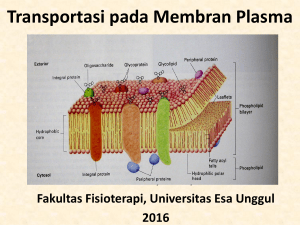
BAB III CILIOPHORA Ciliophora merupakan protozoa
advertisement

BAB III
CILIOPHORA
Ciliophora merupakan protozoa yang memiliki silia sebagai alat geraknya.
Protozoa merupakan binatang yang paling banyak di dunia. Ciliophora, disebut
juga Ciliata, berperan sebagai konsumen bakteri (Prokaryotes). Dimana bakteri
memainkan peranan penting dalam menjaga bumi sebagai tempat yang cocok
untuk
tempat tinggal dan protozoa memainkan peranan penting dalam
mengendalikannya.
Istilah Protozoa berasal dari bahasa Yunani, yaitu protos berarti
pertama dan zoon
termasuk
berarti
Protista yang
hewan.
Sesuai
menyerupai
dengan klasifikasi,
hewan.
Kelompok
ini
Protozoa
mulanya
“dibentuk” untuk mengelompokan organisme yang bukan tumbuhan dan
bukan
hewan.
Itulah sebabnya Protozoa disebut organisme seperti hewan
(animal like).
Sebagian besar Protozoa uniseluler memiliki ukuran tubuh antara (2-1.000) µm, protozoa termasuk eukariot. Biasanya hidup di dalam air, namun
ada juga yang ditemukan di dalam tanah bahkan di dalam tubuh organisme
lain sebagai parasit. Di perairan laut ataupun air tawar, Protozoa berperan
sebagai zooplankton.
Ciliata atau Infusoria merupakan kelompok terbesar di Protozoa, di
mana anggotanya sekitar 8.000 species. Ciri khas filum ini adalah alat
geraknya berupa cilia (rambut getar). Cilia tersebut ada yang terdapat di
seluruh tubuh, ada pula yang hanya di bagian tertentu. Selain sebagai alat gerak,
cilia pun berguna membantu mengumpulkan makanan. Habitat kelompok ini
adalah air tawar dan air laut yang mengandung zat organik tinggi.
A. Morfologi Phylum Ciliophora
Kehadiran silia yang menutupi sel dari organisme merupakan ciri khas
utama dari kelompok ini, oleh karena itu dinamai phylum Ciliophora. Namun,
perbedaan sitologi utama adalah kehadiran dua jenis inti, yaitu mikronukleus dan
macronucleus. Radiasi adaptif kelompok ini selama evolusi telah menghasilkan
31
beberapa spesies sangat bagus dan beragam. Beberapa adalah sessile (misalnya
Suctorians atau Stentor) dan menangkap makanan dengan tentakel yang
menembus sitoplasma dan menarik mangsa, atau dengan kompleks membranelles
mendorong air yang membawa partikel aliran air ke dalam rongga vakuola
makanan bucal yang terbentuk. Meskipun sebagian besar ciliates "telanjang",
beberapa menghasilkan mineralisasi Lorica (Tintinnids) atau mensekresi skala
organik (misalnya Lepidotrachelophyllum). Penyusunan silia pada permukaan
tubuh dan di wilayah aparatus oral, kehadiran struktur makanan yang khusus, dan
organisasi dari pita subpellicular dari mikrotubulus adalah kriteria penting yang
digunakan dalam menciptakan kategori taksonomi (Roger, 1988).
Anggota subphylum Ciliophora memiliki silia atau derivat ciliary pada
beberapa tahap siklus hidup. Perlengkapan berkisar dari penutup lengkap dari silia
sederhana untuk sebuah membranelles yang relatif sedikit lebih atau kurang
lengkap terbatas pada daerah peristomial. Dalam rentang ini, tipe spesialisasi
ciliary dan pola distribusi menjadi dasar utama untuk membedakan taksonomi
subdivisi. Ciliophore ini biasanya dibagi menjadi dua kelas, Ciliatea dan Suctorea.
Dalam ciliatea, silia atau komponen derivat mereka terdapat dalam fase siklus
dominan. Suctorea tidak bersilia pada fase dewasa dan telah mengembangkan
tentakel khusus yang berfungsi dalam mencari makan. Tahapan larva bersilia
merupakan karakteristik spesies yang paling menghubungkan kelompok ini
dengan Ciliatea dan pola larva ciliary menyiratkan bahwa Suctorea lebih erat
terkait dengan Holotrichida daripada ke ciliates yang lebih spesifik (Hall, 1961).
32
B. Klasifikasi subphylum Ciliophora
Klasifikasi subphylum Ciliophora yang diadopsi oleh Jahn dan Jahn dalam
Hall (1961) adalah sebagai berikut.
Sub phylum
Class 1
: Ciliophora
: Ciliatea
Sub class 1 : Protociliatia
Ordo 1
: Opalinida
Subclass 2 : Euciliatia
Ordo 1
: Holotrichida
Sub ordo 1
: Astomina
Sub ordo 2
: Gymnostomina
Sub ordo 3
: Trichostomina
Sub ordo 4
: Hymenostomina
Sub ordo 5
: Thigmotrichina
Sub ordo 6
: Apostomina
Ordo 2
: Spirotrichida
Sub ordo 1
: Heterotrichina
Sub ordo 2
: Tintinnina
Sub ordo 3
: Oligotrichina
Sub ordo 4
: Entodiniomorphina
Sub ordo 5
: Hypotrichina
Sub ordo 6
: Ctenostomina
Ordo 3
: Peritrichida
Ordo 4
: Chonotrichida
Class 2
: Suctorea
C. Sejarah Kehidupan, Ekologi, dan Reproduksi pada Phylum Ciliophora
Kebanyakan ciliata hidup bebas. Relatif sedikit yang parasit, dan hanya
satu spesies, Balantidium coli yang diketahui menyebabkan penyakit pada
manusia. Beberapa ciliata lainnya menyebabkan penyakit pada ikan, yang lainnya
adalah parasit atau commensals pada berbagai invertebrata. Sebagian besar yang
lain hidup di saluran pencernaan mamalia, di mana mereka menjalankan aktivitas
33
untuk menstabilkan populasi besar bakteri simbiotik yang memecah selulosa
dalam makanan hewan.
Ciliata yang hidup bebas dapat memakan bakteri, ganggang, atau bahkan
ciliata lainnya; Didinium adalah pemburu yang rakus dan konsumen ciliata
lainnya. Beberapa ciliata bersimbiosis dengan bakteri atau ganggang. Ciliata yang
hidup bebas dapat ditemukan hampir di mana saja di air, namun bentuk yang
berbeda mendominasi dalam habitat yang berbeda. Ciliata dalam tanah cenderung
berbentuk kecil yang dapat membentuk kista resisten untuk bertahan hidup lama
ketika kondisi kering. Tintinnids berlimpah di plankton laut, di mana mereka dan
ciliates lain mungkin mengkonsumsi sampai 90% dari produksi bakteri plankton
dan ganggang. Ciliates besar umum di lingkungan air tawar, khususnya air yang
telah diperkaya zat organik, misalnya oleh limbah (Linn dan Small, 1991).
Ciliata bereproduksi secara aseksual dengan pembelahan: mikronukleus
mengalami mitosis, sedangkan pada sebagian besar ciliates, macronucleus hanya
terpisah menjadi dua. Namun, ciliata juga bereproduksi secara seksual, melalui
proses yang dikenal sebagai konjugasi. Konjugasi sering disebabkan oleh
kekurangan makanan. Dua ciliata dengan tipe kawin yang berlawanan datang
mendekat, bersama-sama dan membentuk sebuah jembatan sitoplasmik antara dua
sel, membagi micromuclei oleh meiosis, macronuclei hancur, dan konjugasi selsel haploid micronuclei akan tertukar melalui koneksi sitoplasma. Mereka
kemudian memisahkan macronuclei, baru reformasi dari micronuclei, dan
membagi. Esensi reproduksi seksual adalah membentuk organisme baru dari
gabungan bahan genetik dari orang tua. Setelah konjugasi, masing-masing
pasangan Ciliata telah mengakuisisi materi genetik baru, dan membagi
menimbulkan progeni dengan kombinasi gen baru. Hal ini penting untuk
kelangsungan hidup garis keturunan Ciliata; ciliata paling tidak dapat
mereproduksi selamanya dengan pembelahan aseksual, dan akhirnya mati jika
tidak terjadi konjugasi (Linn dan Small, 1991).
D. Kelas Ciliatea
Silia yang tersusun dari organel siliari merupakan tahap aktif dari siklus
kehidupan. Siliata dibagi ke dalam sejumlah kelompok. Kelas Ciliatea mencakup
34
Subkelas Protociliatia dan Euciliatea. Inti Protociliatia menunjukkan kemiripan
struktur dan fungsi dengan Euciliatea, yaitu memiliki karakteristik inti yang
dimorfisme (macronucleus dan mikronukleus).
Subkelas 1. Protociliatia
Merupakan siliata opalinid, kecuali untuk beberapa spesies parasit pada
ikan dan ular dan dalam usus besar amfibi. Opalinid tidak memiliki sitostom,
meskipun bukan merupakan fitur eksklusif di antara siliata. Distribusi silia
seragam sehingga opalinid kadang-kadang diklasifikasikan menyerupai siliata
Holotrichous.
Pola siliari agak sederhana. Seperti pada Opalina obtrigonoidea, baris
dorsal mengikuti jalur sigmoid sedangkan baris ventral relatif lurus (Gambar 3.1,
A). Sepanjang permukaan anteroventral di O. obtrigonoidea, sejumlah silia
falcular muncul dari fibril falcular (Gambar 3.1, C, E). Fibril subpericular yang
memanjang sepanjang margin anteroventral dan kemudian melebur menjadi satu
fibril tunggal. Fibril berbentuk sabit dihubungkan dengan granula basal pertama di
setiap baris dari silia somatik (Gambar 3.1, E). Pada Opalina ranarum, tidak ada
hubungan antara fibril dan spherules endoplasma yang dapat dideteksi dalam
O.obtrigonoidea. Tidak adanya hubungan tersebut juga telah dilaporkan untuk
Cepedea metcalfi, Opalina coracoidea, dan O.ranarum.
Gambar 3.1
35
E. Distribusi geografis
Distribusi geografis Protociliatia yang menarik. Genus Protoopalina konon
merupakan genus yang paling primitif, terdistribusi sangat luas. Genus Zelleriella
terdapat di pusat Amerika, Amerika Selatan, dan selatan Amerika Utara. Cepedea,
meskipun tidak terwakili di Australia, dinyatakan secara luas terdistribusi,
sedangkan Genus Opalina belum ditemukan di Australia atau Amerika Selatan.
Subkelas 2. Euciliatia
Merupakan siliata khas dengan macronuclei dan micronuclei. Subdivisi,
ordo dan subordo sebagian besar didasarkan pada distribusi silia dan turunannya
dan pada diferensiasi struktur seperti di daerah peristomial. Subkelas ini dibagi
menjadi empat ordo, yaitu Holotrichida, Spirotrichida, Peritrichida, dan
Chonotrichida.
Membranelles zona adoral pada Holotrichida sangat kurang berkembang
begitu juga karakteristik dari peristome di Spirotrichida. Silia somatik merupakan
ciri khas, meskipun umumnya silia dibatasi untuk zona tertentu atau untuk satu
permukaan tubuh.
Spirotrichida menunjukkan perkembangan luas dari membranelles dan ciri
dalam kelompok tertentu telah sepenuhnya diganti silia sederhana. Sebuah zona
adoral dari membranelles muncul di kiri sitostom dan sepanjang anterior tubuh.
Kelompok ini secara keseluruhan menunjukkan kecenderungan kuat terhadap total
pengurangan area bersilia.
Epistom (daerah peristomial) pada Peritrichida umumnya merupakan
daerah diskoid yang dibatasi oleh dua atau lebih baris dari silia. Mayoritas adalah
sessile dan biasanya dilengkapi dengan tangkai.
