POKOK BAHASAN 8. BUNGA Struktur bunga serta
advertisement

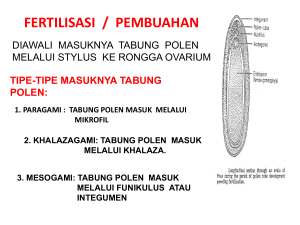
POKOK BAHASAN 8. BUNGA Struktur bunga serta bagian- bagian bunga Bunga merupakan alat reproduksi seksual. Bunga dikatakan lengkap apabila mempunyai daun kelopak, daun mahkota, benang sari, putik, dan daun buah. Bunga terdiri atas bagian fertil, yaitu benang sari dan daun buah, serta bagian yang steril yaitu daun kelopak dan daun mahkota. 8.1 DAUN MAHKOTA DAN DAUN KELOPAK Secara anatomi daun mahkota dan daun kelopak mempunyai struktur yang sama, terdiri atas sel-sel parenkimatis. Parenkim dasar terletak di antara epidermis atas dan epidermis bawah. Jaringan ini juga disebut mesofil. Sistem pembuluh terdapat pada jaringan dasar. Pada jaringan dasar mungkin terdapat sel-sel yang mengandung kristal, idioblas atau saluran getah/ sel getah. Sel-sel tersebut berhubungan dengan unsur pembuluh. Daun kelopak suku Geraniacea mempunyai hipoderinis yang berdinding tebal, masing-masmg dengan kristal drusen. Daun kelopak sel-selnya mengandung kloroplas. Epidermis daun kelopak dilapisi kutin pada bagian luarnya, terdapat stomata dan trikomata, seperti pada daun. Struktur sistem pembuluh seperti pada daun hanya kurang jelas strukturnya. Daun mahkota mempunyai satu atau banyak pembuluh yang kecil-kecil. Epidermis bentuknya khusus, merupakan tonjolan yang disebut papila, dilapisi oleh kutikula. Adanya warna yang bermacam-macam pada daun mahkota disebabkan oleh adanya kromoplas atau pigmen tambahan yang terdapat pada cairan sel. Zat tepung sering dibentuk pada daun mahkota yang masih muda. Minyak volatil yang karakteristik pada bunga umumnya terdapat pada sel-sel epidermis. Gambar 8.1. Diagram struktur anatomi petala beberapa jenis tumbuhan A. Amelanchia laevis; B. Lysimachia nummularia; C. Pinguicula vulgaris t. trikoma kelenjar; u. ruang sekretoris sek (dikutip dari Eames & McDaniels, McDa 1953) 8.2 BENANG SARI Benang sari terdiri atas kepala sari sa dan tangkai sari. Tangkai sari tersusun oleh jaringan dasar, yaitu sel-sel sel parenkimatis yang mempunyai vakuola, tanpa ruang antar sel. Sel-sel ini sering mengandung pigmen. Epidermis dengan kutikula, trikoma atau mungkin stomata. Kepala sari mempunyai struktur struktur yang sangat kompleks, terdi terdiri atas dinding yang berlapis-lapis, lapis, dan di bagian terdalam terdalam terdapat loculus/ruang sa sari (mikrosporangium) yang berisi isi butir-butir butir serbuk sari. Jumlah lapisan dinding kepala sari untuk setiap jems tumbuhan bervariasi. Struktur kepala sari (antera) Pada umumnya suatu antera terdiri atas 4 mikrosporangia krosporangia (4 lokuli). Pada waktu masak 2 sporangia dan masing-masing mas sisi akan menyatukan diri menjadi teka, sehingga ada 2 teka. Suatu keadaan yang berbeda, bahwa pada pada antera terdapat jaringan steril yang disebut septa, memisahkan me deretan lobus, misalnya salnya pada beberapa anggota suku Inimosace mosacea. Jems lam seperti Viscum, masing-masing masing polen dikelilingi oleh jaringan pelindung, dung, dan letaknya berderet-deret, berderet deret, secara horis horisontal dan vertikal sehingga masing-masing masing antera mempunyai 50 lokuli. Gambar 8.2. Struktur kepala sari pada bunga Lilium sp. A. Penampang lintang kepala sari muda mud Kepala sari terdiri atas 4 lobi (lokuli), tapetum menyelubungi jaringan sporogen. B. Penampang lintang kepala sari sa dewasa (masak) Antera masak dengan serbuk sari yang yang banyak. Kedua lobi pada mas masingmasing sisi mengadakan persatuan, disebut teka. Lamina fibrosa (endotesium) tampak lebih tebal, epidermis menipis. en. Endotesium; ep. Epidermis; Epidermis js. Jaringan sporogen (sel induk mikrospora mikrospora); k. konektivum; 1. lapisan tengah; ss. Serbuk sari (pollen); st. stoinium; sto um; ts. Sisa tapetum; t. tapetum. (dikutip dan Foster ter & Gifford, 1974; Maheswari, 1950) Perkembangan kepala sari (antera) Suatu antera yang muda terdiri atas suatu masa sel yang homogen yang dikelilingi oleh lapisan epidermis. epidermis. Selama perkembangan antera menghasilkan 4 lobi dan setiap lobus beberapa sel hipodermal menjadi lebih menarik perhatian diband dibanding yang lain karena ukurannya yang besar, bentuk selnya memanjang ke arah radial dan intinya jelas. Sel-sel ini adalah sel arkesponum. Sel-sel sel arkesporium membelah dengan dinding perikimal ikimal (sejajar permukaan) permukaa menghasilkan sel-sel sel parietal primer di sebelah luar dan sel-sel sel sporogen primer di sebelah dalam. Sel-sel Sel sel parietal primer membelah lagi secara periklinal menghasilkan lapisan parietal sekunder. Lapisan parietal sekunder nder inilah yang nantinya nant akan menghasilkan dinding antera. Sel sporogen primer membelah-belah membelah lagi secara mitosis, tosis, dan sel sel-sel hasil pembelahan mitosis menjadi sel induk mikrospora.. Sel sporogen primer dapat langsung berfungsi sebagai sel induk mikrospora tanpa mitosis.. Setelah itu sel induk mikrospora membelah secara meiosis menghasilkan tetrad mikrospora.. Selanjutnya sel-sel dalam tetrad memisahkan sahkan diri di menjadi sel mikrospora yang soliter. (Skema perkembangan antera lihat gambar!) Gambar 8.3. Skema perkembangan kepala sari. Lapisan dinding kepala sari dan mikrospora berasal dari jaringan arkesporium. Gambar 8.4. Struktur dan perkembangan kepala sari pada tumbuhan Angiospermae A1, B1 : Jaringan meristematis dikelilingi epidermis Sel-selnya mempunyai intl yang jelas C1 Sel-sel hipodermal terdiferensiasi menjadi sel-sel arkesporium D1 Lapisan parietal primer dan sel spongen primer telah terbentuk. E1 : Lapisan parietal primer mulal membelah E:epidermis, m:lapisan tengah, sp:sel sporogen primer, t:sel induk Tapetum Gambar 8.5. Struktur antera pada beberapa anggota Mimosacea A.pada Parkia ada 2 deret pollinia. B dan C. pollinia (kumpulan tetrad mikrospora) D. penampang bujur antera Pichrostachys menunjukkan kumpulan pollinia yang berderet deret. gl:glandula (disadur dan Maheswari. 