pengaruh superovulasi pada laju ovulasi, sekresi estradiol
advertisement

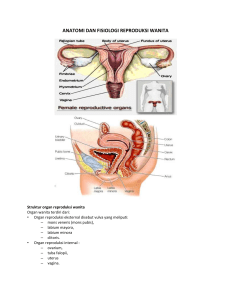
PENGARUH SUPEROVULASI PADA LAJU OVULASI, SEKRESI ESTRADIOL DAN PROGESTERON, SERTA PERTUMBUHAN DAN PERKEMBANGAN UTERUS DAN KELENJAR SUSU TIKUS PUTIH (Rattus Sp.) SELAMA SIKLUS ESTRUS TESIS OLEH : HERNAWATI PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR 2001 endometrium dan miometriurn (Hafez, 1993). Pada tikus, enam jam setelah penyuntikan estrogen, respon pertama yaitu pembendungan air dalam jaringan uterus (Partodihardjo, 1992), peningkatan kadar DNA, RNA, sintesis protein, clan aktivitas enzim (Frandson, 1996). Estrogen dapat menyebabkan meningkatnya vaskularisasi dan aktivitas mitosis uterus yang lebih besar, mengakibatkan organ bertambah berat. Pada tikus terapi dengan estrogen menyebabkan akumulasi air pada lumen uterus (Nalbandov, 1990). Upaya untuk menyiapkan uterus menjadi lingkungan yang cocok untuk pertumbuhan dan perkembangan embrio serta fetus, estradiol bekerja dengan cara mempengaruhi sintesis kolagen sehingga merubah struktur kolagen uterus (Pastore et al., 1992), meningkatkan kandungan glikogen pada uterus dengan mempengaruhi aktivitas glikogen sintetase (Williams dan Provine, 1966), dan meningkatkan metabolisme fosfolipid (Gould et al., 1978), serta meningkatkan dan proliferasi sel-sel uterus (Yarnashita et al., 1990). pada uterus melalui peningkatan ekspresi reseptor sintesis DNA Estradiol juga bekerja progesteron (Kraus dan Katzenellebogen , 1993). Kelenjar Susu Tikus mempunyai 12 buah kelenjar susu, tiga pasang berada di bagian thorak dan tiga pasang di bagian abdominal-inguinal (Morrow, 1986; Harknes dan Wagner, 1989). Kelenjar susu dianggap homolog dengan kelenjar keringat, karena keduanya berasal dari kulit yang turnbuh ke dalam. Setiap kelenjar terdiri atas beberapa lobus Hormon-hormon Ovarium Fungsi utama ovarium adalah memproduksi ova dan membuat hormon rerpoduksi. Hormon yang dibuat oleh ovarium adalah yang termasuk kelompok steroid, seperti estrogen, progesteron, adrogen dan hormon non steroid (peptida) yaitu relaksin (Binkley, 1995). Hormon steroid menimbulkan respon terhadap aktivitas reproduksi seperti sifat seksual sekunder, perilaku persiapan kawin, mempersiapkan uterus untuk implantasi blastosit, menyiapkan perkembangan kelenjar susu untuk memproduksi susu, dan mengatur kontraksi uterus pada saat kelahiran (Hafez, 1993). Diagram yang memperlihatkan pengaturan siklus reproduksi pada hewan betina disajikan pada Gambar 4. Estrogen dihasilkan oleh sel teka interna dan granulosa folikel ovarium, korpus luteum, plasenta, dan dalam jumlah kecil oleh korteks adrenal dan testis. Estrogen mempunyai kontrol umpan balik positif terhadap hipotalamus dalam mengubah LH pada ovarium dari fase folikuler menjadi fase luteal dan mempunyai kontrol umpan balik negatif terhadap pituitari anterior dalam mengatur sekresi FSH d m LH (Ganong, 1995; Binkley, 1995). Estrogen bersama-sama FSH dapat merangsang pertumbuhan sel-sel * granulosa secara mitosis. Faktor inilah yang menyebabkan terpisahnya sel-sel granulosa sehingga membentuk folikel. Selain itu estrogen dan FSH bekerja secara sinergis dalam menaikkan sensitivitas reseptor untuk estradiol dan estradiol sendiri akan merangsang