Gonotrichida merupakan ektokomensal yang melekat pada host mereka
dengan disk basal atau tangkai pendek. Peristom di tiang distal, biasanya
dikelilingi oleh saluran atau kadang-kadang oleh dua saluran konsentris. Silia
dapat dibatasi ke peristom dan saluran dalam tahap dewasa. Reproduksi oleh
budding adalah karakteristik dari ordo ini.
36
ORDO 1. Holotrichida
Merupakan ordo yang besar, biasanya dianggap lebih primitif daripada
Euciliatia yang lainnya, menunjukkan diversifikasi besar daerah peristomial dan
dalam satu kelompok, sitostom telah menghilang. Spesialisasi seperti ini
memberikan dasar untuk membagi ke dalam subordo Holotrichida.
Subordo 1. Gymnostomina
Sitostom terbuka langsung di permukaan. Dalam banyak genus sitostom
terletak pada atau dekat anterior tubuh.
Subordo 2. Trichostomina
Sitostom biasanya terletak pada permukaan, biasanya dilengkapi dengan
satu atau lebih silia. Penggabungan silia peristomial ke dalam membran sederhana
atau membranelles, atau keduanya, ditemukan pada beberapa spesies.
Subordo 3. Hymenostmina
Cilia peristomial telah termodifikasi menjadi beberapa membran, mungkin
berasal filogenetis dari silia peristomial Trichostomina.
Subordo 4. Thigmotrichina
Bentuk yang paling karakteristik dari komensal ini adalah kelompok silia
thigmotactic yang terletak di anterior. Sitostom bergeser ke posisi pada atau dekat
ujung posterior tubuh. Pada beberapa Famili terdapat pengisap anterior, yang
merupakan suatu organel baru.
Subordo 5. Apostomina
Sitostom ventral ukurannya sangat sedikit, mungkin terbatas pada partikel
yang sangat kecil. Di bawah sitostom terdapat "roset" (Gambar. 3.2, A, F) dengan
fungsi yang belum pasti. Siliata somatik mencakup kurang dari 22 baris lengkap
silia. Siklus hidupnya cukup kompleks.
Gambar 3.2
37
Subordo 6. Astomina
Merupakan Holotrichs endoparasitik tanpa sebuah sitostom.Tubuh
seragam, bersilia, namun mungkin terdapat daerah bebas silia di ujung anterior.
Subfilum Ciliophora memiliki klas Ciliatea, dan Suctoria. Ciliatea
memiliki dua sub klas Protociliatea dan Euciliatia, ordo Holotrichida. Pada
makalah ini akan dibahas mengenai ordo Holotrichida. Ordo tersebut memiliki 6
sub ordo, yaitu Gymnostomina, Trichostomina, Hymenostmina, Thigmotrichina,
Apostomina, Astomina.
1. Subordo Gymnostomina
Subordo 1. Gymnostomina
Bentuk tubuh bervariasi. Berbentuk bulat telur, buah pir, gelendong, dan
umumnya yang berjenis panjang berbentuk ulat. Di beberapa genus, silia somatik
terbatas atau untuk satu permukaan tubuh. Sitostom biasanya terbuka di
permukaan.
Posisi sitostom bervariasi dalam Famili yang berbeda dan atas dasar ini,
Kahl telah membagi menjadi tiga subordo suku. Famili Actinobolinidae,
Butschliidae,
Colepidae,
Didiniidae,
Holophryidae,
Metacystidae,
dan
Spathidiidae merupakan kelompok yang memiliki sitostom anterior ("Familia
prostomata"). Kelompok kedua ("Familia hypostomata") memiliki sitostom yang
terletak
pada
permukaan
ventral.
Kelompok
ini
terdiri
dari
Famili
Chlamydodontidae, Dysteriidae, dan Nassulidae. Sitostom kelompok ketiga
("Familia pleurostomata") terletak pada margin atau lateral tubuh. Kelompok ini
terdiri dari Famili Amphibotrellidae, Amphileptidae, Loxodidae, dan Tracheliidae.
Famili 1. Actinobolonidae
Tentakel pada Dactylochlamys pisciformis adalah mirip dengan banyak
Suctorea (Gambar. 3.5, C). Tentakel pada Actinobolina vorax memiliki struktur
ramping di meridian siliari (Gbr. 3.2, J). Ujung dari setiap tentakel dikatakan
mengandung toxicyst. Berdasarkan struktur yang berbentuk seperti sungut, Kahl
telah menyarankan bahwa Actinobolinidae berhubungan dengan moyang
Holotrichs dari mana Suctorea itu berevolusi dan bahwa Dactylochlamys mungkin
38
merupakan tipe primitif dari Suctorea yang belum berkembang untuk
tahap
sessile.
Famili 2. Amphibotrellidae
Famili ini berisi genus R. amphibotrella dan L. grandori, ditandai dengan
lokasi sitostom dalam alur nonsilia anterior yang meluas hampir ke ujung
tubuh. Dekat ujung posterior tubuh juga ada alur bersilia.
Famili 3. Amphileptidae
Sitostom terletak di permukaan ventral dan biasanya dibatasi oleh zona
trichocyst. Biasanya terdapat dua atau lebih macronuclei. Kebiasaan karnivora
adalah karakteristik dari Famili ini, siliata lain dan rotifera menjadi mangsa umum
dari berbagai Amphileptidae.
FamilI 4. Butschliidae
Siliata ini terdapat di saluran pencernaan herbivora seperti kuda dan
unta. Tubuh kurang lebih bulat telur dengan sitostom biasanya di ujung
anterior. Sebuah anterior concretion-vakuola (Gambar 3.3, I) yang telah dianggap
sebagai statocyst dan memiliki satu atau lebih vakuola kontraktil. Memiliki
cyptopyge posterior yang khas. Silia dapat merata atau terbatas pada daerah
tertentu.
Gambar 3.3
39
Famili 5. Chlamydodontidae
Silia terbatas pada permukaan ventral (Gambar 3.4, D). Sebuah pita lurik
sempit melintang di daerah bersilia di Chlamydodon. Sitostom adalah anteroventral dan faring dikelilingi oleh keranjang faring (Gambar 3.4, J). Membran
adoral kadang-kadang ada namun kecil dan kurang berkembang. Tidak ada stylus
ventral bersifat protoplasma seperti yang ditemukan dalam Dyteriidae (Gambar
3.5, I). Chlamydodontidae biasanya memakan diatom dan alga lainnya,
phytoflagellates, dan bakteri.
Famili 6. Colepidae
Memiliki bentuk agak bulat dengan korteks yang terlindungi. Bentuk dan
susunan pelindung korteks berbeda pada banyak spesies (Gambar 3.4,
I). Pelindung dari colepshirtus mengandung kalsium karbonat dan ditutupi oleh
kulit tipis organik. Sebuah cincin trichites chyrcumpharyngeal (Gambar 3.4, B, G)
merupakan karakteristik dari Famili ini. Meskipun Colepidae kadang-kadang
memakan ganggang kecil, namun hewan ini karnivora.
Famili 7. Didiniidae
Merupakan siliata yang simetris radial terhadap sumbu longitudinal. Sitostom
anterior (Gambar 3.5, B, F) tidak dikelilingi oleh silia meskipun ada cincin
tentakel circumoral di mesodonium. Sebuah ciclet dari pectinellae (membranelles
ramping) biasanya terletak di atau dekat tepi tiang anterior. Sebuah pita dari
trichites circumpharyngeal adalah karakteristik Famili ini. Meskipun beberapa
spesies makan ganggang, mereka adalah karnivora. Didinium nasutum terkenal
karena kebiasaan yang menangkap dan mencerna siliata seperti Paramecium.
Gambar 3.4
Gambar 3.5
40
Famili 8. Dysteriidae
Hipostoma ini berbeda dari Chlamydodontidae dan Nassulidae stylus
ventral yang bersifat protoplasma (Gambar 3.5, I, K). Pada Dysteria, struktur ini
dapat menempel pada permukaan dan berfungsi sebagai jangkar sementara,
sedangkan stylus di Trochilioides mengeluarkan filamen ramping yang memiliki
fungsi yang sama. Permukaan dorsal tidak bersilia. Sebuah faring merupakan
karakteristik Famili ini. Dysteriidae, terutama siliata laut, sebagian besar
memakan diatom, ganggang lainnya, atau bakteri.
Famili 9. Holophrydae
Sitostom
terletak
di
atau
dekat
kutub
anterior
terbuka
dan
berbentuk. Terdapat zona circumpharyngeal dari trichites (Gambar 3.6,A), dan
baris adoral silia menyatu (syncilia) kadang-kadang tampak (Gambar 3.6,
F). Bentuk
tubuh
bulat
telur
atau
silinder. Kebiasaan
makan
sangat
bervariasi. Beberapa spesies karnivora rakus, mengejar dan menangkap siliata lain
atau rotifera. Beberapa biasanya memakan flagelata kecil, makanan lain yang
paling utama ialah
bakteri dan ganggang kecil, sedangkan spesies tertentu
memakan berbagai bakteri, diatom, dan nematoda kecil.
Famili 10. Loxodidae
Seperti dalam Amphileptidae, sitostom terletak di "ventral" margin tubuh
dan cenderung agak cekung. Permukaan yang tepat bersilia. Ganggang dan bakteri
merupakan makananya.
Famili 11. Metacystidae
Karakteristik dari Famili ini adalah sitostom terminal. Sitostom dapat
berbentuk bulat atau seperti celah, dan dalam spesies tertentu, membuka ke
rongga endoplasma ("receptable"). Makanan yang utama pada siliata ini adalah
bakteri. Lorica pseudochitinous adalah contoh spesies dalam Famili ini.
41
Famili 12. Nassulidae
Tubuh sepenuhnya bersilia. Keranjang faring merupakan ciri khas Famili
ini tetapi tidak ada stylus ventral seperti pada Dysteriidae. Kelompok siliata dalam
Famili ini memakan diatom dan alga lainnya.
Famili 13. Pycnothricidae
Siliata ini terdapat di sekum dan usus besar berbagai mamalia. Alur tubuh
sepenuhnya bersilia dan panjang biasanya menyebabkan sitostom yang mungkin
terletak dekat bagian tengah atau di ujung posterior tubuh. Dalam Pycnothrix
monocystoide (yang
mencapai panjang 2-3
mm),
tidak ada sitostom
tunggal. Sebuah lapisan tebal ektoplasma di bagian anterior tubuh adalah fitur
mencolok pada Gollinella, Nicollella, dan Pycnothrix.
Famili 14. Spathidiidae
Siliata Prostomatous ini memiliki celah pada sitostom (Gambar 3.10, B,
D). Dalam Legendrea, bagian posterior tubuh beruang sejumlah tentakel, masingmasing dilengkapi dengan trichocysts (Gambar 3.11, D). Spathidiidae merupakan
karnivora yang memakan siliata dan flagelata lainnya.
Famili 15. Tracheliidae
Sitostom sekitar melingkar, yang terletak agak jauh dari tiang anterior dan
pada akhir baris ventral trichocysts, dikelilingi oleh trichites atau trichocysts dan
kadang-kadang oleh keduanya. Tubuh benar-benar bersilia. Para Tracheliidae
terdapat di air tawar dan garam dan biasanya karnivora, memakan siliata lain dan
di flagelata.
Gambar 3.10
Gambar 3.11
42
Banyak terdapat di luar ujung posterior dari cytostome. Terdapat tonjolan,
jika ada, dapat dilindungi dengan trichities atau trichocysts. Dalam legendrea,
bagian posterior tubuh mempunyai sejumlah tentakel, masing-masing dilengkapi
dengan trichocysts (Gambar 3,11, D). tidak seperti kondisi biasa, cytostome di
teutophrys terletak di dasar ekstensi dari tubuh meskipun rekonsiliasi somatik
biasanya lengkap dan seragam, deretan dipipihkan "silia" bentuknya memanjang
sepanjang sisi cytostome di Spathidioides. Selain itu, sebuah kerugian silia pada
permukaan kiri telah terjadi di Homalozoon. Spathhidiidae merupakan pemakan
karnivora yang biasa memangsa ciliates dan flagelata.
Famili 15. Tracheliidae
Cytostome melingkar,terletak jauh dari anterior dari kutub anterior dan
pada akhir baris ventral trichocysts. Tubuh bersilia. Tracheliidae terdapat di air
tawar dan garam dan biasanya karnivora, memakan ciliates lain dan flagelata.