1950’) Menurut Bhojwani dan Bhatnagar (1978, 1999) kepala sari mempunyai lapisan dinding sebagai berikut. Epidermis (eksotesium) Merupakan lapisan terluar, terdiri dari satu lapis sel. Epidermis menjadi memipih dan membentuk tonjolan (papila) pada kepala sari yang masak, dan berftingsi sebagai pelindung epidermis. Disebut eksotesium apabila sel-selnya mengalami penebalan berserabut. Endotesium Endotesium merupakan lapisan yang terletak di sebelah dalam epidermis. Pada kepala sari yang masak endotesium mengadakan penebalan ke arah radial, tangensial sebelah dalam atau antiklinal. Penebalan sel tersebut tidak teratur dan menunjukkan struktur berserabut. Adanya struktur berserabut menyebabkan endotesium mempunyai fungsi untuk membantu membukanya antera. Dengan adanya struktur yang berserabut pada dindingnya maka endotesium sering disebut lamina fibrosa. Endotesium biasanya hanya satu lapis sel, tetapi beberapa kepustakaan menyebutkan ada yang terdiri atas beberapa lapis sel. Pada tumbuhan air biasanya tidak dijumpai adanya penebalan berserabut pada endotesium. Pada tumbuhan kleistogam (bunga tidak pemah membuka) serta beberapa jenis termasuk Hydrochanitaceae, endotesium gagal mengadakan perkembangan, sehingga mikrospora (butir serbuk sari) keluar melalui lubang di bagian apikal kepala sari. Lapisan tengah Lapisan tengah merupakan lapisan yang terletak disebelah dalam endotesium, terdiri dan 2-3 lapis sel atau lebth, tergantung jenis tumbuhannya. Dengan berkembangnya antera sel-selnya menjadi tertekan dan memipih, karena terdesak oleh endotesium, sehingga sering pula disebut lapisan tertekan. Keadaan ini terjadi pada waktu sel induk spora (sporosit) mengalami pembelahan meiosis. Pada tumbuhan tertentu tidak dijumpai adanya lapisan tertekan. Tapetum Tapetum merupakan dinding terdalam dari antera dan berkembang mencapai maksimum pada saat terbentuknya serbuk sari tetrad. Lapisan tapetum berfungsi memberikan seluruh isi selnya selama perkembangan mikrospora. Tapetum umumnya merupakan derivat lapisan parietal primer. Namun pada suatu spesies, misalnya pada Alectra thomsoni sel-sel tapetum mempunyai 2 tipe berdasarkan atas sel penyusunnya, yaitu: 1. Sel tapetum berukuran besar, merupakan derivat dan sel-sel konektivum; 2. Sel tapetum !ebih kecil dibanding tipe pertama, merupakan derivat dan lapisan parietal primer. Menurut Maheswari Devi (1963) tapetum pada Calotropis gigantea terdiri dari beberapa lapis sel. Menurut Bhojwarn dan Bhatnagar (1999) ada 2 tipe tapetum, yaitu: a. Tapetum ameboid (plasmodial) Pada tipe ini tapetum mengeluarkan seluruh masa protoplasnya ke dalam lokulus (ruang sari) dan dinding selnya mengalami lisis. Kemudian protoplas tapetum ini menggabungkan diri dengan protoplas yang ada di da!am lokulus, se!anjutnya protoplas tersebut bergerak menyelubungi sel induk spora. Tapetum tipe ini biasanya dijumpai pada tumbuhan Monocotyledoneae dan Dycotyledoneae tingkat rendah. b. Tapetum sekresi (glandular) Tapetum menge!uarkan isi selnya secara berkala, sedikit demi sedikit. Dinding selnya tidak mengalami lisis, dan sisa selnya masih dapat dilihat selama perkembangan mikrospora. Tipe ini dijumpai pada tumbuhan Angiospermae yang telah maju tingkatannya. 6.3 MIKROSPOROGENESIS Setiap jaringan sporogen kadang-kadang langsung berfungsi sebagai sel induk mikrospora, atau mungkin mengalami beberapa kali pembelahan mitosis, sehingga jumlah selnya bertambah banyak sebelum menga!aini meiosis. Se! induk mikrospora (disebut pula sporosit) mengalami pembelahan meiosis, menghasilkan mikrospora yang bersifat haploid. Sitokinesis Pembentukan dinding setelah pembelahan meiosis sel induk mikrospora dapat terjadi secara susesifatau secara simultan. Secara susesif Setelah pembelahan meiosis, terbentuk dinding yang memisahkan dua inti, sehingga stadium 2 sel (diad). Pembentukan dinding secara sentrifugal (dari bagian tengah ke tepi). Pada stadium meiosis II, dinding pemisah dibentuk dengan cara yang sama, sehingga terbentuk serbuk sari tetrad yang bertipe isobilateral. Misalnya pada Zea mays. Secara simultan Pada pembelahan meiosis I tidak diikuti pembentukan dinding, sehingga terdapat stadium 2 inti (binuldeat). Jadi disini tidak terdapat stadium 2 sel. Selanjutnya 2 inti tersebut mengadakan pembelahan, terbentuk serbuk sari tetrad yang bertipe tetrahidris.Contoh: Dryinis winteri Gambar 8.6 Pembentukan dinding pollen secara susesif menghasilkan tipe tetrad isobilateral. A. sel induk mikrospora B. pembelahan meiosis I C. awal pembelahan meiosis II D. fase anafase pembelahan meiosis II E. akhir pembelahan meiosis II, dthasilkan 4 sel (tetraci) mikrospora. Gambar 8.7 pembentukan dinding pollen setelah pembelahan sel induk mikrospora tipe simultan A - D. pembelahan meiosis I tanpa dinding sekat. E - I. Pembelahan meiosis II. E, F:diantara inti terdapat vakuola kecil, terjadi ikatan longgar (lihat daerah yang berwarna putth), G-I. Mulai terbentuk dinding pemisah dari bagian tepi ke tengah. Tetrad Mikrospora Pada umumnya susunan mukrospora pada tetrad adalah tetrahidris atau isobilateral. Tetapi pada jenis yang lain susunan tetrad mikrospora adalah: dekusata, linier, bentuk huruf T. Gambar 8.8. Tipe tetrad mikrospora pada Angioispermae 1. tetrahedral; 2. isobilateral; 3. dekusata; 4. bentuk T ; 5. linier. Perkembangan Gametofit jantan Mikrospora merupakan awal dari generasi gametofit jantan. Mikrospora dewasa yang telah lepas dari tetrad, dikenal sebagai butir pollen (serbuk sari). Serbuk sari I pollen pada uintimnya mempunyai 2 lapisan dinding yaitu eksin merupakan lapisan terluar dari inti lapisan dalam. Eksin tersusun dari sporopolenrn, sedang inti tersusun dan polisakarida. Serbuk sari yang baru terbentuk mempunyai sitoplasma yang padat, dengan inti di bagian tengahnya. Setelah antera masak pollen keluar melalui lubang yang disebut stomium. Epidermis yang letaknya berdekatan dengan stomium dinding mengalami penebalan membentuk struktur yang khusus. Perkembangan pollen (Inikrogametogenesis) Pollen yang baru dibentuk umumnya mempunyai sitoplasma yang padat. Selnya secara cepat bertambah volumenya, diikuti oleh vakuolisasi dan perpindahan inti dari bagian tengah menuju ke bagian yang berdekatan dengan dinding sel. Pada tanaman tropis, biasanya inti segera membelah tetapi pada tanaman yang hidup di daerah dingin terdapat fase istirahat beberapa han sampai beberapa ininggu. Pada Tradescantia reflexa fase istirahat 4 hari atau kurang dari 4 hari, sedang pada Himantoglossum hircinum 2 sampai 3 ininggu. Pembentukan sel vegetatif dan sel generatif Pada awal gametogenesis inti serbuk sari membelah menjadi dua sel, yaitu sel vegetatif dan sel generatif. Kedua sel tersebut ukurannya tidak sama. Sel Vegetatif lebih besar dibanding sel generatif Selanjutnya Sel generatif membelah secara mitosis menghasilkan 2 sel sperma. Gambar 8.9. Perkembangan gametofit jantan A. Serbuk sari yang barn terbentuk dengan 1 inti. B. Serbuk sari membesar, inti pmdah ke bagian tepi, dan di bagian tengah terbentuk vakuola. C. Inti serbuk sari mengadakan pembelahan. D. Stadium 2 inti pada serbuk sari. Inti sel vegetatif lebih besar ukurannya dan terletak di bagian tengah. Sel-sel generatif letaknya dekat