proliferasi sel-sel granulosa, menaikkan sensitivitas reseptor untuk FSH, peningkatan CAMP dan merangsang FSWLH dalarn menginduksi - reseptor LH (Saxena dan Rathrnan, 1982 dalam Yusuf, 1990). Dijelaskan pula bahwa FSH dan LH menyebabkan pertambahan besar folikel dan dibutuhkan untuk proses ovulasi serta pembentukan korpus luteurn. Impuls saraf hipoblamus positif pada hipotalamus " . . inhibin pituitrari - 1 U II F,, \ estrogen \ Cairan antnl .. inhibin activin ) FASE FOLIKULER progesteron I LUTEAL Gambar 4. Diagram yang memperlihatkan pengaturan siklus reproduksi pada hewan betina. (Sumber: Binkley, 1995) Kerja estrogen pada organ kelamin asesoris umumnya dikaitkan dengan perilaku estrus yang khas pada seekor hewan. Di samping itu estrogen dapat merangsang aktivitas muskular tuba uterus dan menaikkan kepekaan organ tersebut. Perubahan yang terjadi pada uterus yang dirangsang oleh estrogen adalah peningkatan kadar air dalarn sel, DNA, RNA, sintesis protein dan aktivitas enzim (Frandson, 1996). Estradiol berperan pula untuk rnemelihara korpus luteum agar tetap mensekresikan progesteron. Kehadiran estradiol di korpus luteum sesuai dengan fungsinya yaitu untuk merangsang biosintesis kolesterol, mengatur aktivitas asilCoA: kolesterol asiltranferase (ACAT) agar tersedia kolesterol bebas untuk pembentukan hormon steroid progesteron (Azhar et al., 1989). Konsentrasi estradiol dalarn serum induk meningkat secara drastis sebelum ovulasi (McDonald, 1980) kemudian menurun dan naik sesuai perkembangan umur kebuntingan (hcketts dan Flint, 1980; Sheldrick et al., 1981; Sumaryadi dan Manalu, 1995a; Manalu dan Sumaryadi, 1995b). serum induk mencapai 56,397 + 9,163 Pada tikus, konsentrasi estradiol dalam pg/ml sebelum kebuntingan, kemudian menurun pada urnur kebuntingan 4 hari (42,717 2 0.0016 pglml) sampai umur kebuntingan 12 hari (43,7 12 5 1.795 pg/ml), selanjutnya konsentrasi estradiol \ melonjak secara drastis hingga mencapai konsentrasi tertinggi (68,268 2 1,919 pg/ml) pada umur kebuntingan 16 hari dan selanjutnya menurun pada umur kebuntingan 20 hari (5 1,951 + 1,947 pg/ml) yaitu menjelang kelahiran (Tuju dan Manalu 1996a). Progesteron merupakan hormon yang disekresikan oleh korpus luteum, plasenta, dan kelenjar adrenal. Progesteron diedarkan ke dalam darah karena suatu 8 Metode Penelitian Rancangan percobaan Sebanyak 80 ekor tikus putih betina yang sudah dewasa kelamin dikelompokkan dalam suatu rancangan acak lengkap pola faktorial4x4 dengan 5 ekor tikus sebagai ulangan untuk setiap unit percobaan. Faktor pertama adalah dosis penyuntikan PMSG (0, 37.5, 75 dan 150 I.U.per kilogram bobot badan). Faktor kedua adalah fase siklus estrus (folikuler, luteal 1, luteal2, dan luteal 3). Penentuan level dosis PMSG di atas didasarkan pada penelitian sebelumnya yang telah dilakukan oleh Miller dan Armstrong (1981) pada tikus yang belum dewasa dengan memberikan dosis PMSG secara bertingkat pada dosis 4, 8, 16, dan 40 I.U. untuk per ekor tikus. Pada penelitian ini dicoba memberikan PMSG pada tikus yang sudah dewasa yaitu mulai dosis terendah 37.5 I.U. per kilogram bobot badan, kemudian ditingkatkan dua kali lebih tinggi dari 37.5 yaitu menjadi 75 I.U. per kilogram bobot badan, dan selanjutnya ditingkatkan dua kali dosis 75 yaitu 150 