Subordo 2. Trichostomina
Cytostome biasanya terletak di dasar alur yang didefinisikan baik oral atau
pit, dinding yang dikenakan satu atau lebih medan padat silia Adoral. Bidangbidang seperti sering mengandung silia gratis. Namun, baik membranelles Adoral
dan membran bergelombang (atau mungkin "pseudo-membran") telah dilaporkan
di
genera
tertentu,
seperti
Woodruffia
dan
Colpoda.Dalam
beberapa
Trichostomina primitif, cytostome terletak hampir di kutub anterior. Lebih sering,
mulut di posterior pada permukaan ventral. Spiral torsi tubuh, cenderung
mempersulit pola silia peristomial, adalah karakteristik dari marga tertentu. Lima
belas Famili telah diakui.
Famili 2. Clathrostomidae
Peristome yang tumbuh, dangkal ventral dan normalnya tidak bersilia.
Dengan cytostome oval berbaring dibagian anterior. Tepi cytostome merupakan
sebuah pita yang membedakan dari sirkum fibril faring memperpanjang ke
endoplasm tersebut.
Famili 3. Colpodidae
Mulut, di paruh anterior permukaan luas, dapat berbentuk corong atau
segitiga, atau kadang-kadang memanjang. Dinding alur oral sering lebih tegak
43
lurus terhadap permukaan tubuh, sedangkan dinding kiri cenderung kemiringan
lebih bertahap. Baris somatik ciliary umumnya berbentuk rangkaian yang agak
konsentris di sekitar margin kanan peristome. sedangkan organisasi silia Adoral
bervariasi. Dalam Woodruffia metabolica, margin kiri dari alur oral mempunyai
ruang membran, masing-masing terdiri dari dua atau tiga silia yang menyatu,
sementara membran bergelombang halus memanjang sepanjang kanan. Biasanya
membran bergelombang dan memiliki struktur dalam, Adoral jenis ini mirip
dengan yang ditemukan dalam Heterotrchina. Fisi dalam kista reproduki adalah
khas, meskipun telah memungkinkan untuk mendapatkan fisi dalam tahap aktif di
bawah kondisi eksperimental Colpoda. Makanan biasanya berkisar dari ciliates
kecil untuk ganggang dan bakteri.
Famili 4.Conidiophryidae
Ektoparasit yang hidup menempel di rambut exoskeletal dari amphipod
dan Isopoda Crustacea. Pada dewasa, tahap non-bersilia dalam membran
dikeluarkan. Pada tahap migrasi akan berenang hingga tertusuk pada sebuah
rambut exoskeletal yang melewati faring. silia kemudian menghilang dan parasit
muda mengeluarkan sebuah membran di sekitar itu sendiri dan bagian distal
rambut atas. selama pertumbuhan, nutrisi ternyata dilengkapi oleh sekresi dari
rambut exoskeletal.
Famili 5. Cyathodiniidae
Kelompok ini, terdapat pada usus babi guinea, berisi beberapa jenis
cyanthodinium da Cunha. silia yang terbatas pada sekitar anterior tubuh.
peristome nonsilia adalah segitiga alur agak panjang. Dari deretan papila
sepanjang tepi kiri, trichites peristome berbentuk ramping ("endosprits" Lucas)
memperpanjang ke endoplasm tersebut. Eksternal, sebuah silia adoral muncul
dari papilla masing-masing.
Famili 6. Entorhipidiidae
Ciliates ini landak laut. Tubuh lebih rata dan ujung anterior membentuk
lobus frontal yang overhang yang cytostome. somatik yang rekonsiliasi selesai
dan pada dasarnya seragam.
44
Famili 7. Isotrichidae
Famili ini memiliki karakteristik dari rumen ungulata, meskipun spesies
Isotricha diketahui dari kecoa. Mulut adalah terminal atau subterminal dari faring
bersilia yang telah dilaporkan. striations longitudinal yang juga telah dijelaskan
dalam dinding faring. Sebuah cytopyge terletak di aboral dalam spesies tertentu.
Meskipun thecytostome biasanya dianggap anterior, beberapa spesies berenang
dengan thisend tubuh diarahkan posterior.
Famili 8. Marynidae
Ciliata soliter atau koloni, dengan lorica gelatin. Peristom yang sebagian
atau seluruhnya mengelilingi ujung bebas dari tubuh dan meluas ke posterior
untuk beberapa jarak pada permukaan ventral. organisme berenang dengan ujung
aboral.
Famili 9. Paraisotrichidae
Mulut subterminal, yang membuka hanya posterior vakuola. Rekonsiliasi
Thesomatic selesai dan, kecuali untuk seberkas anterior silia.
Famili 10. Parameciidae
Alur oral memanjang dari ujung anterior menuju tengah tubuh. silia
Adoral termasuk pada zona dorsal dibedakan dari silianya yang panjang
('membran quadripartite' dari von Gelei) dan penniculus, sebuah pita padat silia
yang meluas dalam spiral menuju cytostome.
Famili 11. Plagiopylidae
Merupakan dorso-ventral dari silia yang diratakan dengan alur peristomial
pusat yang terletak di bagian anterior tubuh dan meluas kurang lebih melintang
dari tepi kanan menuju atau melewati sagital. Sebuah pita nonsilia punggung
berbentuk lurik, strip menebal terdapat di Lechriopyla dan Plagiopyla. Fungsi
signifikan dari pita ini tidak diketahui. Rekonsiliasi somatik dinyatakan lengkap.
Sebuah cytoproct bersilia telah dilaporkan dalam Lechriopyla.
Famili 12. Spirozonidae
Famili termasuk Spirozona Kahl. sebuah band dari silia erat diatur
memanjang dari ujung posterior peristome posterior beruang seberkas bulu ekor.
45
Jika tidak, isuniform rekonsiliasi somatik. Spesies hanya dijelaskan terjadi di air
tawar dan memakan bakteri.
Famili 13. Trichopelmidae
Merupakan silia lateral dikompresi memiliki kulit tipis yang biasanya
menunjukkan beberapa tulang rusuk dengan alur membujur, yang terakhir
kadang-kadang terbatas ke permukaan kiri. Tepi dorsal berbentuk setengah
lingkaran atau bulan sabit halus secara garis besar. Mulut mungkin terletak dekat
anterior atau ujung posterior atau dekat equator. Hanya ada sedikit deretan silia
somatik. Trichopelmidae menyerupai Ctenostomina tetapi tidak terdapat
karakteriskik membran adoral dari heterotrichnya.
Famili 14. Trichospiridae.
Famili ini berisi genus Trichospira Roux. Sebuah kumpulan rapat diatur
silia, sebanding pada Spirozona, perpanjangan posterior dari peristom tetapi
berbentuk spiral ke kanan bukan ke kiri. Tubuhnya tidak runcing ke titik posterior
dan tidak terdapat bulu kaudal. Spesies yang diketahui terdapat pada air tawar.
Famili 15. Trimyemidae.
Hanya genus Trimyema Lackey (Sciadostoma Kahl) yang diketahui.
Kecuali bulu kaudal, silia somatik terbatas hanya tiga hingga empat baris pada
setengah anterior tubuh. Sitostomnya merupakan subterminal. Spesies yang telah
diketahui berasal dari air tawar dan air laut.
Subordo 3. Hymenostomina
Silia adoral berfusi ke dalam membran, jumlah, ukuran, dan susunan yang
bervariasi pada genus yang berbeda. Area peristomial juga memperlihatkan
beberapa variasi. Pada beberapa genus terdapat sejenis kantong oral yang tersusun
dari membran adoral dan membuka permukaan tubuh.
Pada genus lainnya,
siliatur adoral tumbuh di pada alur dengan kemungkinan menjadi panjang secara
wajar atau kemungkinan terjadinya pemanjangan sebagian tubuh.
Kahl membagi subordo ke dalam lima Famili. Cohnilembidae (Lembidae),
Frontoniidae, Ophtyoglenidae, Philasteridae, dan Pleuronematidae. Dari jumlah
tersebut, Frontoniidae menjadi kelompok heterogen yang akan menjadi kurang
jika Tetrahymena dan beberapa hubungan genus dihapus. Seperti perbaikan yang
46
telah dipengaruhi
oleh
Murgard
pdalam
mentransfer
silia
ke
Famili
Leucophyridae. Sejak Corliss menyimpulkan bahwa tidak ada sartupun spesies
Muller
dari Leucophra merupakan “congeneric” dengan beberapa spesies
kemudian disebut Leucophyrys dan Leuchophra harus menjadi genus “dubium”,
Leucophyrys harus diganti dengan genus Tetrahymenidae Furgason sebagai jenis
genus. Produk tersebut akan ditambah ke dalam enam Famili menjadi 5 Famili
Kahl yang asli, tetapi tanpa penguranan yang dibutuhkan lebih lanjut dalam
pembelajaran sisa Frontoniidae.
Famili 1. Cohnilembidae.
Rekonsiliasi somatic dari silia tersebut lengkap dan lebih seragam dan silia
adoral terdiri dari empat membran. Paroral, lateral, membran terbentang
sepanjang margin kanan dari pemanjangan peristom. Tiga membran adoral, yang
terletak ke kiri pada kantong oral Tetrahymenidae yang bergeser ke kanan pada
Anophrys seperti linear sisi padallel ke membran paroral. Tampaknya tidak ada
tiga baris dari akhir silia somatic pinggiran posterior pada “cavity” oral. Pada
stomatogenensis selama pembelahan Anophrys, organel adoral dari posterior
keturunan diambil dari granula basal dengan multiplikasi pada pusat membran
paroral. Tipe stomatogenesis ini berbeda dari Tetrahymenidae.
Famili 2. Frontoniidae.
Organel adoral bertanggung jawab terhadap banyak ketidakpastian dalam
hal komposisi generasi dari Frontoniidae. Investigasi selanjutnya menunjukkan
ketidakpastian tersebut dibenarkan.
Famili 3. Tetrahymenidae.
Siliatur adoral tersusun dari tiga membran, tiga adoral mengarah ke kiri
kantong oral dan membran paroral terbentang sepanjang pinggir kanan (Fig, 7.19,
C). Keistimewaan lainnya adanya satu atau lebih baris silia (“baris
stomatogenous) dengan akhir pada pinggir posterior kantong oral. Pada
stomatogenesis “branes” posterior keturunan berasal dari bidang dasar granula
yang berasal dari multiplikasi granula pada satu atau lebih baris stomatogenesis.
Spesies tertentu menunjukkan dua fase morfologi, makrostom dan
mikrostom. Dilengkapi dengan kantong oral yang besar adalah karnovora. Kedua
47
T. patula dan T. vorax terlihat dimorfik. Famili Tetrahymenidae sangat penting
bagi beberapa spesies, termasuk T. patula, T. pyriformis, T. vorax, telah dibentuk
pada kultur bakteri bebas dan digunakan pada investigasi fisiologi dan biokimia.
Lebih dari 20 strain T. pyriformis, sejarah yang terlacak oleh Corliss menjadi
peliharaan pada berbagai macam laboratorium.
Famili 4. Hysterocinetidae.
Silianya seragam dan padat, pada dorso-ventral silianya rata dengan
sitostom posterior. Mungkin hanya sebuah alur terminal transfersal, sepanjang
membran bergelombang diperpanjang ke dalam faring pendek. Karakteristiknya
pada “sucker” anterior nonsilia. Silia ini telah dilaporkan dari pencernaan
beberapa siput dan oligochaetes air tawar.
Famili 5. Ophryoglenidae.
Terdapat siliata pada peristome, invaginasi pada dinding tubuh, dan faring
terbuka ke dalam vestibula. Vestibula relatif dalam pada Ophryoglena, sedikit
pada Protophryoglena dan banyak mengalami penurunan pada Ichthyophthirius.
Spiral punggung memanjang dari pembukaan vestibula hingga faring pada
Ophryoglena. Karakteristiknya adalah pengaturan tetrahymenal pada membran
oral. Pada stomatogenesis, membran oral berasal dari basal granula biasanya
tersusun dari sejumlah pigmen.