dengan dinding sel. E. Inti sel generatif mulai kehilangan kontak dengan dinding sel, dan bentuknya berubah menjadi bulat. F. Inti sel generatif terdapat bebas pada sitoplasma. G-H. Inti sel generatif mulai mengadakan pembelahan dan dan hasil pembelahan terbentuk 2 sel sperma. I-J. Inti sel generatif membelah di dalam buluh serbuk sari. Dinding pollen Dinding pollen berlapis-lapis. Dinding terluar disebut eksin dan dinding dalam disebut inti. Eksin terdiri atas ekteksin dan endeksin. Ekteksin tersusun oleh: 1. tektum di bagian luar; 2. bagian dalam adalah lapisan kaki (foot layer) berbatasan dengan endeksin; 3. bakulum lapisan yang terdapat antara tektum dan lapisan kaki. Eksin tersusun atas sporopolenin, merupakan derivat dan karotenoid yang mengalami polimerisasi oksidatif. Sporopoleurn sangat resisten terhadap faktor fisik dan dekomposisi biologik. Lapisan intin terdiri atas pekto-sellulose. Struktur selulose terdiri atas inikrofiblir yang tersusun paralel terhadap permukaan dinding. Gambar 8. 1 0. Struktur sel pollen pada Angiospermae A. sel pollen dilindungi oleh dinding yang tebal, dengan 2 inti yang jelas, yaitu inti vegetatif(besar) dan inti generatif(kecil) B. Perbesaran dan DP. b: bakulum; ek: eksin; in : intin; en : endeksin; t tektum; k: lapisan kaki. Perkembangan abnormal dan pollen Perkembangan abnormal dari polen dijumpai pada tubuh monokotil maupun dikotil. 8.4 PISTILUM Megasporangium dan Megasporogenesis Tumbuhan Angiospermae pada umumnya mempunyai megasporofil (daun buah) yang berkembang ke dalam suatu pistilum. Pistilum (putik) biasanya mengalami diferensiasi menjadi 3 bagian yaitu: 1. bagian basal yang menggelembung disebut ovarium (bakal buah). 2. bagian yang memanjang disebut stilus (tangkai putik) 3. bagian ujung stilus disebut stigma (kepala putik) Di dalam ovarium terdapat dua atau lebth dan dua ovulum (bakal biji). Ovulum berkembang (berasal) dan plasenta. Suatu ovulum terdiri atas: 1. megasporangium (kandung lembaga embiyo sac) suatu badan sentral, 2. merupakan hasil perkembangan lebih lanjut dan megaspora yang berfiingsi. 3. nuselus, yakni jaringan yang menyelubungi badan sentral. Nuselus diselubungi oleh sath atau dim integumen. 4. integumen, suatujaringan yang menyelubungi nuselus. 5. funikulus, tangkai yang mendukung bakal biji, dimana bakal biji itu melekat pada plasenta. Ukuran nuselus, jumlah integumen dan bentuk ovulum sangat pentmg untuk membedakan ciri khas suatu ovulum pada kelompok tumbuhan berbunga. Ovulum digolongkan ke dalam 5 tipe, tergantung aksis ovulum tersebut, apakah tegak atau melengkung terhadap Mikropil dan funikulus. Tipe ovulum tersebut adalah: 1. orthotropus : Mikropil menghadap ke atas terletak segaris dengan hilus. 2. Anatropus : Mikropil dan hilus letalmya sangat berdekatan. 3. Kampilotropus : ovulum berbentuk kurva. 4. Heinianatropus : apabila nuselus dan intigumen terletak kurang lebih di sudut funikulus. 5. amfitropus: ovulum berbentuk seperti sepatu kuda. Integumen Suatu ovulum kebanyakan mempunyal satu atau dua integumen. Ovulum dengan satu intigumen disebut unitegmik, dan yang mempunyai dua intigumen tersebut bitegmik. Pada tumbuhan Sympetalae umumnya menunjukkan keadaan unitegmik, sedang pada Polypetalae dan monokotil adalah bitegmik. Pada beberapa anggota Olacaceae menurut Davis (1966). ovulum tidak berintegumen dan disebut ateginik. Ovulum pada umumnya berasal dari jaringan plasenta di dalam ovarium, sedang integumen berasal dari bagian basal primordium ovulum. Keadaan unitegmik mungkin disebabkan karena hilangnya salah satu intigumen, seperti pada Cyilnus perkembangannya sehingga hanya mempunyai satu intigumen. Pada beberapa dijumpai adanya integumen ketiga atau arilus, Pada Ulmus dilaporkan, bahwa integumen ketiga berasal dari pembelahan integumen luar, tetapi struktur tersebut dapat pula berasal dari pangkal ovulum. Pada anggota Euphorbiaceae dikenal adanya karunkula yang berasal dari poliferasi sel-sel integumen di daerah mikrofil. Kadang-kadang poliferasi ini sangat kuat dan karunkula ini masih dapat dilihat sampai biji masak. Misalnya pada biji Ricinus Communis Mikropil Mikropil dapat dibentuk oleh integumen luar dan atau integumen dalam. Mikropil yang dibentuk oleh integumen dalam seperti pada Centrospermales dan Plumbagmales, oleh integumen luar dan dalam, seperti pada suku Pontederiaceae. Jarang sekali Mikropil dibentuk oleh integumen luar misalnya pada suku Podostemaceae. Rhamnaceae, dan Euphorbiaceae. Lubang Mikropil yang dibentuk oleh integumen luar disebut eksostoma, sedang yang dibentuk oleh intigumen dalam disebut endostoma. Tapetum integumen (endotelium) Pada beberapa tumbuhan nuselus segera mengalami disorganisasi dan kantong embrio langsung mengadakan kontak dengan lapisan integumen yang semula berbatasan dengan nuselus. Lapisan yang semula berbatasan dengan nuselus itu terdiferensiasi menjadi lapisan yang khusus, baik bentuk maupun kandungan selnya. Sel-selnya memanjang ke arah radial, kadang-kadang menjadi binuldeat (mengandung dua inti). Sel-sel ini mempunyai persamaan dengan sel-sel tapetum pada antera, oleh karena itu disebut tapetum integumen (endotelium). Endotelium berfungsi nutritif, membantu transport bahan makanan dan integumen menuju ke kantong embrio. Pada waktu embrio dewasa permukaan dalam dan lapisan endotelium mengalami kutinisasi dan lapisan tersebut berubah menjadi lapisan pelindung. Endotelium merupakan lapisan tunggal, dijumpai pada beberapa taksa yang mempunyai tipe intigumen unitegmik, seperti Compositae, Lentibulariaceae dan Orobanchaceae. Pada Compositae endotelium lebih dari satu lapis sel yaitu 2-10 lapisan seperti pada bunga matahari. Hipostase dan Epistase Hipostase adalah sekelompok sel yang terdapat di bawah kantong embrio di bagian khalaza, berhadapan dengan jaringan pengangkut yang ada di funikulus. Merupakan derivat sel-sel nuselus di bawah kantong embrio. Hipostase mempunyai dinding yang tebal dan dingin, sedikit sitoplasma. Jaringan inti terdapat pada beberapa suku, antara lain Crossosomataceae clan Umbelliferae. Kadang-kadang hipostase terbentuk setelah pembuahan. Epistase merupakan jaringan yang letaknya di daerah mikropil, dan dibentuk oleh sel-sel epidermis nuselus. Strukturnya seperti kaliptra pada akar, oleh karena itu sering disebut tudung nuselus (operkulum), misalnya pada Castalia dan Costus. Obturator Obturator adalah jaringan yang merupakan poliferasi sel-sel funikulus atau plasenta. Yang berasal dari funikulus misalnya pada famili Acanthaceae, Anacardiaceae, Labiatae dan Magnoliaceae. Jaringan ini berfungsi untuk membantu pembuahan yaitu memandu buluh pollen menuju mikropil. Sel-selnya mengalami degenerasi setelah terjadinya pembuahan. Pada Tetragonia tetragonioides obturator mempunyai struktur seperti trikomata (rambut-rambut) berasal dan epidermis kedua sisi funikulus yang letaknya berhadapan dengan mikropil. Obturator yang berasal dari sel-sel plasenta misalnya pada suku Euphorbiaceae dan Cuscutaceae. Gambar 8. 