGC TATTCAATAC GATGTACTTA AGGAAGAATT GTTAGCCCAC CCCTCCTCTT TGAATGTTTC TGCAGATCTC GAAGACGGGA TGAACGAGGG GCCGTCTACA ATTTGGCCCG ATGCTGAGCT TGCCCCATTC CCTACCTCGG TTTTATCGAC TCTTCATCAA GACTTTGCTC TAAACGAAGA ACCAGTTGCT AAACTACCTT CGATCCTCTT TTTGGTGATG TCCTCCTTAT TTGGATTGGC ATCCTCTAGC ATCGAAAGCA ATGGAAGAAT CAACCACTGC TGGTCCGGTT AAGCATGATA AACCAGGATG ACGTATTCAA AAGTTACTGA GTGATGAGCC TGTAATGGAC ACTCATACAT GTCTCCAAGG AATCGGGATA CTCCATGTCT CTATATTAAA TCTACGGCTA TATGACAAGC TCTCCGGCTA Gambar 31. Urutan nukleotida gen CAO dari Arabidopsis thaliana ecotype Columbia Daun di atas kanopi mendapatkan cahaya yang cukup untuk melakukan proses fotosintesis. Namun tidak demikian dengan daun yang berada dibawah kanopi. Oleh karena itu, laju fotosintesis menjadi berbeda-beda tergantung berapa cahaya yang dapat ditangkap oleh daun sesuai letaknya. Horton (2000) melaporkan pada padi, kontribusi daun bawah terhadap keseluruhan fotosintesis tanaman sangat signifikan meskipun laju fotosintesisnya lebih rendah dibandingkan daun atas yang mendapat cahaya penuh. Pembuangan daun ketiga dan keempat dapat menurunkan laju fotosintesis sampai 45%. Ekspresi gen CAO secara berlebih memungkinkan kita untuk memperbesar antena penangkap cahaya sehingga daun dibawah kanopi dapat menangkap cahaya secara lebih baik. Dengan demikian kontribusi daun bawah kanopi akan menjadi lebih signifikan dalam menyumbangkan fotosintat sehingga mungkin pada akhirnya dapat meningkatkan produksi tanaman. SIMPULAN DAN SARAN Simpulan Transformasi gen hemA ke dalam genom Arabidopsis thaliana ecotype Columbia telah berhasil dilakukan menggunakan Agrobacterium tumefaciens. Hasil analisis PCR membuktikan bahwa gen tersebut berhasil diintroduksikan ke genom Arabidopsis thaliana dan berdasarkan hasil analisis dengan RT-PCR juga dapat diekspresikan pada tarap RNA. Hasil analisis aktivitas protein ALAS menunjukkan adanya aktivitas sebesar 40.5 nmol.mg-1.h-1. Ekspresi ALAS menyebabkan peningkatan sintesis ALA sebesar 153.6% dan klorofil 43% dibandingkan tanaman kontrol. Hasil analisis berat basah dan berat kering pada tanaman transgenik menunjukkan kenaikan tertinggi sebesar 16.8% dan 16.6% dibandingkan kontrol. Namun satu diantaranya mengalami penurunan berat basah dan berat kering yang dikarenakan perkembangan akar yang jelek (transgenik no 3) sedangkan satu lagi mengalami penurunan jumlah biji yang dihasilkan (transgenik no 4). Ekspresi ALAS ini juga menyebabkan tanaman Arabidopsis transgenik mempunyai ketahanan terhadap cekaman salinitas sampai dengan 200 mM. Gen penyandi Chlorophyll A Oxygenase (CAO) berhasil diklon dari Arabidopsis thaliana dan hasil pengurutan nukleotida menunjukkan tidak terjadinya mutasi. Saran Ekspresi ALAS yang terkontrol mungkin dapat mengatasi abnormalitas pembungaan yang terjadi. Oleh karena itu pemilihan promoter menjadi penting untuk mengetahui pada fase mana ekspresi ALAS secara berlebih diperlukan. Pemakaian promoter yang diinduksi oleh cekaman juga diperlukan untuk menguji lebih lanjut potensi ALAS dalam meningkatkan ketahanan terhadap cekaman. Ekspresi secara bersama hemA dan CAO perlu dicoba untuk mengetahui efek sinergistik kedua gen ini dalam meningkatkan ukuran antena penangkap cahaya dan pengaruhnya terhadap laju fotosintesis pada daun dibawah kanopi DAFTAR PUSTAKA Alscher RG, Erturk N, Heath LS. 2002. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J Exp Bot 53:1331-1341 Amtmann A, Sanders D. 1999. Mechanisms of Na+ uptake by plant cells. Adv in Bot Res 29: 75-112 An G, Ebert PR, Mitra A, Ha SB. 1988. Binary vectors. In: Gelvin, SB, Schilperoot RA, Editor. Plant Molecular Biology Manual. Dordrecht, Netherlands: Kluwer Academic Publishers, p. A3:1-19 Ankenbauer, RG, Nester EW. 1990. Sugar-mediated induction of Agrobacterium tumefaciens virulence genes: structural specificity and activities of monosaccharides. J Bacteriol 172: 6442-6446. Apse MP, Aharon GS, Snedden WA, Blumwald E. 1999. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285:1256-1258. Beale SI, Gough SP, Granick S. 1975. Biosynthesis of δ-aminolevulinic acid from the intact carbon skeleton of glutamic acid in greening barley. Proc Natl Acad Sci USA 72:2719-2723 Bellemare G, Bartlett SG, Chua NH. 1982. Biosynthesis of chlorophyll a/bbinding polypeptides in wild type and the chlorina f2 mutant of barley. J Biol Chem 257:7762-7767 Bindu RC, Vivekanandan M. 1998a. Hormonal activities of 5-aminolevulinic acid in callus induction and micropropagation. Plant Growth Regulation 26:15-18 Bindu RC, Vivekanandan M. 1998b. Role of aminolevulinic acid in improving biomass production in Vigna catjung, V. mungo, and V. radiata. Biologia Plantarum 41:211-215 Blokhina O, Virolainen E, Fagerstedt KV. 2003. Antioxidants, oxidative damage and oxygen deprivation stress: a review. Ann Bot 91:179-194 Bossmann B, Knoetzel J, Jansson S. 1997. Screening of chlorina mutants of barley (Hordeum vulgare L.) with antibodies against light-harvesting proteins of PSI and PSII: Absence of specific antenna proteins. Photosynth. Res. 52:127-136 Boyer JS. 1982. Plant productivity and environment. Science 218: 443-448 Bradley LR. Kim JS, Matthysse AG. 1997. Attachment of Agrobacterium tumefaciens to Carrot Cells and Arabidopsis wound sites is correlated with the presence of a cell-associated, acidic polysaccharide. J Bacteriol 179:5372-5379. Bundock P, den Dulk-Ras A, Beijersbergen A, Hooykaas PJJ. 1995. Transkingdom T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. EMBO J. 14:3206–3214 Cangelosi GA, Ankenbauer RG, Nester EW. 1990. Sugars induce the Agrobacterium virulence genes through a periplasmic binding protein and a transmembrane signal protein. Proc Natl Acad Sci USA 87:6708- 6712. Chan MT, Chang HH, Ho S, Tong WF, Yu SM. 1993. Agrobacteriummediated production of transgenic rice plants expressing a chimeric alphaamylase promoter/beta-glucuronidase gene. Plant Mol Biol 22:491–506. Chang CH, Winans SC. 1992. Functional roles assigned to the periplasmic, linker and receiver domains of the Agrobacterium tumefaciens VirA protein. J Bacteriol 174:7033-7039. Clough SJ, Bent AF. 1998. Floral dip: a simplified method for Agrobacteriummediated transformation of Arabidopsis thaliana. Plant J 16:735-743 Demidchik V, Tester M. 2002. Sodium fluxes through non-selective cation channels in the plasma membrane of protoplasts from Arabidopsis thaliana roots. Plant Physiol 128:379–387 Deng W, Chen L, Peng WT, Liang X, Sekiguchi S, Gordon MP, Comai L, Nester EW. 1999. VirE1 is a specific molecular chaperone for the exported single-stranded-DNA-binding protein VirE2 in Agrobacterium. Mol Microbiol 31:1795–1807. Doty SL, Yu NC, Lundin JI, Heath JD, Nester EW. 1996. Mutational analysis of the input domain of the VirA protein of Agrobacterium tumefaciens. J Bacteriol 178: 961-970. Elstner EF. 1991. Mechanisms of oxygen activation in different compartments of plant cells. In: Pell EJ, Steffen KL, editor. Active oxygen/oxidative stress and plant metabolism. Rockville, MD. Am Soc Plant