Siklus hidup mengikuti pola umum. Reproduksi terjadi antar kista. Silia
muda (tomites) berkembang menjadi fase tropik (theronte) yang hidup bebas,
tumbuh menjadi besar dengan pelan pada fase melayang dengan banyak cadangan
makanan. Pada fase matang ketika encysy persiapan untuk reproduksi. Pada
Ichthyophthiris membelah memproduksi banyak silia kecil yang diatur bebas
dengan perkembangan mulut yang tidak sempurna. Pada tahap aktif berenangrenang sampai mati kelapadan atau pada ikan yang cocok. Setelah penetrasi,
padasit muda berkembang menjadi cytostom fungsional, menjadi makanan
sebagian pada fragmen sel epitel dan tumbuh hingga diameter 100-1000µ. Silia
muda kemudian keluar dari inang dan encyst.
Famili 6. Philasteridae.
Anggota Famili ini pemanjangan silia dengan panjang dan kira-kira
berbentuk segitiga beralur pada peristom. Tiga membran adoral digantikan oleh
48
tiga silia dasar. Membran silia tetap melakukan serupa, memperluas bagian
wilayah sepanjang margin kanan pada peristom. Stomatogenesis termasuk
multiplikasi dari granula basal posterior akhir pada membran paroral.
Famili 7. Pleuromatidae.
Membran paroral lebih besar dan meluas sepanjang margin posterior
peristom. Silia adora saat berhenti perkembangannya kurang seragam dan
ditunjukkan dengan membran single pada kiri peristom atau dasar silia pada
wilayah ini. Pada Balantiophorus membran berkelanjutan meluas sepanjang
kanan, posterior, margin kiri pada peristom. Ketika meluas secara penuh,
membran membentuk seperti struktur penutup persitom kecuali anterior akhir.
Beberapa Pleuronematidae , sejumlah silia dorso-lateral “thigmotatic”. Satu atau
lebih kaudal terdakang terlihat.
Subordo 4. Thigmothrichina
Silia ini terjadi sebagian besar pada rongga mantel atau insang dan palp
moluska bovalvia, juga ditemukan pada pernafasan keong dan tentakel
Phoronopsis. Dasar anterior pada silia “thicmotactic” merupakan karakteristik
umum. Sitostom dan siliatur adoral terlihat pada keseluruhan, tidak ada pada
sebagian posterior tubuh. Pada beberapa spesies sitostom tidak ada pada posterior
kutub. Pada spesiasi genus kehilangan sitostom dan berkembang pada tentakel
suktorial anterior. Kecuali tipe primitive, terdapat reduksi silia somatikdan
mencapai klimaks pada genus dengan hanya menahan silia “thigmotatic”. Chatton
and
Lwoff
membagi
6
Famili.
Conchophthiriidae,
Hemispeiridae,
Thigmophryidae, Acistrocomidae, Hypocomidae, Sphenophryidae.
Famili 1. Ancistrocomidae.
Siliata menjauh, periform, atau beberapa silinder. Secara khas dengan
banyak atau sedikit titik kutub anterior. Penempelan diatur silia “thigmotactic”,
tidak sepanjang silia somatic, terbatas pada beberapa dasar anterior pendek.
Tubuh secara ekstensif terdapat silia pada beberapa genus. Pada genus lainnya
silia terbatas pada dasar “thigmotactic”. Karakteritiknya, yaitu pada tentakel
49
anterior
suktorial
berkelanjutan
dengan
kanal
internal.
Tentakel
ini
memungkinkan organisme menjadi dempet pada sel epitel dan pencernaan.
Famili 2. Conchophthiriidae.
Terdapat sitostom fungsional pada sebagian posterior tubuh. Silia
“thigmotactic” terlihat pada bagian anterior baris somatik daripada beberapa
bentukan dasar. Tubuhnya secara lateral dikompres dan sitostomnya hilang pada
permukaan sempit ventral.
Famili 3. Hemispeiridae.
Memiliki tipe silia lebih padat pada posterior dan subterminal sitostom.
Siliatur adoral terlihat dua komponen khas. Pada kanan baris single, baris 1,
sedikit kurva hingga hamper keseluruhan kiri dan kemudian secara tajam. Di
baris kiri 1 baris ganda dari silia (baris B) mengikuti banyak alur yang sama tetapi
tidak memperpanjang sejauh posterioly. Selain itu, baris pendek silia terletak di
dekat ujung anterior B baris dalam beberapa spesies dan ada juga deretan silia di
faring (30). Pola ini bervariasi secara detil dalam spesies yang berbeda. Silia
thigmotatic dari beberapa dari genus hanyalah silia anterior di deretan yang
normal. ketidakmampuan silia somatik posterior telah menghilang, hanya
menyisakan thigmotatic silia.
Famili 4. Hypocomidae.
Parasit ini berbentuk bulat telur untuk di mana silia terpapar pada bagian
permukaan dorsal. Terdapat sebuah tentakel pada bagian dorsal anterior atau
antero suctorial dan terdapat sebuah cystostome. Namun, ada yang menarik
menurut dugaan yaitu antero-lateral silia yang vestigial.
Famili 5. Sphenophryidae.
Tahap dewasa pada ciliata ini tidak bersilia meskipun dalam bereproduksi
"embyo" dan mengembangkan silia dan membentuk sebuah Ancistrcomidae.
tentakel suctorial umumnya lebih pendek dibandingkan dengan Ancistrocomidae
dan Hypocomidae dan cenderung berbentuk seperti corong.
50
Gambar 3.24
Keterangan : A,B, Hemispeire asteriasi C. Sphenophyra dosiniae D,E. Gargarius
gargarius F. Cheissinia (Tiarella) baicalensis (Cheiisen) G. Ancistrum pernix H.
Hypocoma parasitica I,J. Boveria teredini
Famili 6. Thigmophyridae.
Bentuk ciliata ini pada dasarnya sama dengan pada ciliate lainnya. Bidang
thigmotactic menggambarkan ciliata yang pendek, dengan teliti silia ini diatur
dalam berbagai deretan somatik. Conchophyllum caryoclada (Kidder) Raabe
menunjukkan macronucleus bercabang dan tidak terjadi seperti biasanya.
Subordo 5. Apostomina.
Kelompok ciliata ini sering ditemukan, untuk urutan
dari ciliates ini
dimulai dengan cystome ventral yang kecil dan kemungkinan terdapat sedikit
cairan atau partikel. Sebuah bentuk yang khas pada ciliate yaitu “roset” adalah
suatu karakteristik dari permukaan ventral. Dalam Foettingeria cystostome
terletak pada suatu alur akhir, di periode berikutnya membentuk sebuah roset, dan
membuka ke sebuah kanal yang mengarah ke dalam, di sepanjang dinding yang
cekung (thypolose) dari bentuk yang rosetakan membentuk “kantong oral”.
Bentukan Roset mengandung 8-10 sekat vertikal yang bergabung dengan dinding
luar ke “thyplosole”, dan pada dasar thyplosole ini dilengkapi dengan cincin silia.
De Morgan tidak pernah melihat gerakan membentuk roset dalam siklus hidup F.
Actinarium dan tidak ada petunjuk yang diperoleh. Kurang dari 22 deretan
lengkap silia somatik tetapi memiliki jumlah yang tepat dan kelompok ciliata
yang menarik dan bervariasi dari genus yang berbeda.
51
Sebuah siklus hidup yang kompleks dengan suatu karakteristik. Pada tahap
pertumbuhan
atau “trophonte”, makanan ini akan dicerna dan terakumulasi
selama pertumbuhan tetapi tidak ada reproduksi yang terjadi. Dewasa ini yang
“trophonte” berhenti melakukan aktivitas makan, dan dengan atau tanpa
encysment pada spesies yang berbeda, berubah menjadi sebuah “protomonte”.
Jika yang “protomonte”. Adalah tahap encysted, silia kemudian dibuang.
“protomonte” berkembang menjadi “tomonte” di mana makanan diubah menjadi
sebuah akumulasi yang kemudian cadangan makanan ini disimpan. Kemudian
berulang terjadi fisi untuk menghasilkan “protomites” (Gambar 3.25 M), yang
masing-masing menjadi aktif berenang “tomite” (Gambar 3.25, E, G, I). Namun,
Chromidina elegans telah dilaporkan dari suatu Cephalopoda yang bentuknya
mirip ginjal. Setelah lengkap, “tomite” berkembang menjadi kista, atau
“phoronte” (Gambar 3.25 K). Ketika hostnya ditelan oleh sebuah coelenterate atau
ctenophore, atau ketika host mencair, fase muda “trophonte” muncul dari kista.
Tahap vegetatif (trophotomonte) dari chromidia elegans biasa dalam hal itu dapat
mengalami fisi untuk membentuk rantai yang mirip dengan yang dihasilkan oleh
Astomina
Gambar 3.25
Keterangan: A-D. Foettingeria actinarium yang berbentuk roset, A-C.
memperlihatkan sekat dan silia, E. fase ciliate muda (tomite) dari Spirophyra
subparasitica. F,G. Foettingeria actinarium, H,I. Chromidina elegans
(Foettinger), J. Synophyra hypertropica “trophonto”, K. fase sessil (“phoronte”)
dari Foettingeria actinarium, L. Gymnodioides inkystans, M. kista reproduktif
dari Spirophyra subparasitica
52
Subordo 6. Astomina.
Ciliate ini adalah sebuah parasit tanpa cytostome. Tubuh bersilia sama
tetapi kadang-kadang ada yang telanjang kecil didaerah kutub anterior. Panjang
rata-rata adalah untuk golongan Astomina ini, kemungkinan 200-500μ, tapi
spesies seperti Haptophyra gigantean, H. Michiganesis dan Mesnilella 53adiate
yang panjangnya mencapai 1,5-2,0 mm. Pada bagian korteks ini menunjukkan
spesialisasi yang kecil, meskipun berkisar dari zona yang sangat tipis dengan
tebalnya lapisan 1,0-2.0μ
yang terdapat dalam beberapa coelozoic untuk di
parasit usus tertentu.
Sejumlah Astomina telah mengembangkan bagian organelnya. Struktur
seperti ini biasanya terletak dekat kutub anterior yang sering terjadi kontak dengan
epitel dari inangnya. Pengisap antero ventral (Gambar 3.26 J, K) adalah
karakteristik dari spesies yang terletak diusus tertentu. Dalam berbagai Astomina
lainnya bagian anterior dilengkapi seperti duri atau spikula (Gambar 3,26 N, P:
27, BK: 28, C, G). Di samping itu, fibril kerangka fungsinya yang tidak diketahui
dan dapat memperpanjang untuk beberapa jarak dekat permukaan tubuh (Gambar
3. 27 H, J) dan kadang-kadang melewati dari bagian korteks ke endoplasm.
Vakuola kontraktil kadang-kadang memiliki sebuah kanal kontraktil yang
umumnya terdapat seperti pada Haptophyra (Gambar 3.26 J). Mungkin ada satu
vacuola kontraktil atau di daerah lain. Banyak vakuola diatur dalam satu atau
lebih urutan yang membujur (Gambar 3.26, L, O).
Gambar 3.26
Keterangan: A-F. formasi dari rantai Astomina: tipe Haptophyra (A-C), tipe
Radiophyra (D-F). G,H. Butschliella nasuta Rossolimo, I. Aboplophyra gammari,
J,K. Haprophrya michiganensis, L. Perseia dogieli Rossolimo, M,N.
Bucheneriella criodrili, O,P. Metaradiophyra asymmetrica
53
Sedikit yang diketahui tentang siklus hidup Astomina. Sebagian besar
spesies yang diketahui dari oligochaetes: beberapa dari krustasea amphipod yang
berasal dari saluran pencernaan turbellaria dan amfibi. Reproduksi kemungkinan
melibatkan yang khas yaitu pembelahan biner, pembelahan di mana terdapat satu
organisme yang lebih sedikit dan lebih kecil dari yang lainnya atau pembelahan
yang berturut-turut yang dapat menyebabkan produksi suatu rantai (gambar 7. 26
A, F). di beberapa rantai baik anterior (primite) dan organisme (satelit) posterior
mengalami pembelahan yang diulang; ketidakmampuan pada
primata
menghasilkan beberapa satelit tanpa mengalami reproduksi dalam suatu ukuran.