14. Struktur tambahan pada Ovulum I. Obturator pada Tetragonia tetragonloider Perhatikan perkembangan obturator pada tangkai funikulus yang sebelah luar lebih baik di banding dengan yang berdekatan dengan Mikropil (A,B). II. Endotelium pada Asteraceae: D. Volutacella ramose ; E. Glossocardia bosvallia. III. Integumen ketiga pada Trianthema monogyna. Nuselus Nuselus merupakan dinding megasporangium. Setiap ovulum hanya mempunyai satu nuselus. Yang mempunyai dua nuselus antara lain adalah Aegle marmelos dan Hydrocleis nymphoides. Pada awal terbentuknya calon ovulum, nuselus terbentuk lebih dulu, terdiri atas sel-sel yang homogen diselubungi oleh epidermis. Dibawah lapisan epidermis nuselus terdapat sekelompok sel-sel arkesporium. Pada Sympetalae sel-sel arkesponum berfungsi langsung sebagai sel induk megaspora (sel sporogen), sehingga sel sporogen adalah sel hipodermal (hipo = bawah; dermal = epidermis). Berdasarkan asal sel-sel sporogen (sel induk megaspora) maka nuselus dibedakan menjadi dua tipe yaitu: 1. tenumuselat Sel sporogen (sel induk megaspora) adalah sel hipodermal, sehingga sel sporogen berbatasan langsung dengan epidermis nuselus. 2. krasmuselat Antara sel-sel sporogen dengan epidermis nuselus dipisahkan oleh lapisan sel parietal primer. Gambar 8.15. Perkembangan ovulum tipe tenuinuselat dan tipe krassinuselat. I. Megasporogenesis pada ovulum Elytraria acaulis tipe tenumuselat. II. Megasporogenesis pada ovulum Myriophyllum intermedium tipe krassinuselat. Sel induk megaspora membelah meiosis menghasilkan tetrad linier (Gambar I dan II: C-D). ini. sel induk megaspora (sel sporogen) ; sp. sel parietal, en:epidermis nuselus. Megasporogenesis Pada ontogeni ovulum, nuselus terbentuk lebih dulu, merupakan masa sel yang diselubungi oleh epidermis, berasal dari proliferasi sel-sel plasenta. Suatu sel hipodermal pada nuselus mempunyai ukuran yang besar, sitoplasma padat dan ini besar berfungsi sebagai sel arkesporium. Sel ini membelah secara perildinal atau langsung berfungsi sebagai sel induk megaspora. Kalau membelah secara periklinal sel arkesporial tersebut ke arah dalam menghasilkan sel sporogen primer dan ke arah luar menghasilkan sel parietal primer. Sel sporogen berfungsi langsung sebagai sel induk megaspora. Sel induk megaspora (megasporosit) membelah secara meiosis membentuk 4 megaspora yang haploid dan umumnya bertipe linier, tetapi ada yang berbentuk huruf T, antara lain pada Orchic maculata dan Driniys winteri. Sedang pada beberapa suku Crassulaceae, Hydrochaitaceae dan Musaceae dilaporkan mempunyai tipe berturut turut isobilateral, tetrahidris dan bentuk T. Dari 4 inti megaspora hasil meiosis yang tersusun linier tersebut hanya satu inti megaspora yang berfungsi yaitu yang letaknya paling bawah dari tetrad, tiga lainnya mengalami degenerasi. Perkembangan gametofit betina (Megagametogenesis) Gametofit betina (kantong embrio) yang dewasa terdiri atas 7 sel, yaitu sel sentral yang besar dengan 2 inti kutub, di bagian mikrofil 2 sel sinergid dan 1 sel telur serta di bagian khalaza 3 sel antipoda. Perkembangan kantong embrio dimulai dengan memanjangnya inti megaspora yang berfungsi. Tergantung jumlah inti megaspora yang berperan dalam pembentukannya, gametofit betina (kantong embrio) mungkin bertipe monosporik, bisporik atau tetrasponik. Masing-masing kelompok tersebut mempunyai lebih dari satu tipe. (Lihat diagram). Tipe tersebut adalah sebagai berikut: 1. Monosporik Pada tipe ini inti megaspora yang berperan selama perkembangan gametofit jumlahnya satu. Tipe ini merupakan tipe normal (tipe Polygonium) Tipe kedua yaitu Oenothera*, pada tipe ini hanya terjadi 2 kali pembelahan inti megaspora, sehingga hanya ada 4 inti di bagian mikrofil. Gambar 8.17. Megasporogenesis dan perkembangan (megagametofit) tipe Normal (polygonum) pada Angiosperm. kandung lembaga 2. Bisporik Inti megaspora yang berfungsi pada perkembangan gametofit betina ada 2. Setelah meiosis pertama pada proses megasporogeilesis terbentuk 2 set, dan 2 sel tersebut hariya satu, sel yang melanjutkan meiosis II, sedang yang lain mengalami degenerasi. Pada pembelahan meiosis II tidak terjadi pembentukan dinding sekat, dan kedua inti megaspora berperan dalam pembentukan kandung lembaga. Dua inti ini kemudian membelah mitosis 3 kali, menghasilkan 8 inti. Akhirnya orgamsasi kandung lembaga seperti pada tipe normal (Polygonum). Tipe bisporik dibedakan menjadi 2 yaitu: a. tipe Allium Pada tipe ini megaspora yang berfungsi adalah yang berada di bagian khalaza, sedang yang ada di bagian mikrofil mengalami degenerasi setelah meiosis 1. b. tipe Endyinion Megaspora yang berfungsi pada tipe ini adalah yang ada di bagian mikrofil. Inti megaspora yang ada di bagian khalaza mengalami degenerasi. 3. Tetrasporik Pada tipe ini pembelahan meiosis dari sel induk megaspora selama megasporogenesis tidak diikuti oleh pembentukan dinding sekat, sehingga pada akhir meiosis 4 inti haploid tetap di dalam sitoplasma sel yang sama (terjadi pembelahan inti bebas). Pola organisasi kandung lembaga tetrasponik ini sangat bervariasi. Susunan kandung lembaga sebelum mengalami mitosis adalah sebagai berikut: a. Terdiri 4 inti yang tersusun 1+1+1+1, masing-masing ada di bagian mikrofil, khalaza dan di bagian lateral kandung lembaga. Misalnya pada tipe Peperoinia, Penae dan Plumbago. b. Terdini dari 4 inti tersusun 1+3. Satu ini bagian mikrofil 3 di bagian khalaza. Pada tipe ini 3 inti di khalaza ada yang mengadakan fusi seperti tipe Fritillaria dan Plumbagela, sedang pada tipe Drusa tidak tenjadi fusi. c. Terdiri dari 4 inti dengan susunan 2+2, dua inti di bagian mikrofil, dua inti di bagian khalaza. Inisainya tipe Adoxa. 1. Suatu tipe perkembangan kandung lembaga tetrasporik yang spesifik di jumpai pada Chrysanthemum cinerariaefolium. Perkembangan tipe ini setelah stadium 4 inti, pada akhir meiosis, dengan susunan 1+2+1. Sam inti tenletak di bagian khalaza dan mikrofil, sedang 2 inti terletak di bagian tengah. Gambar 8.19 Diagram berbagai tipe perkembangan kandung lembaga pada Angiospermae. 