Physiol 13-25 Escoubs JM, Lomas M, Laroche J, Falkowski PG. 1995 Light intensity regulation of cab gene transcription is signaled by the redox state of the plastoquinone pool. Proc Natl Acad Sci USA 92:10237-10241 Espineda CE, Linford AS, Devine D, Brusslan JA (1999) The AtCAO gene, encoding chlorophyll a oxygenase, is required for chlorophyll b synthesis in Arabidopsis thaliana. Proc Natl Acad Sci USA 96:10507-10511 Flachmann R, Kuhlbrandt W. 1995. Accumulation of plant antenna complexes is regulated by post-transcriptional mechanisms in tobacco. Plant Cell 7: 149-160 Flachmann R 1997. Composition of photosystem II antenna in light-harvesting complex II antisense tobacco plants at varying irradiances. Plant Physiol 113:787-794 Flowers TJ, Troke PF, Yeo AR. 1977. The mechanism of salt tolerance in halophytes. Annu Rev Plant Physiol. 28:89-121. Fridovich I. 1986. Superoxide dismutase. Advances in Enzymology and Related Areas of Molecular Biology 58:61-97 Fu HH, Luan S. 1998. AtKUP1: a dual-affinity K+ transporter from Arabidopsis. Plant Cell 10:63-74. Fullner KJ, Lara JC, Nester EW. 1996. Pilus assembly by Agrobacterium TDNA transfer genes. Science 273:1107-1109. Gheysen G, Villarroel R, Van Montagu M. 1991. Illegitimate recombination in plants: a model for T-DNA integration. Genes Dev 5:287-297. Gibson KD, Laver WG, Neuberger A. 1958. Formation of δ-aminolevulinic acid in vitro from succinyl-coenzime a and glycine. Biochem J 70-71-81 Halliwell B, Gutteridge JMC. 1989. Protection against oxidants in biological system: The super oxide theory of oxygen toxicity. In: Halliwell B, Gutteridge JMC, editor. Free Radicals in Biology and Medicine Oxford: Clarendon Press; p. 86-123 Hamada A, Shono M, Xia T, Ohta M, Hayashi Y, Tanaka A, Hayakawa T. 2001. Isolation and characterization of a Na+/H+ antiporter gene from the halophyte Atriplex gmelini. Plant Mol Biol 46:35-42. Hasegawa PM, Bressan RA, Zhu JK, Bohnert HJ. 2000. Plant cellular and molecular responses to high salinity. Annu Rev Plant Physiol Plant Mol Biol 51:463-499. Hellens RP, Edwards EA, Leyland NR, Bean S. Mullineaux PM. 2000. pGreen: a versatile and flexible binary vector for Agrobacterium-mediated plant transformation. Plant Mol Biol 42:819-832 Hooykaas PJJ, Hofker M, den Dulk-Ras A, Schilperoort RA. 1984. A comparison of virulence determinants in an octopine Ti plasmid, a nopaline Ti plasmid, and an Ri plasmid by complementation analysis of Agrobacterium tumefaciens mutants. Plasmid 11:195–205. Hood EE, Gelvin SB, Melchers LS, Hoekema A. 1993. New Agrobacterium helper plasmids for gene transfer to plants. Transgene Res 2:208-218 Horton P. 2000. Prospects for crop improvement through the genetic manipulation of photosynthesis: morphological and biochemical aspects of light capture. J Exp Bot 51:475-485 Hotta Y, Tanaka T, Takaoka H, Takeuchi Y, Konnai M. 1997a. New physiological effects of 5-aminolevulinic acid in plant: The increase of photosynthesis, chlorophyll content, and plant growth. Biosci Biotech Biochem 61:2025-2028 Hotta Y, Tanaka T, Takaoka H, Takeuchi Y, Konnai M. 1997b. Promotive effects of 5-aminolevulinic acid on the yield of several crops. Plant Growth Regulation 22:109-114 Hotta Y, Watanabe K. 1999. Plant growth-regulating activities of 5aminolevulinic acid. Syokubutu-no-kagaku-Tyousetu 34:85-96 Huang Y, Morel P, Powell B, Kado CI. 1990. VirA, a coregulator of Ti-specified virulence genes, is phosphorylated in vitro. J Bacteriol 172:1142-1144. Ito H, Tanaka Y, Tsuji H, Tanaka