Famili di bawah ini dijelaskan untuk mewakili lima dari enam yang diakui oleh
Cheissin.
Famili 1. Anoplopharyidae.
Memiliki tubuh yang kemungkinan bulat telur memanjang dan silia
tersebut diatur dalam deretan yang membujur. Sebuah alat pengisap yang kurang
baik tumbuh. Mungkin ada satu, dua, atau vakuola kontraktil lebih, atau kadangkadang tidak ada.
Famili 2. Haptophyiridae.
Terdapat sebuah kanal kontraktil panjang, karakteristiknya bukan vakuola
kontraktil yang terpisah. Alat pengisap antero-ventral terdapat dalam beberapa
spesies. Spikula atau kait mungkin ada atau mungkin juga tidak terdapat spikula
atau kait yang biasanya terletak di akhir anterior.
Famili 3. Hoplitophyridae. Astomina ini dilengkapi dengan alat seperti pegangan,
spikula membentuk longitudinal atau memiliki kedua jenis struktur tersebut.
Mungkin terdapat beberapa vakuola kontraktil.
54
Gambar 3.27
Keterangan: A,B. kait apparatus dari Radiophyra lumbrici, D,E. Protoradiophyra
fissispiculata, F,G. Mrasekiella intermedia, H-J. Masnilella multispiculata, K.
ujung anterior dari Hoplitophyra secans
Famili 4. Intoshellinidae.
Ini adalah ciliata yang bentuknya memanjang dengan deratan longitudinal
atau deretan spiral dari silia. Terdapat alat pengait yang erat dalam bentuk duri
atau bergigi, area anterior non-bersilia. Sejumlah vakuola kontraktil yang disusun
dalam satu atau dua deretan membujur atau yang didistribusikan. Rantai biasanya
membentuk suatu formasi.
Famili 5. Maupasellidae.
Sedikit yang diketahui tentang kelompok ini yang hanya mengandung dua
dari genus, Mupasella Cepede (Gambar 3. 28, F, G) dan Schulzellina Cepede.
Ciliates ini serupa dengan Hoplitophyiridae dan Cheissin telah menyarankan
bahwa dilakukan penyelidikan yang lebih lanjut yang mungkin membenarkan dan
menggabungkan dua Famili tersebut. Selain lima Famili yang dijelaskan di atas,
Cheissin termasuk Chromidinidae, yang mengandung genus Chromidina Gonder
(= Opalinopsis Foettinger.). chatton dan Lwoff, di sisi lain, menyimpulkan bahwa
Chromidina termasuk dalam Apostomina.
55
Gambar 3.28
Keterangan: A-C. Intoshellina poljansky, D,E. Monodontophyra kijenskiji, F,G.
Maupesella criodrili
F. Ciri masing-masing ordo, sub ordo, dan famili dalam phylum Ciliophora
1. Ordo Spirotrichida
Karakteristik yang paling menonjol adalah membran yang ada di daerah
mulut, bagian dasar yang sempit yang biasanya terletak di kanan atau sudut miring
terhadap sumbu yang panjang pada daerah mulut. Rangkaian membran meluas ke
depan dari tepi kiri cytostome, dan dalam genus tertentu, mungkin berubah bagian
dorsal pada kutub anterior dan meluas ke kanan untuk beberapa jarak sepanjang
permukaan antero-dorsal. Pelat basal membran biasanya terdiri dari dua baris
butiran dasar/basal, walaupun tiga baris (jarang, empat) mungkin tampak.
Kelompok dapat dibagi menjadi enam sub ordo (Hall, 1961).
a. Sub ordo 1. Heterotrichina
Somatik ciliation biasanya lengkap. Namun, permukaan dorsal mungkin
jarang bersilia dalam beberapa famili dan menunjukkan penurunan lebih luas
dari rekonsiliasi dalam kasus luar biasa. Peristome, biasanya memanjang dan
cukup sempit, menuju daerah mulut dari membran sepanjang dinding kiri.
56
Selain itu, membran yang bergelombang sering meluas untuk beberapa jarak di
sepanjang tepi kanan.
b. Sub ordo 2. Oligotrichina
Meskipun membran pada mulut berkembang dengan baik, terdapat
pengurangan yang ditandai dengan rekonsiliasi somatik dan bidang
peristomial, sekitar daerah mulut meluas, bebas dari silia. Membran yang
bergelombang terletak di tepi kanan area mulut dalam genus tertentu.
c. Sub ordo 3. Tintinnina
Siliata ini kadang-kadang dikelompokkan dengan Oligotrichina, ciri
khasnya berbentuk kerucut dengan lorica. Daerah mulut dari membran
mengikuti bagian spiral dan kutub mulut yang rata/ datar.
d. Sub ordo 4. Entodiniomorphina
Kelompok ini, kadang-kadang ditempatkan dalam Oligotrichina, termasuk
parasit dari rumen dan usus herbivora. Rekonsiliasi mungkin terbatas pada
daerah mulut atau tiga, mungkin salah satu atau lebih pita tambahan atau
kelompok membran.
e. Sub ordo 5. Hypotrichina
Silia somatik digantikan dengan cirri yang umumnya didistribusikan dalam
bidang tertentu dan terbatas utamanya pada permukaan ventra
f. Sub ordo 6. Ctenostomina
Bagian samping dimampatkan, siliata berbentuk baji dengan kulit tipis
kaku dihiasi dengan rusuk yang membujur. Tubuhnya jarang memiliki silia,
dan peristome merupakan kantong yang berisi delapan membran pada daerah
mulut.
Sub ordo 1. Heterotrichina
Mayoritas rekonsiliasi somatik praktis lengkap, siliata ini biasanya
dianggap Spirotrichida paling primitif. Namun, ada kecenderungan menuju
pengurangan silia bagian dorsal pada beberapa genus, dan Peritromidae
merupakan ciliata yang hanya memiliki silia pada permukaan ventral. Selain
membran pada daerah mulut sebelah kiri, sering terdapat membran
bergelombang di sebelah kanan peristome. Membran ini kadang-kadang
57
digantikan oleh sepasang baris dari silia padat. Sebanyak tigabelas famili
telah diakui.
Family 1. Balantidiidae
Famili Balantidiidae meliputi Balantidium Claparede and Lachmann
(Gambar 3.29, A-E), diwakili oleh parasit pada saluran pencernaan pada
vertebrata dan invertebrata. Rekonsiliasi somatik telah selesai dan silia
tersebut diatur dalam baris membujur/ longitudinal. Peristome merupakan
kantong dengan lubang berbentuk segitiga, palung dengan pita pendek di
dalam mulut dan membran tidak mudah dikenali dari luar. Banyak fibril
memanjang ke dalam endoplasma dari granula basal pada cilia dan
membran.
Gambar 3.29. A. Balantidium praenucleatum (irisan membujur) (setelah K dan
M). B, C, irisan melintang Balantidium coli; (B) daerah anterior
menunjukkan peristome, (C) irisan posterior tampak faring. D,E,
Balantidium sushilii: (D) irisan membujur tampak peristome, (E)
peristome di dekat ujung anterior (setelah R). F-H, irisan melintang
Bursaria truncatella O. F. M: (F)bagian anterior dalam peristome, (G)
dekat ujung posterior peristome, (H) dan pada permukaan cytostome. I.
Bursaria truncatella, tampak ventral. J. Bursaridium pseudobursaria
tampak ventral. K. Chattonidium setense, membran dan membran
bergelombang, makronukleus, rongga postero-axial (cytoproct). L.
Chattonidium setense, tampak polar, membran, dasar membran (setelah VB.).
Family 2. Bursariidae
Karakteristik yang paling menonjol adalah peristome berbentuk corong
besar, menutupi seluruh bagian ventral atau sebagian besar panjangnya
(Gambar 3.29, F-I). Penutupan daerah ventral ini mungkinhasil dari
58
pertumbuhan yang terlalu cepat/ berlebih dari “oral lip”, piringan seperti
memanjang dari dinding tubuh yang memanjang dari tepi kanan peristome
pada berbagai Heterotrichina. Dalam Bursariidae, pemanjangan ini
mungkin telah menyatu dengan tepi kanan peristome. Membran
bergelombang telah menhilang pada sebagian besar spesies. Tiga genus
menunjuk pada famili: Bursaria Muller (Gambar.7.29, F-I), Burasidium
lauterborn (Gambar 3.29, J), dan Thylacidium Schwiakoff .
Family 3. Chattonidiidae
Siliata ini menunjukkan kemiripan ke Oligotrichina dalam peristome yang
tergeser ke kutub anterior. Membran membentuk spiral yang hampir
lengkap di sekitar tepi dari corong peristomial, pada bagian dasar dimana
terletak cytostome (Gambar 3.29. K). Dalam cincin membran, membran
bergelombang memanjang untuk beberapa jarak di sekitar peristome.
Rekonsiliasi somatik adalah sama/ seragam. Genus Chattinidium
Villeneuve (Gambar 3.29, K,L), adalah satu-satunya yang ditempatkan
dalam famili.
Family 4. Clevelandellidae
Merupakan ciliata heterotrichs yang meruncing ke arah kutub anterior
(aboral). Daerah membran memanjang menjadi peristome berbentuk
corong menuju faring (Gambar 3.30, G,N). Famili ini meliputi dua genus,
keduanya diwakili oleh spesies dalam saluran pencernaan kecoak
(Panesthia): Clevelandella Kidder (112; Gambar 3.30, E,f,n) and
Paraclevelandia Kidder (Gambar 3.30, G).
Family 5. Condylostomidae
Peristome yang luas berbatasan di sebelah kiri daerah adoral membran
(Gambar 3.30, D). di sebelah kanan, membran bergelombang yang
panjang muncul dari alur yang tersembunyi oleh ventral ledge (“oral lip”).
ventral ledge ini diperluas ke kiri dalam beberapa spesies untuk
membentuk bagian dasar dari rongga peristomial. Pada permukaan antero
59
ventral, kadang-kadang terdapat penggantian progresif dari sebuah silia
sederhana oleh kelompok silia yang berfusi, mencapai puncak pada daerah
cirri pada tepi kanan peristome (Gambar 3.30, C). Genus Condylostoma
Bory (Gambar 3.30, A,C,D) adalah satu-satunya yang ditempatkan dalam
famili.
Gambar 3.30 A. Irisan melintang Condylostoma vorax dengan membran
bergelombang dan membran; B. Lorica Parafolliculina hirundo; C. Daerah
antero-ventral Condylostoma arenarium menunjukkan membran
bergelombang dan transisi dari silia somatik menjadi cirri. D.
Condylostoma vorax tampak ventral. E, F. irisan melintang Clevelandella
elongata; (E) faring, (F) peristome. G. Paraclevelandia brevis tampak
ventral; karyophore merapat dengan makronukleus anterior. H. Lorica
Microfolliculina limnoriae. I. Migrasi larva Folliculina aculeata. J.
Folliculinopsis
producta spesimen panjang menunjukkan ikatan
makronuklear. K. Lorica Folliculina viridis. L. tampak polar dari
peristome Folliculinopsis producta, menunjukkan dasar membran adoral
dan baris silia somatik. M. Lorica Pseudofolliculina arctica. N.
Clevelandella elongata tampak ventral.
Family 6. Folliculinidae
Famili ini distribusinya luas pada siliata laut yang hidup menempel pada
berbagai tanaman dan hewan.tubuhnya ditutupi dalam lorica yang relatif
tipis “pseudochitinous”. Pada kutub oral, tubuh meluas ke dalam dua lobus
yang aktif melintasi daerah spiral dari membran yang meluas ke sitostome
(Gambar 3.30, L). Tubuh berbentuk sama/ seragam dengan silia. Setelah
menyelesaikan fisi Folliculina, anak daun anterior lorica seperti larva yang
60
berenang bebas (Gambar 3.30, I), setelah periode migrasi yang singkat,
menjadi terikat dan mengeluarkan lorica.
Genus yang dibedakan oleh perbedaan dalam bentuk umum dari lorica
telah diakui: Folliculina Lamarck (Gambar 3.30, K), Folliculinopsis
Faure-Fremiet (Gambar 3.30, J,L), Metafolliculina Dons, Microfolliculina
Dons (104; Gambar 3.30,H), Parrafolliculina Dons (Gambar 3.30, B),
Pebrilla Giard, and Pseudofolliculina Dons (Gambar 3.30, M).