8.5. POLINASI DAN PEMBUAHAN 8.5.1. Polinasi Polinasi adalah jatuhnya butir pollen pada kepala putik. Pada Gymnospermae karena tidak mempunyai putik, butir pollen langsung jatuh pada nuselus. Perpindahari pollen pada Angiospermae ada 2 cara yaitu: 1. Pollen yang jatuh pada kepala putik berasal dari satu bunga yang sama. Ini disebut penyerbukan sendiri (autogaini selfpollinaiion). 2. Pollen berasal dari bunga lain, ini disebut penyerbukan silang (cross pollination). Pada tipe ini dibedakan menjadi 2: 2.1. pollen berasal dari bunga yang berbeda, tetapi sam tanaman. Penyerbukan semacam ini disebut geitonogaini 2.2. pollen berasal dari bunga 2 tanaman yang berbeda. Tipe demikian disebut xenogami. Setelah berada pada kepala putik, pollen akan berkecambah. Lama waktu yang dibutuhkan oleh pollen untuk berkecambah sangat bervariasi untuk setiap jenis tumbuhan. Langkah pertama dari perkecambahan adalah bertambahnya ukuran pollen, karena mengabsorpsi cairan yang ada pada permukaan kepala putik (Stigma), dan desakan intin melalui lubang perkecambahari. Suatu buluh kecil tumbuh memanjang, menembus jaringan stigma dan stilus (tangkai putik). Pada umumnya buluh pollen bertipe monosifonus. (Sam buluh), tetapi ada yang mempunyai buluh banyak, seperti pada Malvaceae, Cucurbitaceae dan Campanulaceae. Keadaan ini disebut polisifonus. Pada Althaea rosea mempunyai 10 buluh pollen, sedang pada Malva neglecta 14 buluh. Stigma merupakan bagian yang berperanan penting dalam perkecambahan pollen. Setelah buluh muncul dari butir pollen, buluh tersebut mencari jalan pada permukaan papila stigma, misalnya pada Gossypium atau melalui lapisan dinding stigma yang sel-selnya terdiri atas pektoselulosa misalnya pada Lilium, ke dalam jaringan stilus. Dinding buluh pollen terdiri atas 3 lapisan yaitu terluar terdiri atas pektin, lapisan tengah dan pektoselulosa, dengan struktur fibliler yang kaya akan - 1,4 linked glucan. Sitoplasma pada buluh kaya akan mitokondria dan badan Golgi, Retikulum endosplasma halus dan kasar, vesikel , amiloplas dan badan lipid. Vesikel kaya akan polisakanda atau RNA. Berdasarkan keadaan morfologi ada 3 tipe stilus: 1. tertutup ; banyak dijumpai terutama pada tumbuhan dikotil. 2. terbuka ; dijumpai adanya saluran stilus yang lebar (tidak ada jaringan transinisi), epidennis berfungsi nutritif. Sel-sel saluran stilus diselubungi oleh zona sekretoris. 3. setengah tertutup; saluran stilus tidak lebar dikelilingi oleh jaringan transinisi yang rudinienter terdiri atas 2-3 lapisan sel kelenjar (sekresi). Gambar 8.20. Tipe- tipe stilus pada Angiospermae A. Potongan bujur pistihini B. Potongan bujur bagian atas dan stigma C. Potongan bujur stilus tipe terbuka D. Potongan lmtang stilus tipe tertutup. E. Serbuk sari yang telah berkecambah. Waktu yang dibutuhkan buluh pollen untuk mencapai kandung lembaga setelah polinasi sampai terjadmya pembuahari untuk setiap jenis tumbuhan bervariasi. Quercus membutuhkan waktu 12-14 bulan Alnus glutinosa & Corylus avellana 3-4 bulan Paphiopedium mandiae 19-20 ininggu Orchis maculata 14 hari Carica papaya 10 hari Carya illinoensis 4-7 hari Oryza sativa, Coffea arabica 12-14 jam Crepis capillaris 60 menit Taraxacum kok-saghys 15-45 menit 8.5.2. Pembuahari Setelah berkecambah, buluh menembus jaringan stilus (pada tipe tertutup) atau membuat jalan pada permukaan epidermis yang membatasi saluran stilus (pada tipe terbuka) yang kemudian masuk ke dalam janingan stilus. Akhirnya buluh sampai di dalam ovarium, dan segera menuju ovulum. Masuknya buluh pollen ke dalam ovulum kemungkinan secara: 1. poligami, ini merupakan cara yang umum, yaitu buluh melalui mikrofil. 2. khalazogaimi, buluh melalui ujung khalaza, misalnya pada Casuarina. 3. misogami, buluh masuk melalui funikulus misalnya Pistacia, atau melalui integumen seperti pada Cucurbita. Gambar 8.21. Skema Pola masuknya buluh pollen ke dalam ovulum (keterangan: baca teks) Buluh pollen yang membawa sperma, setelah sampai di mikrofil masuk ke dalam kandung lembaga dengan 3 cara yaitu: 1. buluh pollen masuk di antara dmding sel telur dan dinding sinergid. 2. antara dinding kandung lembaga dan sam sel sinergid. 3. langsung masuk ke dalam salah satu sel sinergid. Kalau langsung masuk ke dalam sel sinergid, buluh menembus aparatus fihiforinis, kemudian ujung buluh pecah, isi sel buluh (sitopasma, inti vegetatif dan sel sperma) keluar, bergabung dengan sitoplasma sel sinergid. Dua sel sperma berubah bentuk, kemudian keluar dari sel sinergid. Satu sel sperma menuju sel telur, dan yang lain mendekati sel sentral (sel kutub) sel sinergid kemudian mengalami degenerasi. Telah dilakukan penelitian, dengan pengecatan khusus ada 2 badan yang berwarna gelap didalam sel sinergid dan badan tersebut dinamakan badan x. Menurut Jensen (1972) telah ditetapkan bahwa satu diantaranya adalah sisa inti sinergid dan yang lain sisa inti vegetatif, karena mengandung DNA. Badan x setelah sperma masuk ke dalam sel telur terjadilah fusi antara inti sel telur dengan inti sperma. Ini disebut singami. Sperma yang lain berfusi dengan sel sentral. Peristiwa ini disebut fusi tripel (tripel fusion). Dengan adanya dua macam pembuahari tersebut pada Angiospermae dikenal dengan pembuahan ganda (double fertilization). Suatu keadaan yang menyimpang, dimana banyak buluh pollen yang masuk masing-masing membawa 2 sperma, atau lebih dan sperma dalam satu buluh pollen masuk ke dalam kandung lembaga. Hal ini akan menyebabkan terjadinya polispermi. Polispermi adalah suatu keadaan dimana satu sel telur dibuahi lebih dan satu gamet Hasil peleburan (fusi) sel gamet jantan dengan sel telur adalah zigot, dan sel gamet jantan dengan inti kutub adalah endosperm. Endosperm pada umumnya berkembang lebih dahulu dari pada zigot. Fungsi endosperm memberi makan embrio. Ploidi endosperm pada Angiospermae adalah 3n sedang pada Gymnospennae n (haploid). Gambar 8.24. Pembuahan ganda pada Lilium martagon A. Kandung lembaga yang masak; B. Buluh serbuk sari (bs) masuk ke dalam kantong embrio yang masak; salah satu sperma mendekati inti telur, dan yang lain mengadakan kontak dengan inti kutub. Salah satu inti sinergid mengalami degenerasi (d); C. Inti sperma mengadakan kontak dengan inti telur dan sel sentral; D. Perkembangan lebih lanjut dan pembuahari; E-H. Fusi antara inti telur dengan sperma; I-N. (fusi antara inti sperma dengan kedua inti kutub (tripel fusion). 