A. 1993. Conversion of chlorophyll b to chlorophyll a by isolated cucumber etioplasts. Archives Biochem Biophys 306:148-151 Ito H, Takaichi S, Tsuji H, Tanaka A. 1994. Properties of synthesis of chlorophyll a from chlorophyll b in cucumber etioplasts. J Biol Chem 269:22034-22038 Ito H, Ohtsuka T, Tanaka A. 1996. Conversion of chlorophyll b to chlorophyll a via 7-hydroxymethyl chlorophyll. J Biol Chem 271:1475-1479 Ito H, Tanaka A. 1996. Determination of the activity of chlorophyll b to chlorophyll a conversion during greening of etiolated cucumber cotyledons by using pyrochlorophyllide b. Plant Phys Biochem Paris 34: 35-40 Jimenez A, Hernandez JA, Pastori G, del Rio LA, Sevilla F. 1998. Role of the ascorbate-glutathuone cycle of mitochondria and peroxisomes in the Senescence of pea leaves. Plant physiol 118:1327-1335. Jin S, Prusti RK, Roitsch T, Ankenbauer RG, Nester E.W. 1990a. The VirG protein of Agrobacterium tumefaciens is phosphorylated by the autophosphorylated VirA protein and this is essential for its biological activity. J Bacteriol 172:4945-4950. Jin S, Roitisch T, Christie PJ, Nester EW. 1990b. The regulatory VirG protein specifically binds to a cis acting regulatory sequence involved in transcriptional activation of Agrobacterium tumefaciens virulence genes. J Bacteriol 172:531-562. Jin S, Song Y, Pan S, Nester EW. 1993. Characterization of a virG mutation that confers constitutive virulence gene expression in Agrobacterium tumefaciens. Mol Microbiol 7:55-562. Kim EJ, Kwak JM, Uozumi N, Schroeder JI. 1998. AtKUP1, an Arabidopsis gene encoding high-affinity potassium transporter activity. Plant Cell 10:51-62. Kovach ME, Phillips RW, Elzer PH, Roop RM, Peterson KM. 1994. pBBR1MCS2: a broad-host range cloning vector. Biotechniques 16:800802 Kuttkat A, Edhofer I, Eichacker LA, Paulsen H. 1997. Light-harvesting chlorophyll a/b-binding protein stably inserts into etioplast membranes supplemented with Zn-pheophytin a/b. J Biol Chem 272:20451-20455. Lassner MW, Peterson P, Yoder JI. 1989. Simultaneous amplification of multiple DNA fragments by polymerase chain reaction in the analysis of transgenic plants and their progeny. Plant Mol Biol Rep 7:116-128 Lazof DB, Bernstein N. 1999. The NaCl induced inhibition of shoot growth: the case for distributed nutrition with special consideration of calcium. Adv Bot Res 29:113-189. Lehman CW, Trautman JK, Carroll D. 1994. Illegitimate recombination in Xenopus: characterization of end-joined junctions. Nucleic Acid Research 22:434-442. Li W, Guo G, Zheng G. 2000. Agrobacterium-mediated transformation: state of the art and future prospect. Chinese Sci Bull 45:1537-1546 Matile P, Hortensteiner S, Thomas H, Krautler B. 1996. Chlorophyll breakdown in senescent leaves. Plant Physiol 112:1403-1409 May MJ, Vernoux T, Leaver C, Van Montagu M, Inze D. 1998. Glutathione homeostasis in plants: implications for environmental sensing and plant development. J Exp Bot 49: 649-667. Maxwell DP, Laudenbach DE, Huner NPA. 1995 Redox regulation of lightharvesting complex II and cab mRNA abundance in Dunaliella salina. Plant Physiol 109: 787-795 Meinke DW, Cherry JM. 1998. Arabidopsis thaliana: A model plant for genome analysis. Science 282: 662 Neidle E, Kaplan S. 1993a. Expression of the Rhodobacter sphaeroides hem A and hem T genes, encoding two 5-aminolevulinic acid synthase isozymes. J Bacteriol 175:2292-2303 Neidle E, Kaplan S. 1993b. 5-Aminolevulinic acid availability and control of spectral complex formation in HemA and HemT mutants of Rhodobacter sphaeroides. J Bacteriol 175:2304-2313 Nishihara E, Kondo K, Parvez MM, Takahashi K, Watanabe K, Tanaka K. 2003. Role of 5-aminolevulinic acid (ALA) on active oxygen-scavenging system in NaCl-treated spinach (Spinacia oleracea). J Plant Physiol 160:1085-1091 Noctor G, Foyer CH. 1998. Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49: 249-279. Ohtsuka T, Ito H, Tanaka A. 1997. Conversion of chlorophyll b to chlorophyll a and the assembly of chlorophyll with apoproteins by isolated chloroplasts. Plant Physiol 113:137-147 Oster U, Tanaka R, Tanaka A, Rudiger W. 2000. Cloning and functional expression of the gene encoding the key enzyme for chlorophyll b biosynthesis (CAO) from Arabidopsis thaliana. Plant J 21:305-10 Pan SQ, Charles T, Jin S, Wu ZL, Nester EW. 1993. Preformed dimeric state of the sensor protein VirA is involved in plant-Agrobacterium signal transduction. Proc Natl Acad Sci USA 90:9939-9943. Parkinson JS. 1993. Signal transduction schemes of bacteria. Cell 73: 857-871. Peoples MB, Gifford RM. 1990. Regulation of the transport of nitrogen and carbon in higher plants. In Dennis DT, Layzell DB, Lefebvre DD, Turpin DH, editor. Plant Metabolsm, 2nd. Essex, England. Addison Wesley Longman. p 525-538 Puchta H. 1998. Repair of genomic double-strand breaks in somatic cells by oneside invasion of homologous sequences. Plant J 13:331-339. Rhoades JD, Loveday J. 1990. Salinity in irrigated agriculture. Agronomists Monograph 30:1089-1142. Am Soc Richards RA. 2000. Selectable traits to increase crop photosynthesis and yield of grain crops. J Exp Bot 51-447-458 Rebeiz CA, Montazer-Zouhool A, Hopen HJ, Wu SM. 1984. Photodynamic herbicides. I. Concept and phenomenology. Enzyme Microbiol Technol 6:390-401 Rebeiz CA, Montazer-Zouhool A, Mayasich JM, Tripathy BC, Wu SM, Rebeiz CC. 1988. Photodynamic herbicides. Recent development and molecular basis of selectivity. CRC Critical Rev Plant Sciences 6:385-436 Rubio F, Gassmann W, Schroeder JI. 1995. Sodium driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 270:1660-1663. Sasaki K, Tanaka T, Nishizawa Y, Hayashi M. 1990. Production of a herbicide, 5-aminolevulinic acid, by Rhodobacter sphaeroides using the effluent waste from an anaerobic digestor. Appl Microbiol Biotechnol 32:727-731 Sasaki K, Watanabe M, Tanaka T, Tanaka T. 2002. Biosynthesis, biotechnological production and applications of 5-aminolevulinic acid. . Appl Microbiol Biotechnol 58:23-29 Scheumann V, Ito H, Tanaka A, Schoch S, Ruediger W. 1996a Substrate specificity of chlorophyll(ide) b reductase in etioplasts of barley (Hordeum vulgare L.). Eur J Biochem 242:163-170 Sen Gupta A, Heinen JL, Holaday AS, Allen RD. 1993. Increased resistance to oxidative stress in transgenic plants that overexpress chloroplast Cu/Zn superoxide dismutase. Proc Natl Acad Sci USA 90:1629-1633 Scheumann V, Helfrich M, Schoch S, Ruediger W. 1996b. Reduction of the formyl group of zine pheophorbide b in vitro and in vivo: A model for the chlorophyll b to a transformation. Zeitschrift fuer Naturforschung Section C J Biosci 51:185-194 Scheumann V, Schoch S, Ruediger W. 1998. Chlorophyll a formation in the chlorophyll b reductase reaction requires reduced ferredoxin. J Biol Chem 273:35102-35108 Scheumann V, Schoch S, Ruediger W. 1999. Chlorophyll b reduction during senescence of barley seedlings. Planta 209:364-370. Sen-Gupta A, Webb RP, Holaday AS, Allen RD. 1993. Overexpression of superoxide dismutase protects