Family 7. Lichnophoridae
Kedua ujung dari tubuh memanjang discoidal, saat willayah tengah
mengerut. Daerah membran mengelilingi sebagian besar antero-ventral,
atau mulut, disk dan memanjang ke dalam bagian peristome tersebut. Disk
posterior (disk basal) dikelilingi oleh beberapa membran bergelombang
konsentris, dan hanya anterior ini yang mengarah fleksibel, atau velum
(bagian belakang langit-langit). Disk basal berbentuk seperti cangkir dan
berfungsi sebagai pelekat pada host. Dengan satu atau dua pengecualian,
Lichnophoridae adalah ektocommensal laut. Hanya satu genus yang
diketahui, Lichnophora Claparede (Gambar 3.31, J)
Gambar 3.31. A-C irisan melintang Metopussigmoides tampak bagian anterior peristome dan
permukaan posterior, menunjukkan beberapa membran (A, B) dan membran
bergelombang. D, E. Peritromus kahli: tampak ventral. F. Plagiotoma lumbrici,
menunjukkan adoral ciliation, silia somatik, dan vakuola kontraktil. G. Caenomorpha
medusula, menunjukkan pola silia dan dasar membran. H. Spirorhynchus verrucosa. I.
Metopus mathiastampak ventral. J. Lichnophora macfarlandi, menunjukkan basal dan
peristomial disk, adoral ciliation, pemanjangan membran lateral ke arah cytostome.
61
Family 8. Metopidae
Banyak siliata heterotrichs yang tidak seragam/ sama dimana peristome
cenderung membelok ke posterior kanan. Daerah membran lebih lurus
pada Metopidae primitif tetapi torsi spiral mungkin ditandai, seperti pada
Caenomorpha (Gambar 3.31, G). Membran bergelombang relatif pendek
dan sering melebar di sepanjang tepi kanan peristome. Trichocysts
mungkin tampak, kadang-kadang mendasari daerah pellicular atau pita
yang memisahkan baris silia.
Metopidae, diwakili di air tawar dan air asin, meliputi genus berikut:
Bryometopus Kahl, Caenomorpha Perty (Gambar 3.31, G), Copemetopus
Villeneuve Brachon, Ludio Penard , Metopus Claparede and Lachmann
(Gambar 3.31, A-C,I), Palmarium Gajevskaia, Spirorhynchus da Cunha
(Gambar 3.31, H), Trochella Penard, dan Tropidoatractus Levander.
Family 9. Peritromidae
Siliata laut yang menyerupai hypotrichs dalam tubuh dorsoventral mereka
yang rata dan pengurangan silia pada permukaan ventral. Permukaan
dorsal kadang terdapat bulu-bulu yang disebut trichocysts mucilagi. Pita
membran meluas di ujung anterior atau permukaan antero ventral dan
kemudian posterior sepanjang tepi ventral kiri menuju sitostome, biasanya
dekat dengan bagian tengah tubuh. Hanya dua genus yang ditempatkan
dalam famili: Pediostomum Kahl (104) dan Peritromus Stein (Gambar
3.31, D,E).
Family 10. Plagiotomidae
Kelompok ini mencakup parasit dari oligochaeta dan invertebrata yang
lain dan berbagai vertebrata. Tubuh padat bersilia, pita membran
berkembang baik (Gambar 3.32, A,B,L,M), dan membran bergelombang
terletak di tepi kanan peristome (Gambar 3.32, L). Siliata cytoproct
dideskripsikan dalam Nyctotherus (Gambar 3.32, C).
62
Tiga genus dirujuk dalam famili ini. Nyctotherus Leidy (Gambar 3.32, AC, F-H, L, M) diwakili oleh parasit pada usus vertebrata dan invertebrata.
Paranyctotherus Sandon (Gambar 3.32, I-K), didirikan untuk siliata dari
Afrika Selatan yang mirip dengan Nyctotherus tetapi menunjukkan deretan
membran di sepanjang tepi kanan peristome. Plagiotoma Dujardin
(Gambar 3.31, F) meliputi parasit pada rongga selom cacing.
Family 11.Reichenowellidae
Famili didirikan oleh Kahl untuk Reichenowella Kahl (fig. 7.32, E) dan
Balantidioides Penard (Gambar 3.32, D). siliata ini disebut berbeda dari
Heterotrichina lain dalam adanya bentukan seperti celah pada mulut,
biasanya tertutup dan tidak mudah terdeteksi, dan kekurangannya dari
lubang mulut. Terdapat pita membran, tetapi tidak ada membran
bergelombang.
Gambar 3.32. A-C. Nyctotherus kyphodes: A. Spesimen menunjukkan nukleus,
karyophore, vakuola kontraktil, cytoproct. B. Irisan melintang
menunjukkan peristome dan nukleus. C. Silia cytoproct dimana vakuola
kontraktilnya kososng. D. Balantidioides muscicola. E. Reichenowella
nigricans, menunjukkan membran adoral tetapi tanpa membran
bergelombang atau faring yang jelas. F-H. irisan melintang Nyctotherus
macropharingeus tampak anterior, tengah, dan daerah posterior dari
peristome. I-K. Paravyctotherus kirbhy: tampak dari sebelah kiri. (I)
menunjukkan fibril endoplasmik, makronukleus, dan peristome; (J) daerah
antero-ventral; (K) menunjukkan peristomial siliature dan fibril
endoplasmik. L, M. Irisan melintang Nyctotherus cordiformis. (L)
menunjukkan membran, membran bergelombang, vakuola kontraktil dan
makronukleus; (M) menunjukkan nukleus, karyophore, peristome, dan
cytostome.
63
Family 12. Spirostomidae
Beberapa Spirostomidae memanjang, dengan tubuh lebih atau kurang
kontraktil; yang lain adalah bagian dorsoventralnya rata sampai batas
tertentu. Pita membran yang panjang (Gambar 3.33, L, M), atau baris
ganda silia yang homolog meluas ke cytostome. Membran bergelombang
kadang-kadang cukup pendek, atau baris yang sesuai atau silia memanjang
di sepanjang tepi kanan peristome. Peristome ini mungkin agak lurus atau
dapat menunjukkan beberapa tingkat torsi spiral. Setidaknya dalam spesies
tertentu, pita dari trichocysts digantikan dengan deretan silia.
Genus berikut ditempatkan dalam famili: Blepharisma Perty (Gambar
3.33, G,I), Gruberia Kahl (Gambar 3.33, F), Paraolepharisma Kahl
(Gambar 3.33, L), Phacodinium Prowaze, Protocrucia da Cunha (Gambar
3.33, H), Pseudoblepharisma Kahl, Spirostomina
Gruber, and
Spirostomum Ehrbg. (Gambar 3.33, A-C, M).
Gambar 3. 33. A-C irisan melintang Spirostomum ambiguum tampak anterior dan daerah tengah
peristome dan dekat permukaan sytostome. D. Stentor felici menunjukkan rantai
makronuklear, membran, pola silia somatik. E. Fabrea salina; dasar membran,
peristomial striation, pola silia somatik. F. Gruberia calkinsi. G. Blepharisma hyalinum.
H. Protocrucia tuzeti. I. Blepharisma lateritium. J. Stentor auriculatus. K.
Climacostomum virens dengan peristome yang luas. L. Parablepharisma bacteriophora.
M. Spirostomum teres menunjukkan membran dan pola silia somatik.
Family 13. Stentoridae
Daerah membran cenderung meluas di sekeliling kutub anterior tubuh, dan
dalam beberapa kasus peristome itu sendiri telah bergeser ke kutub
(Gambar 3.33, D, J, K). Membran bergelombang telah menghilang.
64
Rekonsiliasi somatik relatif sama dengan silia diatur secara longitudinal
atau sedikit baris spiral.
Stentoridae, diwakili dalam air tawar dan air asin, termasuk dalam genus
berikut: Climacostomum Stein (Gambar 3.33, K), Fabrea Hennegui
(Gambar 3.33, E), dan Stentor Oken (Gambar 3.33, D, J).
Subordo 2 : Oligotrichina
Subordo ini ditandai dengan silia di bagian somatik mulai mereduksi dan
menghilang. Zona membran biasanya dibedakan ke dalam ikatan pendek bagian
oral dan ikatan spiral bagian anterior, dimana memiliki peran lebih dalam
pergerakan hewan ini.
Subordo ini dibedakan menjadi 2 family :
Family 1: Halteriidae. Peristom dan ikatan membranella di bagian
posterior di permukaan ventral. Contoh : Halteria , Meseres,
Metastrombidium, Strombidium, Tontonia.
Family 2 : Strombiidae. Zona membranela membentuk sebuah spiral di
ujung anterior. Contohnya : Cephalotrichium, Ciliospina, Lohmaniella,
Strombilidium, Parastrombidium, dan Sphaerotricium.
Subordo 3 : Tintinnina
Ciri dari anggota subordo ini adalah adanya sebuah konikal atau trumpet
bebrbentuk siliata yang berisi sebuah lorika dimana tertahan oleh adhesive aboral
tip pada badannya. Bagian peristomial menutupi lebih pada bagian lubang oral
65
dan lebih funnel berbentuk area leading sebuah sitostoma. Ada juga zona untuk
12-24 membranela yang berbentuk sebuah spiral sekitar peristom. Di bagian
adoral biasanya disebut tentakuloid, satu di antara sepasang membranella.
Masing-masing tentakuloid adalah sebuah bola yang membentuk struktur di
tangkai, dimana dasar dari sebuah konikal “accessory comb” bertahan pada area
peristomal. Belum diketahui fungsi dari tentakuloid, beberapa siliata terlihat
jarang ada, beberapa terbatas pada tubuh bagian anterior ketiga, kadang-kadang
juga terlihat di bagian posterior akhir.
Subordo 4 : Entodiniomorphina
Siliata ini terdapat pada rumen dari lembu, domba, maupun hewan
ruminansia lainnya, dan juga pada intestin pada herbivora. Bentuk siliata
mereduksi dan sebagian besar Ophryoscolecidae, yaitu Entodinium, terbatas pada
membranella pada zona adoral. Pada sebagian besar Ophryoscolecidae, tetapi ada
juga sebuah bagian dorsal membran, terbentang dari ikatan anterior pendek hingga
panjang dan sedikitnya mendekati garis akuator. Membran adoral muncul dari
sebuah alur yang terbentuk oleh sebuah lipatan ektoplasmik dan perluasan spiral
ke sitostom yang terletak di lempeng oral di lubang anterior. Sedikitnya di zona
adoral, dan di beberapa yang lain juga ada zona dorsal, yang dapat mungkin
terjadi lagi. Antara zona adoral dan dorsal ada sebuah sudut, membentuk
operculum. Makronukleus, mikronukleus, vakuola kontraktil, dan skeletal juga
66
terletak di zona, yang terpisah dari endoplasma oleh membran berlanjut pada
bagian anterior ke faring dan bagian posterior berlanjut ke rektum.
Subordo Entodiniomorphina dibedakan 2 famili:
Ophryoscolecidae dengan tidak lebih dari satu „dorsal‟ gabungan membran
di zona adoral, dan
Cycloposthiidae, yang memiliki lebih dari grup caudal di bagian posterior.
Subordo 5. Hypotrichina
Silia somatic digantikan oleh cirri yang hampir selalu terbatas pada
permukaan ventral. Permukaan dorsal sering terdapat garis yang disebut bulu
sensor (sensory bristle) yang kadang-kadang muncul pada permukaan ventral
berdekatan dengan cirri. Letak cirri pada berbagai spesies :
2. Frontal cirri : terletak diantara peristome dan pada sisi kanan tubuh.
3. Ventral cirri : memanjang pada posterior sampai frontal cirri
4. Marginal cirri : timbul dari sisi kanan dan kiri hingga batas permukaan
ventral
5. Caudal cirri : muncul dari batas posterior:
6. Anal cirri : timbul secara tranversal atau diagonal dari posterior hingga
ujung tubuh.
Daerah peristomial berukuran luas dan berbentuk kurang lebih segitiga
pada banyak spesies tapi ukurannya berkurang dan sempit pada spesies lainnya.