8.6 Endosperm Pada umumnya endosperm merupakan hasil pembelahan sel endosperm primer secara mitosis berkali-kali, dan berfungsi memberi makan embrio yang sedang berkembang. Tidak semua golongan tumbuhan mempunyai endosperm. Tumbuhan yang tidak mempunyai endosperm adalah suku Orchidaceae, Podostemaceae dan Trapaceae. Derajat ploidi (jumlah kromosom) endosperm bervariasi tergantung pada jumlah inti megaspora yang berfungsi pada pembentukan gametofit betina. Endosperm pada kebanyakan tumbuhan mempunyai derajat ploidi 3 (tripolid). Ploidi pada endosperm haustonum pada Thesium alpinum lebih dari 384 n. Yang mempunyai ploidi sangat tinggi adalah endosperm Arum maculatum, yaitu 24576 n. Terjadinya poliploidisasi pada endosperm disebabkan karena penistiwa endomitosis dan fusi inti di dalam selsel endosperm (Kapoor, dalam Bhojwarn dan Bhatnagar, 1978). Sel-sel endosperm biasanya berbentuk isodiametris, di dalamnya terdapat butir-butir amilum, lemak, protein, atau butir-butir aleuron. Pada serealia, beberapa lapisan endosperm yang terluar menjadi terspesialisasi baik secara morfologi maupun fisiologi, dan menyusun suatu jaringan aleuron. Pada gandum jaringan aleuron terdiri atas 3-4 lapis sel. Pada waktu biji masak, lapisan aleuron masih tetap hidup, dan bagian sel yang mengandung amilum (endosperm) dikelilingi oleh lapisan aleuron. Selsel aleuron mempunyai dinding tebal, inti besar dan sitoplasma tidak bervakuola. Pada dikotil aleuron tidak merupakan lapisan, tetapi merupakan butir-butir yang terdapat di dalam sel endosperm. Misalnya pada Ricinus communis, Vicciafaba dan lain-lain. Apabila di dalam biji tidak dijumpai adanya endosperm, fungsi nutritif bagi embrio yang sedang berkembang diambil alih oleh jaringan yang ada di dalam ovulum. Pada suku tertentu, antara lain Amaranthaceae, Cannaceae, Piperaceae dan Cappatidaceae, jaringan nuselus dapat berfungsi sebagai tempat cadangan makanan. Jaringan nuselus ini disebut perisperm. Pada Piper nigrum jaringan nuselus di bawah kantong embrio membelah, dan aktivitas pembelahannya terus bertambah. Sel-sel tersebut adalah perisperm. Perisperm dan epiderinis nuselus banyak mengandung amilum, sedang endospermnya sendiri yang terdapat di sekitar embrio sangat mereduksi bila dibanding perisperm. Jumlah amilum terus bertambah mulai dan khalaza sampai ke bagian mikrofil, sehingga 90% bagian dan biji Piper nigrum ini ditempati oleh perisperm. Pada biji Myristica fragans (pala) endosperm dan perisperm berkembang sama kuat. Pada Cyanastrum endosperm dan sebagian besar nuselus tidak kelihatan selama perkembangan biji. Tetapi sel-sel nuselus yang ada di bagian khalaza, tepat di atas janingan vaskular aktif mengadakan pembelahan membentuk jaringan yang disebut khalasosperm. Sel-sel jaringan ini penuh dengan lemak dan amilum, berfungsi sebagai pengganti endosperm. Berdasarkan perkembangannya, endosperm dibedakan menjadi 3 tipe yaitu: 1. nuklear. Pada tipe ini pembelahan inti endosperm primer (secara mitosis) yang pertama serta pembelahan selanjutnya tidak diikuti oleh pembentukan dinding sekat, sehingga terjadi inti bebas. 2. seluler. Pembelahan pertama dan pembelahan selanjutnya inti endosperm primer diikuti oleh pembentukan dinding sekat. Di sini kantong embrio terbagi dalam ruangan-ruangan, walaupun di antaranya ada yang mengandung lebih dari satu inti. Misalnya pada Peperomia. 3. helobial. Tipe ini intermediar antara tipe pertama dan tipe kedua. Misalnya pada Helobiae, Zea mays atau Oryza sativa. Menurut data yang dikumpulkan oleh Davis (1966) 288 famili dalam Angiospennae: 161 famili tipe endosperm nuklear, 72 famili tipe seluler, harinya 17 famili bertipe helobial. Seluler merupakan tipe umum yang di jumpai pada tumbuhan dikotil, pada monokotil hariya pada famili Araceae dan Lemnaceae. Dan 17 famili yang bertipe helobial, 14 famili adalah monokotil. Pada beberapa famili endosperm di bagian khalaza sering mengalami perubahari menjadi haustorium. Gambar 8.25. Endosperm tipe helobial pada Asphodelus temafolius A-B. 2 sel endosperm, 1 sel yang besar adalah ruang mikrofil, yang kecil ruang khalaza. C-D. Pada ruang mikrofil terdapat banyak inti, sedang di bagian khalaza terdapat 4 inti, zigot belum membelah; pembelahan inti tidak diikuti oleh pembentukan dinding. E. Terjadi selulerisasi di bagian penfer niang mikrofil, sedang di ruang khalaza tetap terjadi periode inti bebas. F. Perbesaran gambar E di bagian khalaza. Gambar 8.26. Endosperm tipe nuklear pada Acalypha indica A. Kantong embno setelah pembuahari, inti primer dan zigot belum membelah. B-C. Pembelahan inti endosperm, menunjukkan periode inti bebas (tanpa dinding sekat). D. Inti endosperm telah pindah ke bagian tepi kantong embno. E. Terjadi selulerisasi pada akhir perkembangan. Pembentukan dinding sekat biasanya terjadi secara sentnpetal (dan bagian tepi ke bagian tengah). A. Endosperm mengalami selulerisasi secara sempuma, dan embno berkembang menjadi stadium jantung. Perkembangan endosperm pada Loranthaceae adalah unik. Tumbuhan ini ovulumnya tidak mempunyai integumen (ateginik), sehingga kandung lembaga terdapat di dalam ovarium, dan hubungan antara ovulum satu dengan lainnya adalah parenkim penghubung. Pada perkembangannya endosperm di dalam kandung lembaga mengadakan fusi, membentuk endosperm majemuk. Gambar 8.27. Endosperm majemuk pada Tolypanthus involucratus A. penampang lintang ovarium dengan 4 kelompok endospenn. B. Endosperm masing —masing kelompok berfusi, menjadi satu membentuk struktur majemuk. Pada endosperm yang telah dewasa kadang terjadi keadaan yang menyinipang, diniana sel-sel endosperm mengadakan alctivitas pertumbuhan ke arah luar atau dalam yang tidak teratur. Atau kemungkinan kulit biji yang mengadakan pertumbuhan tidak teratur, sehingga menyebabkan perubahari struktur endosperm. Keadaan ini yang menyebabkan endosperm dikatakan bertipe ruminat. Misalnya pada pala Myristicafragran dan Cocolaba. Gambar 8.28. Endosperm ruminat pada biji A. permukaan biji tampak berlekuk-lekuk B. potongan melintang biji, perhatikan struktur endosperm 8.7. EMBRIO Telur yang telah dibuahi disebut zigot, dan ini merupakan sel tunggal yang bersifat diploid. Polaritas embno pada Angiospermae adalah endoskopik, yaltu berlawanan dengan mikrofil. Pembelahan zigot yang pertama kali pada kebanyakan Angiospermae dengan dinding melintang, sehingga menghasilkan proembrio 2 sel. Dan proembrio 2 sel ini; sel a (ca), sel bagian atas disebut terminal (sel apikal) merupakan sel yang jauh dan mikrofil. sel b (cb), sel bagian bawah disebut sel basal, adalah sel yang letaknya dekat dengan mikrofil. Selain dengan dinding melintang pembelahan zigot dengan dinding tegak lurus pada suku Loranthaceae atau miring (Triticum sp.). Pembelahan dengan dinding miring jarang. Variasi pola perkembangan embrio pada awal embriogeni merupakan hal umum pada tumbuhan monokotil maupun dikotil. Dan stadium 2 sel sampai stadium diferensiasi biasanya disebut proembrio. Perkembangan awal proembrio pada monokotil dan dikotil adalah sama sampai stadium oktant (8 sel). Perbedaannya tampak pada saat awal terbentuknya kotiledon dan plumula. Berdasarkan cara pembelahan sel apikal (ca) proembno 2 sel dan peranan sel basal (cb) serta sel apikal pada pembentukan embrio selanjutnya, maka Maheswari (1950) membagi 5 tipe perkembangan embrio pada tumbuhan dikotil sebagai berikut: A. Sel apikal dan proembrio 2 sel membelah secara longitudinal. 