I.U. per kilogram bobot badan sebagai dosis tertinggi. Peubah yang diukur selarna penelitian adalah konsentrasi estradiol dan progesteron, jumlah folikelkorpus luteum, bobot ovarium, uterus, dan kelenjar susu, * bobot kering uterus, BKBL kelenjar susu, serta komponen biokimia uterus dan kelenjar susu meliputi kandungan total dari DNA, RNA, glikogen dan kolagen. Data dianalisis dengan metode sidik ragam (Anova) dilanjutkan dengan uji Duncan multiple range test (DMRT)(Steel and Tome, 1993). Selanjutnya korelasi antara estradiol dan progesteron dengan parameter lain yang diukur diolah dengan luteum yang terbentuk dapat disebabkan meningkatnya sensitivitas sel-sel ova di dalam ovarium terhadap rangsangan hormon. Total jumlah folikelkorpus luteum pada fase siklus yang berbeda menunjukkan bahwa luteal hari ke satu berbeda nyata dengan luteal hari ke tiga (P<0.01). Secara numerik jurnlah korpus luteum pada luteal hari ke satu sampai luteal hari ke tiga cenderung menurun. Beberapa kemungkinan mengapa jumlah korpus luteum pada fase luteal cenderung menurun. Pertama; kemungkinan karena sejumlah korpus luteum telah mengalami peluruhan seiring dengan waktu siklus yang berubah. Kedua; kemunglunan karena korpus luteum yang terbentuk pada fase luteal memang sedikit. Hal ini dapat disebabkan adanya kompetisi dari folikel-folikel yang banyak tumbuh di dalam ovarium, sehingga hanya sebagian dari folikel-folikel tersebut yang diovulasikan dan akhirnya korpus luteum yang terbentuk hanya sedikit. yang kurang tepat. . Ketiga; penentuan antara bentuk folikel dan korpus luteum Jadi ada kemungkinan pada fase luteal lebih banyak folikel I ' yang dihitung dan bukan korpus luteurn. Data percobaan pengaruh penyuntikan hormon gonadotropin (PMSG dan HCG) pada bobot ovarium tikus selama siklus estrus disajikan pada Tabel 2. Dari ' tabel tersebut dapat dilihat bahwa total bobot ovarium tikus yang disuperovulasi pada dosis 37.5,75, dan 150 I.U.lebih berat (P<O.Ol)dibandingan kontrol selama siklus estrus. Hal ini kemungkinan oleh banyaknya folikelkorpus luteum yang berkembang di dalam ovarium yang disebabkan perangsangan hormon gonadotropin * Tabel Larnpiran 15. Daftar sidik ragarn kandungan total glikogen kelenjar susu tikus yang disuperovulasi dengan level dosis PMSG dan fase siklus estrus berbeda yang diamati selama siklus estrus Sumber Keragaman Model Galat Total R-kuadrat 0.87219 Sumber Keragaman Luted Dosis Luted*Dosis DB JK KT Fhit 15 64 79 2282.30077 334.4401 8 26 16.74095 152.15338 5.22563 29.12 Akar MSE 2.28596 K.V. 16.1 1210 Pr > F 0.0001 Rata-rata Glikogen 14.18786 DB Anova JK KT Fhit Pr > F 3 3 9 773.52490 559.21790 949.55797 257.84163 186.40597 105.50644 49.34 35.67 20.19 0.0001 0.0001 0.0001 Tabel Larnpiran 16. Dafiar sidik ragam kandungan total kolagen kelenjar susu tikus yang disuperovulasi dengan level dosis PMSG dan fase siklus estrus berbeda yang diamati selama siklus estrus Model Galat Total DB JK 15 64 79 949.65 177 359.07087 1308.72264 KT 63.31012 5.61048 Fhit 11.28 Pr > F 0.0001 >' R-kuadrat 0.72563 Sumber Keragaman Luteal Dosis Luteal*Dosis K.V. 16.58724 Akar MSE 2.36865 Rata-rata Kolagen 14.27992 DB Anova JK KT Fhit 3 478.07919 356.85635 114.71623 159.35973 1 18.95212 12.74625 28.40 21.20 2.27 3 9 Pr > F 0.0001 0.0001 0.0281