Pada zona mebranelles (semacam membran) sering memanjang dari cystome
bagian anterior lalu secara transversal menuju daerah antero-dorsal atau anteroventral pada permukaan tubuh.
67
Famili 1. Aspidiscidae
Ciri-ciri :
a. Memiliki silia yang rata
b. Memiliki pelllicle seperti armor (baju)
c. Pada bagian anterior terdapat membranelles yang menandakan adral primitive
zone
Family 2 Euplotidae
Ciri-ciri :
a. Ventral cirri mereduksi, dan kehilangan pada bagian batas kanan.
b. Anal cirri berkembang dengan pesat
c. Bagian peristome dan membranelles berkembang.
Family 3. Oxytrichidae
68
Ciri-ciri :
a. susunan dari cirri mengikuti pola umum.
b. Pada batas kanan dan kiri selalu memiliki cirri
c. Adoral-membranelles berkembang dengan baik.
Family 4. Paraeuplotidae
Ciri-ciri :
a. Pada daerah adoral-membranelles berkembang dengan baik
b. Cirri menjulur secara berkelompok sekitar 5 sampai 6 grup dari posterios
hingga ujung tubuh.
Subordo 6. Ctenostomina
Pada subordo ini ditandai dengan ciliata yang wedge-shaped (berbentuk
terjepit) yang biasanya ditemui pada permukaan ventral (oral). Batas (bentuk)
dorsal biasanya bulat mungkin juga semi bulat atau hampir mendekati bulat.
Tubuhnya diselimuti oleh frim pellicle yang terbentuk dari diferensiasi mejadi
piringan longitudinal. Sebagian besar spesies somatik siliatanya mengalami
reduksi.
69
Family 1. Epalcidae
Gambar Pelodinium , Lauterborn 1908.
Ciri-ciri:
a. Pada ujung posterior tubuh tidak terlapisi, tetapi dikelilingi oleh duri pada
piringan pelindung.
b. Pada daerah frontal band, dan empat primitive posterior selalu muncul pelapis
(armor).
c. Bagian ventral dan dorsal juga terlindungi oleh lapisan pelindung (armor).
Family 2. Mylestomidae
Ciri-ciri:
a. Pada kanan dan kiri dorsal, ciliatanya telah hilang atau mereduksi dalam jumlah
tertentu
b. Ciliata pada daerah posterior yang terssisa mungkin membentuk satu atau dua
“rudder cirri”.
Family 3. Discomorphidae
Ciri-ciri :
a. Daerah perlekatan pada dorsal yang merupakan akhir dari bagian anterior yang
berbentuk duri.
b. Cilia pada daerah somatic terbatas menjadi dua arah ventral , dua posterior pada
sebelah kiri dan berkembang baik pada frontal band.
2. Ordo Peritrichida
Protozoa dewasa biasanya melekatkan diri secara langsung dengan lainnya
menggunakan ujung bagian aboral tubuh atau dengan tangkai hasil sekresi,
maupun menempel pada permukaan benda padat. Beberapa tangkai bertipe koloni.
70
Hanya sedikit spesies yang berenang bebas dan rupanya tidak memiliki tahap
sesil.
Pada ordo ini, peristom atau epistom merupakan suatu lempeng polar yang
tampak dari bagian oral pada ujung tubuh dan diperkirakan berbentuk melingkar.
Selain itu, terdapat dua baris atau lebih silia yang biasanya mengelilingi peristom
searah jarum jam sebelum melewati sitostom ke dalam vestibula. Dua baris silia
adoral digambarkan pada Telotrochidium dan Cyclochacta, sedangkan tiga baris
pada Vorticella. Dari ketiganya, setiap baris silia terlihat bebas pada bagian distal
namun megalami penggabungan di bagian basal hingga masuk ke dalam
membran. Setiap baris silia berlanjut masuk ke dalam vestibula pada Cyclochacta
dan Vorticella, namun hanya baris silia yang lebih dalam pada Telotrochidium. Di
dalam vestiula, suatu membran dapat terbentuk oleh penggabungan silia (baris
lebih luar dari Votricella). Batas luar permukaan peristomial meliputi suatu daerah
sempit, hingga sebuah sebuah rak proyeksi seringkali membentuk suatu lingkaran
kontraktil yang dapat mengkerut untuk melingkupi peristom dan siliatur.
Vestibula menerima isi vakuola kontraktil, terkadang melalui suatu
“penampungan” sementara, dan juga bahan yang belum tercerna dari vakuola
makanan lama. Selama di dalam faring, partikel makanan yang masuk juga
melewati vestibula masuk ke dalam vakuola makanan yang sedang berkembang
sebagai sumbernya. Nolan dan Fin telah mencatat pemisahan yang jelas mengenai
saluran masuk dan keluar di dalam vestibula. Scopula merupakan suatu daerah
berbeda pada ujung aboral, seringkali ditunjukkan sebagai suatu invaginasi,
dinding yang terkadang memperlihatkan fibril atau bentukan seperti tongkat. Pada
banyak spesies sesil tidak bertangkai, scopula tampak mensekresikan beberapa
materi yang melekat pada substratum, sedangkan pada spesies yang bertangkai,
scopula mensekrresikan sedikit matriks dari tangkai. Tangkai non-kontraktil pada
Epistylidae seluruhnya terdiri atas bahan sekresi. Tangkai dari Vorticellidae
mengandung setidaknya sebuah myoneme spiral bebas atau “stalk-muscle” yang
71
tumbuh ke arah luar tubuh. Stalk-muscle pada koloni Vorticellidae dapat
terbentuk terus-menerus sepanjang koloni, kecuali mungkin pada bagian basal
tangkai utama atau pada myoneme tangkai individu yang independen. Pada
mulanya, keseluruhan koloni mungkin dapat kembali menuju pusat perlekatan,
pada Carchesium, kontraksi dari tangkai mempengaruhi masing-masing individu
secara terpisah.
Siklus hidup Peritrichida umumnya dimorfik dan terkadang polimorfik.
Mengenai reproduksi sepertinya harus dipertimbangkan pada pembelahan
72
daripada budding, meskipun salah satu keturunan seringkali lebih kecil daripada
lainnya. Metamorfosis menjadi dewasa disertai penempelan scopula pada
permukaan tertentu. Peritrich muda disebut Scyphidia, yang hanya memiliki
tangkai dalam jumlah sangat sedikit. Meski begitu, tahap Scyphidia pada
Vorticella terjadi pada waktu yang sangat singkat. Segera setelah penempelan,
dimulailah sekresi sebuah tangkai dan silia aboral menghilang dalam beberapa
menit. Pada pembelahan selama perkembangan tipe koloni, seperti Zoothamnium,
organisme baru (keturunan) akan membentuk tangkai baru yang akan menjadi
cabang dari tangkai utama.
Telotroch merupakan tahapan umum dalam siklus hidup dan tidak terbatas
hanya pada spesies bertangkai karena terdapat bentuk seperti Scyphidia.
Transformasi secara langsung bentukan tangkai menjadi sebuah telotroch kadangkadang terjadi pada Vorticella dan merupakan cara yang normal dalam
perkembangan aseksual pada beberapa genus seperti Zoothamnium. Setelah
terlepas dari koloni induk, tahap perpindahan dilakukan dengan berenang menjauh
kemudian menempatkan diri pada suatu objek untuk berkembang menjadi koloni
baru. Urceolariidae terkadang dinilai memiliki telotroch tetap yang sangat khusus.
Hal ini yang menarik dari telotroch Epistylis horizontalis adalah kemiripannya
dengan Urceolariidae pada ukuran serta perbandingan kemampuan pergerakan di
atas permukaan. Telotrochidium juga menunjukkan telotroch tetap walaupun
bukannya tidak mungkin bahwa pada genus ini terdapat telotroch yang memiliki
tahap sesil yang tidak jelas.
73
Siklus hidup dari komensalis, Ellobiophrya donacis lebih rumit daripada
sebagian besar Peritrichida lainnya. Organisme dewasa hodup di dalam sebuah
lamellibranch, Donax vittatus, yang menempel pada lembaran/filamen insang.
Pada tahap akhir pembelahan, organisme baru (keturunan) tetap menempel pada
organisme yang lebih besar (induk) menggunakan bagian isthmus sempit dari
sitoplasma. Sebuah scopula segera berkembang di dalam telotroch embrionik
mensekresikan suatu tangkai yang memanjang beberapa jarak untuk dapat
menempel pada organisme lainnya. Pemanjangan telotroch yang sedang
berkembang dan diferensiasi silia aboral terjadi selanjutnya. Telotroch kemudian
berenang bebas, biasanya menerobos ikatan tangkai dan scopula, namun
terkadang justru membawa tangkainya hingga saat akan dilepaskan nanti. Setelah
mencapai filamen insang, tahap perpindahan ini akan menempel dengan ujung
bagian aboral dan mengembangkan lengan protoplasmik yang akan menjadi
pengait terhadap inangnya. Homologi aksial antara Peritrichida dan ciliate lainnya
adalah adanya sedikit ketidakpastian, meskipun peristom biasanya terletak di
bagian anterior, Telotrochidium henenguyi dan Cyclochaeta domerguei, keduanya
berenang dengan ujung aboral di bagian depan.
Famili 1. Astylozoonidae. Peritrich dalam family ini aktif berenang berpindah
dengan peristom di bagian depan. Selain tangkai, satu atau dua thigmotactic
bristle tampak dikembangkan pada ujung aboral.
Famili 2. Epistylidae. Pada tahapan sesil, scopula menghasilkan sebuah tangkai
yang tidak mengandung myoneme (stalk-muscle). Beberapa spesies bersifat
soliter sedangkan lainnya berkoloni.
Famili 3. Lagenophryidae. Merupakan ciliata berlorica yang memiliki lempeng
peristomial pada ujung leher yang kokoh, biasanya berupa bagian tubuh yang
dipanjangkan melalui mulut dari lorica.
74
Famili 4. Ophrydiidae. Mulut pada ujung tubuh diperpanjang hingga ke dalam
leher kontraktil. Di beberapa spesies, ujung aboral meruncing pada satu titik
namun membulat lebar pada lainnya. Scopula dapat atau tidak dapat menghasilkan
sebuah tangkai pendek.
Famili 5. Scyphidiidae. Peritrich sesil berscopula yang berfungsi sebagai organ
penempel. Tubuhnya terkadang memipih lebar pada ujung aboral, namun
terkadang meruncing ke daerah basal stalk-like.
Famili 6. Urceolariidae. Famili ini meliputi ektoparasit dan endoparasit khusus
yang sumbu oral-aboralnya sering memendek. Ujung aboral berupa lempeng pipih
yang dilengkapi dengan cincin kutikular. Contoh organisme dalam famili ini
adalah Cyclochaeta domerguei.
Famili 7. Vaginicolidae. Merupakan peritrich berlorica yang berbeda dari
Lagenophryidae dalam keseluruhan mulut (oral) pada ujung tubuh dipanjangkan
melebihi mulut pada lorica.
Famili 8. Vorticellidae. Famili ini meliputi empat genus dengan bentuk sesil yang
khas dengan berkembangnya tangkai kotraktil.
75
3.Ordo Chonotrichida
Merupakan ektokomensal yang sebagian besar aktif berenang pada
Crustacea, kecuali Trichochona lecythoides yang dilaporkan berasal dari perairan
Eropa. Tubuhnya kurang lebih seperti bentuk vas dan menempel pada inang
dengan lempeng basal atau tangkai yang pendek. Peristom terletak di ujung atas
tubuh dan biasanya dikelilingi oleh bentukan corong tipis. Umumnya, permukaan
tubuh tidak bersilia. Vakuola kontraktil terbuka ke dalam faring (vestibula) seperti
pada Peritrichida (Hall, 1961).
Budding dan konjugasi merupakan cara reproduksi organisme seperti yang
telah
dilaporkan.
Kaitan
antara
anggota
Chonotrichida
meninggalkan
ketidakpastian dan pekerjaan lebih lanjut mengenai morfologi dan siklus hidup.