plants from oxidative stress. Plant Physiol 103:1067-1073 Shimoda N, Toyoda-Yamamoto A, Nagamine J, Usami S, Katayama M, Sakagami Y, Machida Y. 1990a. Control of expression of Agrobacterium tumefaciens genes by synergistic actions of phenolic signal molecules and monocaccharides. Proc Natl Acad Sci USA 87:6684-6688. Shi H, Quintero FJ, Pardo JM, Zhu JK. 2002a. The putative plasma membrane Na+/H+ antiporter SOS1 controls long distance Na+ transport in plants. Plant Cell 14: 465-477. Shi H, Xiong L, Stevenson B, Lu T, Zhu JK. 2002b. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell 14 575-588. Shimoda N, Toyoda-Yamamoto A, Aoki, Machida Y. 1990b. Genetic evidence for an interaction between the VirA sensor protein and the ChvE sugar binding protein of Agrobacterium tumefaciens. J Biol Chem 268:2655226558. Smirnoff N. 2000. Ascorbic acid: metabolism and functions of a multi-facetted molecule. Curr Opinion Plant Biol 3:229-235 Sundberg C, Meek L, Carroll K, Das A, Ream W. 1996. VirE1 protein mediates export of the single-stranded DNA-binding protein VirE2 from Agrobacterium tumefaciens into plant cells. J Bacteriol 178:1207–1212. Sundberg CD, Ream W. 1999. The Agrobacterium tumefaciens chaperone-like protein, VirE1, interacts with VirE2 at domains required for singlestranded DNA binding and cooperative interaction. J Bacterial 181:6850–6855. Sun K, Hauser B. 2001. Salt stress induces anatomical changes in ovules and embryos, ultimately resulting in seed abortion. The 12th International Meeting on Arabidopsis Research. June 23-27. Madison, WI. USA. TAIR (The Arabidopsis Information Resource). 2003. About Arabidopsis thaliana. http://www.arabidopsis.org/info/aboutarabidopsis.html [13 Juli 2004] Tanaka H, Takakashi K, Hotta T, Takeuchi Y, Konnai M. 1992. Promotive effects of 5-aminolevulinic acid on yield of several crops. In: Proceedings of the 19th annual meeting of Plant Growth Regulator Society of America, San Francisco. Plant Growth Regulator Society of America, Washington DC, pp 237-241. Tanaka Y, Tanaka A, Tsuji H. 1993. Effects of 5-aminolevulinic acid on the accumulation of chlorophyll b and apoprotein of the light-harvesting chlorophyll a/b-protein complex of photosystem II. Plant Cell Physiol 34:365-370 Tanaka A, Ito H, Tanaka R, Tanaka Nobuaki K, Yoshida K, Okada K. 1998. Chlorophyll a oxygenase (CAO) is involved in chlorophyll b formation from chlorophyll a. Proc Natl Acad Sci USA 95:12719-12723 Tanaka R, Koshino Y, Sawa S, Ishiguro S, Okada K, Tanaka A. 2001. Overexpression of chlorophyllide a oxygenase (CAO) enlarges the antenna size of photosystem II in Arabidopsis thaliana. Plant J 26: 365373 Tanaka R, Tanaka A. 2005. Mini Review. Chlorophyll b is not just an accessory pigment but a regulator of the photosynthetic antenna. www.lowtem.hokudai.ac.jp/~ayumi/eng/Minireview-tanaka.pdf [24 April 2005] The Arabidopsis Genome Initiative. 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408:796-815 Thomas CE, Mclean LR, Parker RA, Ohlweiler DF. 1992. Ascorbate and phenolic antioxidant interactions in prevention of liposomal oxidation. Lipids 27:543-550 Tinland B, Schoumacher F, Gloeckler V, Bravo AM, Angel M, Hohn B. 1995. The Agrobacterium tumefaciens virulence D2 protein is responsible for precise integration of T-DNA into the plant genome. EMBO J 14:35853595. Tomitani A, Okada K, Miyashita H, Matthijs HC, Ohno T, Tanaka A. 1999. Chlorophyll b and phycobilins in the common ancestor of cyanobacteria and chloroplasts. Nature 