Walaupun begitu, bebrapa informasi yang telah dipercaya saat ini oleh beberapa
pekerja/ilmuwan dapat mengindikasikan bahwa kelompok ini memiliki hubungan
lebih dekat dengan Holotrichida daripada ciliata lainnya. Tiga family dalam ordo
ini yang telah ditemukan oleh Mohr antara lain: Chilodochonidae, Stylochonidae,
dan Spirochonidae (Hall, 1961).
2. Kelas Suctorea
Bentuk tubuh pada berbeda pada spesies, seperti spherical, conical, seperti
tongkat, silindris, vermiform, atau tidak bercabang. Ciri-ciri jelas dari kelompok
yang memilki tentakel dan tidak adanya cilia pada tahap dewasa. Terjadi
pengurangan tentakel pada Endosphaera, yang meliputi endoparasit pada ciliata
tertentu.
76
Gambar 3.50 Tipe Tentakel Pada Kelas Suctorea
Tentakel mungkin terletak pada permukaan atau mereka muncul pada
dalam bentuk ikatan atau bentuk cuping atau lengan yang panjang. Dua variasi
dapat diketahui, salah satu tipe adalah capitate (Gambar 3.50, c-f) distally
berakhir pada perluasan yang rata atau bulat. tipe lain adalah tipe lilin yang lebih
atau kurang dari titik (A,B). Pada beberapa spesies, tentakel mengandung tabung
di dalam (gambar C,E), perluasan kedalam endoplasma dari jarak yang dekat.
Tentakel melekat pada ciliata yang cocok yang melakukan kontak dengan mereka
dan tenaganya cukup untuk menangkap mangsa yan lebih besar dari captor
(gambar H,I). Kelumpuhan yang cepat dari organisme yang ditangkap telah
diketahui. Secara singkat setelah kontak, protoplasma mangsa mulai mengalir
kebawah tentakel ke dasar tabung, dimana vakuola makanan terbentuk. Pellicle
mangsa dipecah oleh hisapan lisis alami pada saat kontak dengan tentakel tidak
tentu. dicerna dengan cepat. Tokophyra lemnarum, sebagai contoh, pencernaan
Euplotes patella sekitar 15 menit.
Aliran material ke tentakel selama makan menggunakan hisapan, sumber
yang tersisa memunculkan masalah. Mungkin aktivitas signifikan dari vakuola
kontraktil meningkat sekitar lima fold pada Tokophrya infusionum untuk
organisme yang memulai makan.
Tangkai suctorian, diperlihatkan pada beberapa spesies, tidak selalu
kontraktil tidak seharusnya homogen pada struktur. Akhir tangkai mungkin
diperluas dengan cangkir kecil yang menjadi dasar dari sisa tubuh, atau pada
kasus lain, akhir distal dari tangkai ke dasar tubuh (gambar G).
77
Pada
metamorfosis larva ciliata, tangkai muncul dari analog organela ke scopula dari
Peritrichida (Gambar 3.51 L).
Beberapa Suctorea yang dilengkapi dengan sekret lorica yang terbuka pada
bagian distally, yang meninggalkan akhir apikal dari tubuh (Gambar 3.56 A) atau
mungkin dinding tebal yang dekat pada tubuh Squalophrya macrostyla (Gambar
3.53, H, I).
Hubungan antara Suctorea ke ciliata ditunjukkan pada siklus hidup pada
sebagian spesies. Secara tipical reproduksi melibatkan pertunasan secara internal
(Gambar 3.51, N-P) atau eksternal. Walaupun terlihat tidak biasa, kedua
pertunasan internal dan eksternal mungkin terjadi pada spesies tunggal, yang
diperlihatkan pada Anarma multiruga. Tunas berkembang menjadi larva ciliata
(Gambar 3.51, A-K), dalam waktu singkat akan berenang, dan mengalami
metamorfosis alami. Setelah tahap larva, Tokophrya lemnarum menjadi terikat
(Gambar 3.51, L-M), sebuah tangkai keluar dalam beberapa menit, lengan
tentakel tumbuh normalsekitar 15 menit setelah mereka melakukan deteksi
pertama, dan bentuk dewasa berkembang secara penuh sekitar satu jam. Kejadian
metamorfosis yang cepat dimiliki pada Tokophrya infusionum.
Gambar 3.51 Tipikal reproduksi kelas Suctorea
Penghilangan cilia selama metamorfosis tidak termasuk granula basal
mereka, yang dilakukan pada tahap dewasa Podophrya fixa. Pada reproduksi,
tunas mendapatkan beberapagranula basal parental, kemudian berlipat dan
memberikan kenaikan pada larva cilia. Jika pada kasus ini mungkin
dipertimbangkan tentang representatif, adanya kelestarian genetic dari granula
basal sepanjang siklus hidup.
78
Gambar 3.52
Pada siklus hidup Podophrya fixa, pertengahan tahap intervensi antara
larva dan dewasa, parasit pada Nassula ornata (Gambar 3.65, F). Hasil dari
metamorfosis Sphaerophrya pada tahap mengapung sampai membuat kontak
dengan inang ciliata. endhosphaera juga termasuk tipe yang tidak biasa dengan
endoparasitik dewasa, dilekatkan pada citoplasma dari inang ciliata. Pada
reproduksinya, hasil dari parasit larva ciliata yaitu hasil bebas dari tahap migrasi
dan menempel pada inang baru (Gambar 3.53, A-D).
Gambar 3.53
Tambahan pada kejadian umum larva ciliata, transformasi dari dewasa ke
tahap migrasi juga mungkin terjadi, pada Podophrya parsitica (Gambar 3.56, D).
Tahap migrasi ini, secara bertahap ditambah oleh perkembangan tangkai, cilia,
dan perkembangan tentakel diatas permukaan menjadi tahap Paracineta (Gambar
3.56, G). Tahap ini tidak makan dan nampak pada tahap sementara didahului
ekistasis (Gambar 3.56, H).
79
Gambar 7.54
Konjugasi, dapat dibandingkan pada tipe ciliata, yang dijelaskan pada
beberapa jenis, termasuk Acineta, Dendrocometes dan Tokophrya. Perbedaan
konjugasi pada Tokophrya lemnarum, pada dua konjugan terpisah pada ciliata.
fusi lengkap dan produksi dari sinkronisasi tunggal yang ditunjukkan pada
Lernaeophrya capitata.
Ekistasis diketahui pada jumlah spesies. Pada Tokophrya lemnarum ,
ekistasis melibatkan deposisi dari sekresi transparan pada basal pertama, dan yang
terakhir adalah akhir apical tubuh. Spesimen awal ekistasis yang menyerupai
loricate suctorea. Precistic dengan drawal tentakel yang tidak dapat djelaskan.
Sebagai gantinya, struktur yang tersisa pada permukaan apical dan ditelah oleh
sekresi material ke bentuk membran kista.
Taxonomy
Suctorea, berkaitan dekat dengan gymnostoma Holotrichida kemudian ke
ciliata yang lain, tampak memiliki dasar defersivikasi pada evolusinya. Sejak
tidak ada dasar logical dalam perbedaan tingkatan, suatu kelompok yang dibagi
kedalam nomer famili. Bahkan pengaturan sederhana menjadi lebih berguna jika
beberapa famili mendefinisikan secara jelas untuk memperhatikan jenis yang
memperlihatkan kombinasi intermediet pada karakteristik. Mungkin dilakukan
studi yang intensif pada siklus hidup, dengan komparasi yang rinci dari migrasi
larva, yang mungkin menghasilkan informasi yang berguna. Sebagai contoh, hal
yang menarik pada larva Podophrya soliformis dan Parapodophrya denticula
famili Podophryidae) yang dapat dijelaskan dengan polar circlet dari cilia yang
80
serupa yang dilihat dengan sabuk equatorial dari cilia yang sama pada spesies
Tokophrya (famili Acinetidae ); larva Podophrya globulifera, P.fixa dan P.
parasitica, dengan baris cilia yang pararel pada panjang poros.
1. Famili acinetidae
famili ini dikarekteristikan oleh tunas endogen dan kepemilikan tentakel ,
biasanya disusun pada kelompok. Sebuah lorica sering hadir dan tangkai mungkin
hadir atau tidak.
Famili Dendrocometidae. bentukan tanpa tangkai pada pertunasan endogen.
Kepala tentakel didistribusikan ke permukaan atau tempat yang sedikit besar dari
tubuh.
2. Famili Dendrosomidae
badan tanpa tangkai yang memiliki bentuk tidak teratur dan bercabang. dan
permukaan basal yang biasanya berada di substratum. Tentakel yang disusun
pada cluster. Tipe reproduksi melibatkan tunas endogen, yang tunggal maupun
berlipat.
3. Famili Discophryidae
Pertunasan endogen dan karakteristik kepala tentakel, walaupun beberapa spesies
demgan tetakel yang tepat
yang berperan dalam famili. Sebuah lorica yang
berkurang dan sebuah tangkai yang mungkin atau tidak ada.
4. Famili Ephilotidae
Tangkai pada suctorea laut dengan kepala atau titik tentakel. Pertunasan adalah
tipe exogen dan mungkin multiplet. Sebuah lorica mungkin atau tidak mungkin
ada.
Gambar 3.55
81
Gambar 3.56
5. Famili Ophryodendridae
Di laut bentukan tentakel dikonsetrasikan pada satu atau lebih gerakan proboscis
seperti pemanjangan. Pada penambahan yang tidak wajar pada tipe larva, jadi
disebut tunas verniform, yang tidak memilki tentakel atau probocis, yang mungkin
dihasilkan oleh pertunasan exogen.
6. Famili Podophryiade
reproduksi melibatkan pertunasan eksternal atau beberapa kasus “pembelahan”
yang memiliki dua organisme sesaudara hampir sama jumlahnya. Sebuah tangkai
dan sebuah lorica mungkin hadir atau tidak ada.
RINGKASAN
Karakterisasi morfologi Filum Ciliophora yang paling utama, yakni memiliki silia
pada beberapa tahap dalam siklus hidupnya. Silia membungkus seluruh tubuh
organisme hingga batas area peristomial. Silia terdapat di permukaan sel yang
memiliki bentuk menyerupai rambut. Ciliophora memiliki dua jenis nukles, yakni
makronukleus dan mikronukleus. Filum ini dibedakan dua kelas, yaitu Ciliatea
dan Suctorea. Pada Ciliatea, silia ada pada sebagian besar tahap dari siklus hidup,
sedangkan dalam Kelas Suctorea memiliki tahap non-ciliated pada tahap dewasa
serta memiliki tentakel khusus yang berfungsi mencari makan. Ciliophora
merupakan protozoa kosmopolitan yang ditemukan pada bermacam-macam
habitat, khususnya di perairan.
Ordo
Holotrichida
memiliki
6
subordo,
yaitu
Gymnostomina,
Trichostomina, Hymenostmina, Thigmotrichina, Apostomina, Astomina. Subordo
82
Gymnostomina memiliki sitostom terbuka di permukaan sentral bagian anterior.
Pada Subordo Trichostomina sitostom terletak di sentral permukaan. Subordo
Hymenostima memiliki cirri pada ciliature peristomial telah termodifikasi
menjadi beberapa membran. Pada Thigmotrichina sitostom bergeser ke posisi
pada atau dekat ujung posterior tubuh. Subordo Apostomin sitostom perut sangat
berkurang. Pada Subordo Astomina tidak memiliki sitostom.
Masing-masing ordo dalam Filum Ciliophora memiliki ciri khas yang
berbeda-beda tergantung pada ordonya. Sebagian besar memiliki silia di
permukaan tubuhnya, meski begitu pada kelas Suctorea terdapat tahapan dewasa
yang tidak memiliki silia serta memiliki tentakel khusus untuk menangkap
mangsa.
DAFTAR PUSTAKA
Ehrenberg. 1830. Loxodes, (Online), (http://protist.i.hosei.ac.jp/pdb/images/Ciliophora/Loxodes/index.html) diakses 29 Oktober 2011
Hall. 1961. Protozoology. Japan: Prentice-Hall, Inc.
Lynn, D.H. and Small, E.B. 1991. Handbook of Protoctista (Philum Ciliophora).
Boston: Jones and Bartlett Publishers.
Roger, A.O. 1988. Comparative Protozoology, Ecology, Physiology, and Life
History. New York: Sringer- Verlag New York Inc.
83