400:159-62 Turk SCHJ, van Lange RP, Sonneveld E, Hooykaas PJJ. 1993. The chimeric VirA-Tar receptor protein is locked into highly responsive state. J Bacteriol 175:5706-5709. Uozumi N, Kim EJ, Rubio F, Yamaguchi T, Muto S, Tsuboi A, Bakker EP, Nakamura T, Schroeder JI. 2000. The Arabidopsis HKT1 gene homolog mediates inward Na+ currents in Xenopus laevis oocytes and Na+ uptake in Saccharomyces cerevisiae. Plant Physiol 122:1249-1259. Urata G, Granick S. 1963. Biosynthesis of δ-aminoketones and the metabolism of aminoacetone. J Biol Chem 238:881-820 Vance CP. 1990. The molecular biology of N metabolism. In Dennis DT, Layzell DB, Lefebvre DD, Turpin DH, editor. Plant Metabolsm, 2nd. Essex, England. Addison Wesley Longman. p 449-477 Vieira J, Messing J. 1991. New pUC-derived cloning vector with different selectable markers and DNA replication origins. GENE 100:189-194 Vincentini F, Hortensteiner S, Schellenberg M, Thomas H, Matile P. 1995. Chlorophyll breakdown in senescent leaves: Identification of the biochemical lesion in a stay-green genotype of Festuca pratensis Huds. New Phytologist 129:247-252 Wang, LJ, Jiang, WB, Huang, BJ. 2004. Promotion of 5-aminolevulinic acid on photosynthesis of melon (Cucumis melo) seedlings under low light and chilling stress conditions. Physiologia Plantarum 121: 258-264. Watanabe K, Tanaka T, Hotta Y, Kuramochi H, Takeuchi Y. 2000. Improving salt tolerance of cotton seedling with 5-aminolevulinic acid. Plant Growth Reg 32:99-103 Weinstein JD, Beale SI. 1983. Separate physiological roles and subcellular compartments for two tetrapyrrole biosynthesis pathways in Euglena gracilis. J. Biol Chem 258:6799-6807 Winans, SC. 1992. Two-way chemical signaling in Agrobacterium-plant interactions. Microbiol Rev 56:12-31. Von Wettstein D, Gough S, Kannagara CG. 1995. Chlorophyll biosynthesis. Plant Cell 7:1039-1057 Wu SJ, Ding L, Zhu JK. 1996. SOS1, a genetic locus essential for salt tolerance and potassium acquisition. Plant Cell 8:671-627. Xiang C, Han P, Oliver DJ. 1999. In solium selection for Arabidopsis transformants resistant to kanamycin. Plant Mol Biol Rep 17:59-65 Yoshida R, Tanaka T, Hotta Y. 1995. Physiological effects of 5-aminolevulinic acid in vegetable crops. Abs of 15th International Conference on Plant Growth Substances, Minneapolis, Minnesota, USA, p 417 Yoshida R, Hotta Y, Tanaka T, Takeuchi Y, Konnai M. 1996a. Promotive effects of 5-aminolevulinic acid on rice plants. Crop Reserch in Asia: Achivements and Perspective(ACSA) 524-525 Yoshida R, Tanaka T, Hotta Y.1996b. Regulation of fructan accumulation in rakkyo (Allium bakeri) and sallot (Allium ascalonicum) by 5aminolevulinic acid. Proc of the 3rd Joint PGRSA-JSCPR Meeting, Calgary, Alberta, Canada, p 177-182 Yoshida R, Fukuta Y, Shimotsubo K, Iwai K, Watanabe S, Toru Tanaka. 2004. Growth promotive effects of 5-aminolevulinic acid in the presence of microelements on yield in Komatsuna, Brassica campestris var.perviridis under alkaline soil conditions. Proc of the 4th International Crop Science Congress Brisbane, Australia, 26 Sep – 1 Oct 2004 Zavgorodnyaya A, Papenbrock J, Grimm B. 1997. Yeast 5-aminolevulinate synthase provides additional chlorophyll precursor in transgenic tobacco. Plant J 12:169-178 Zhu JK. 2001a. Plant salt tolerance. Trends Plant Sci. 6:66-71. Zhu JK. 2001b. Cell signaling under salt, water and cold stresses. Curr Opin Plant Biol 4:401-406. Zhu JK. 2003. Regulation of ion homeostasis under salt stress. Curr Opin Plant Biol 6:441-445