1. Sel basal berperan sedikit atau tidak sama sekali pada perkembangan embrio selanjutnya. Tipe Cruciferae/ Onagraceae. 2. Sel basal dan sel apikal beiperan dalam perkembangan embrio selanjutnya. Tipe Asteraceae. B. Sel apikal dan proembrio 2 sel membelah secara transversal. 1. Sel basal hanya sedikit berperan atau tidak sama sekali pada perkembangan embrio selanjutnya. 1.1. Sel basal biasanya membentuk suspensor. 1.2. Sel basal tidak mengadakan pembelahan selanjutnya, bila ada suspensor, supensor berasal dari sel apikal Tipe Cariyophylaceae 2. Sel basal dan sel apikal berperan dalam perkembangan embrio selanjutnya. Tipe Chenopodiaceae. Menurut Joharisen (1950) dikenal tipe ke 6 yaitu tipe Piperaceae misalnya pada suku Piperaceae dan Loranthaceae. Tipe ini didasarkan atas pembelahan zigot pertama kali dengan dinding vertikal (tegak lurus). Suspensor Merupakan bagian embrio yang letaknya berdekatan dengan ujung radikula. Perkecambahan suspensor mencapai maksimum pada saat embrio mencapai stadium bulat (globular). Pada biji yang masak sisa-sisa suspensor menunjukkan variasi dalam bentuk, ukuran serta sel yang menyusunnya. Variasi ini biasanya berhubungan dengan fungsi nutritif bagi embrio. Pada tumbuhan yang tidak mempunyai endospenn, suspensor bersifat haustorium. Dikatakan pula selain membantu memberi makan, suspensor merupakan akar embrionik yang bersifat sementara. Menurut Sussex et al. (1973), sel-sel suspensor pada Phaseolus coccineus banyak mengandung RNA dan protein. Struktur embrio Setelah pembuahan zigot membelah berkali-kali menjadi embrio. Embrio ini mempunyai potensi untuk membentuk tanaman yang sempurna. Embrio mempunyai poros embrional. Poros (sumbu) embrional pada dikotil menyebabkan terjadinya dua kutub, yaitu kutub yang ada di bagian atas yaitu epikotil dan yang ada dibagian bawah hipokotil. Epikotil akhirnya menjadi pucuk embnônik (plumula), dan hipokotil akan menghasilkan batang sedang pada bagian bawah hipokotil akan menghasilkan calon akar. Pada umumnya embrio dikotil dan monokotil mempunyai persamaan perkembangan sampai stadium 8 sel, yaitu stadium bulat. Embrio pada monokotil bentuknya silindris karena mempunyai satu kotiledon, sedang pada dikotil mungkin biobus (2 lobi) karena mempunyai dua kotiledon. Kotiledon pada dikotil muncul sebagai dua tonjolan meristematik pada ujung apikal embrio. Tonjolan ini disebabkan adanya perluasan ujung apikal embrio ke arah lateral. Karena adanya dua kotiledon ini maka embrio terbelah secara bilateral simetris. Bagian apeks yang terdapat pada lekukkan di antara dua kotiledon menyusun suatu meristem apikal (shoot). Diferensiasi kutub atas sudah ditentukan mulai dari awal, jauh sebelum embrio mencapai ukuran yang maksimum. Meristem yang ada di kutub atas adalah protoderm, prokambium dan meristem dasar. Sedang diferensiasi kutub bawah meliputi organisasi meristem ujung akar dan tudung akar (root). Meristem ujung akar ini mirip sekali dengan titik tumbuh ujung batang, dalam hubungannya dengan pembentukan jaringanjaringan primier. Embrio pada monokotil berbeda dengan dikotil, karena selain jumlah kotiledon, juga berbeda dalam struktur. Kotiledon pada monokotil dinamakan skutelum. Pada potongan membujur embrio dapat dilihat adanya sumbu embrional. Sumbu embrional bagian bawah dan skutelum adalah radikula (calon akar) yang menghasilkan meristem ujung akan dan tudung akar. Radikula dan tuding akar diselubungi oleh selaput pelindung yang disebut koleonza. Epikotil menyusun tunas apeks dengan primordium daun. Epikotil bersama primordium daun diselubungi oleh koleoptil. Disisi lateral koleoriza membentuk tonjolan kecil ke arah luan, dan tonjolan ini disebut epiblas. Pada beberapa tumbuhan yang endospermnya tidak berkembang, embrio berfungsi sebagai penyinipan makanan cadangan sehingga embrio menjadi tebal. Misalnya pada tumbuhan Leguininosae. Sedang pada biji yang endsopermnya berkembang embrio sangat tipis. e. endosperm; en. Endotelium; ep. epiblas; k. kotiledon; kh. koleoriza; ko. Koleoptil; la. Lapisan aleuron; m. meristem apikal; n. nuselus ; p. prokambium; r. radikula; s. skutelum; t. tunas pucuk. (dikutip dan Esau, 1978). 8.8. BUAH DAN BIJI 8.8.1. Buah Berdasarkan derajat kekerasan perikarpium (dinding buah) buah dibedakan ke dalam dua tipe, yaitu buah kering dan buah berdaging. Pada buah yang berdaging, perikarpium, yang berasal dari dinding ovarium terdiferensiasi menjadi epikarpium, mesokarpium dan endokarpium. Endokarpium biasanya keras dan mengandung sel baru. Pada buah kering perikarpium sering mempunyai jaringan sklerenkiniatis. Penggolongan buah yang lain didasarkan pada tingkat kemampuan buah untuk membuka (merekah) atau tidak pada waktu masak. Perkembangan buah Secara normal perkembangan buah terjadi setelah pembuahan. Bakal buah meluas ke arah plasenta dan ovarium. Bertambahnya ukuran buah disebabkan oleh adanya 2 proses, yaitu pembelahan sel (yang diawali oleh membesarnya sel, sebelum pembelahan mitosis) dan pembesaran sel selanjutnya. Biasanya awal terjadmya pembesaran sel tergantung pada pembelahan sel, dan dimulai sebelum antesis, kemudian berlanjut sampai buah nyata. Tingkat ini kemudian secara berangsur diganti dengan pembentangan sel, dan diikuti oleh pertumbuhan memanjang. Periode tingkat perkembangan buah berbeda-beda dan diikuti pula oleh pertumbuhan komponen buah seperti perikarpium, kulit biji, endosperm dan embrio. Faktor yang mempengaruhi perkembangan buah adalah faktor dalam dan faktor luar. Salah satu faktor dalam adalah perkembangan biji. Struktur buah Apabila bakal buah berkembang menjadi buah, dinding ovarium menjadi perikarpium. Dinding ovarium terdiri dari sel-sel parenkim, jaringan pembuluh dari lapisan epidermis dalam dan luar. Selama pemasakan, perikarpium bertambah jumlah selnya. Jaringan dasar secara relatif tetap homogen dan parenkim terdiferensiasi menjadi parenkim dan jaringan sklerenkim. Perikarpium mungkin terdiferensiasi menjadi 3 bagian yang secara morfologi berbeda yaitu eksokarpium (lapisan terluar), mesokarpium (bagian tengah), dan endokarpium (lapisan terdalam). Kadang-kadang eksokarpium dan endokarpium merupakan epiderinis luar dan epiderinis dalam dinding ovarium. Dinding ovarium menyelubungi ovarium dimana biji dihasilkan. Struktur jaringan pembuluh bervariasi untuk setiap jenis buah dan terdapat pada perikarpium. Struktur perikarpium menunjukkan variasi yang luas untuk setiap jenis atau tipe buah. Ada 2 macam tipe perikarpium, yaitu parenkiniatik, pada buah berdaging dan sklerenkimatik pada buah kering. Pada buah polongan, pada waktu buah masak karpel memisah sepanjang sutur atau kampula yang mengelilingi buah, meninggalkan biji yang melekat pada rusuk dan membentuk suatu kerangka di sekitar sekat. Buah pisang (Musa acuininala) mempunyai tipe ovarium inferior, dengan 3 karpel. Ovarium ini kemudian sebagai buah yang mempunyai biji, atau buah tanpa biji (partenokarpi). Buah yang berbiji/partenokarpi mempunyai struktur sama pada awal perkembangan. Akhirnya ovulum pada buah partenokarpi mengalami degenerasi, dan lokulus ditutupi oleh daging buah yang berasal dari perikarp dan sekat. Daging buah kaya akan amilum. Pada varietas yang berbiji, biji yang masak hampir memenuhi lokulus, dan daging buah sangat tipis. Ikatan pembuluh bersama dengan lateks, terselubung dalam jaringan parenkini dinding buah. Berbeda dengan buah pisang, buah tomat (Lycopersicon esculentum), mempunyai jumlah karpel yang banyak, janingan berdaging terdiri atas perikarpium, sekat dan plasenta. Jaringan plasenta meluas, memasuki ruang-ruang antara ovulum. Plasenta menutup lokulus, dan terselubung oleh ovulum. Jaringan antara ovulum berisi gelatin pada waktu buah masak. Perubahari warna kulit buah selama pemasakan disebabkan adanya transfonnasi kloroplas menjadi kromoplas. 8.8.2. Biji Setelah pembuahan bakal biji akan berkembang menjadi biji. Integumen berkembang menjadi kulit biji atau testa, sel telur yang dibuahi (zigot) berkembang menjadi embrio, dan sel endosperm primer akan membelah- belah secara mitosis menghasilkan endosperm. Bentuk, ukuran, warna, struktur dan permukaan biji sangat bervariasi. Struktur biji a. Kulit biji Merupakan bagian terluar biji. Pada Angiospermae bakal biji mempunyai satu atau dua integumen. Pada umumnya semua bagian yang menyusun integumen berperan dalam pembentukan kulit biji. Sening pada biji tertentu jaringan integumen mengalami kerusakan karena adanya perkembangan jaringan lain pada biji, sebingga kulit biji berasal dari bagian yang tersisa di dalam integumeti. Gossypium sp. mempunyai ovulum yang biteginik, dan ke dua integumen berperan dalam pembentukan kulit biji. Perubahan-perubahan histologis tampak jelas 6 hari setelah pembuahan. Struktur anatoini kulit biji sangat bervaniasi untuk setiap jenis tumbuhan. Sel-sel parenkim pada integumen mengalami diferensiasi menjadi aerenkim sel-sel cadangan makanan, sel-sel tanin, sel kristal, sel gabus, sel sklerenkim, dan lain- lain. Mengenai susunan kulit biji pada umumnya adalah: 1. di sebelah luar terdapat epidermis, atau sering tanpa epidermis; 2. di sebelah dalam lapisan epidermis adalah jaringan yang sel-selnya berdinding tebal, mempunyai ukuràn yang panjang, tersusun seperti jaringan tiang pada daun, disebut jaringan palisaden atau dikenal sebagai makrosklereida; 3. di sebelah dalam lapisan ini mungkin masih dijumpai adanya jaringan yang selselnya berdinding tebal disebut jaringan osteoskiereida; 4. selanjutnya di jumpai sel-sel parenkim, sel-sel kristal atau sel-sel yang mengandung pigmen. Pada permukaan kulit biji, pengamatan dengan menggunakan inikroskop elektron skaning menunjukkan adanya ornamentasi pada kulit biji yang bermacammacam bentuknya. Tergantung pada ada atau tidaknya endosperm pada biji, maka dibedakan 2 tipe yaitu: 1. Endospermus (albuininus) Pada biji dijumpai adanya endosperm. Misalnya pada : Zea mays, Ricinus communis, dli. 2. Non endospermus (eks-aibuininus) Pada biji tidak dijumpai adanya endosperm. Misalnya pada : Areca catechu, Piper nigrum, Glycine max, Cucurbita, dli. Pada biji yang masak sering masih dijumpai adanya arilus atau karunkula arilus (salut biji) dijumpai pada Punica granatum, Nephelium, dll, sedang karunkula pada biji Ricinus communis. Gambar 8.34. Struktur umum kulit biji beberapa spesies tumbuhan Angiospermae A. Gymnocladus diolea; B. Viola tricolor; C. Phaseolus multfiorus; (lapisan dalam yang berbatu hariya tampak 1/5 bagian) D. Magnolia macrophylla; E. Plantago lanceolata; F. Lepidium sativum; G. Vaccinium corrymbosum (epidermis dengan sel yang besar, lapisan dalam berlendir); H. Maluspuinila. c. dikotiledon; e. endosperm; i. Integumen dalam; 11. Jaringan palisade; n. nuselus; 0. integumen luar. (dikutip dan Eames & MacDaniels, 1953) Gambar 8.35. Perkembangan kulit biji pada Gossypium sp. A. Penampang bujur ovulum. Ovulum mempunyai 2 integumen. B. Struktur anatomi integumen luar dan dalam. C-D. Integumen dalam dan luar dari ovulum pada saat 2-3 hari dan 5-6 hari setelah pembuahan. Setelah terjadi pembuahan semua jaringan yang menyusun integumen mengadakan diferensiasi, dan natinya menyusun kulit biji. E. Kulit biji (15 hari setelah pembuahan); F. Kulit biji dewasa; G- H. Trikoma (rambut) yang biasanya terdapat pada permukaan kulit biji merupakan derivat epidermis integumen luar. ii. Integumen luar; n. nuselus; ke. Kantong embrio; id. Integumen dalam; it. Initial trikoma; t. trikoma; sp. sel berpigmen; st. Se! tidak berpigmen. Gambar 8.36. Diagram struktur anatomi biji A. Biji Zea mays; B. Biji Glycine max; C. Biji Piper nigrum seluruh biji ditempati perisperm. Perisperm lebih berkembang daripada endospem. 8.3. POLIEMBRIONI Poliembrioni adalah terdapatnya lebih dari satu embrio dalam satu biji. Orang yang melaporkan pertama kali, terjadinya poliembrioni adalah Antom van Leeuwenhoek pada tahun 1719, pada biji jeruk. Poliembrioni pada Angiospermae kemungkinan terjadi karena: 1. pembelah embrio yang sudah ada (Cleavage pro-embryo). 2. embrio berasal dari sel-sel dalam kandung lembaga selain sel telur yang dibuahi. 3. terbentuknya kandung lembaga yang banyak, dalam satu ovulum. 4. aktivitas sel-sel sporofilik (sel-sel sama) pada ovulum. 1. “Cleavage polyembtyony pada Angiospermae dijumpai pada anggrek, seperti Eulophia epidendraea (lihat gambar terlampir). 2. embrio berasal dari sel-sel dalam kandung lembaga selain sel telur yang dibuahi. (lihat gambar). Pada gambar dapat dilihat embrio berasal dari sel antipoda. Klasifikasi poliembrioni Ada 2: 1. Spontan 2. Induksi Ernst (1901; 1910) membedakan poliembrioni spontan menjadi 1. Poliembrioni sejati Dua atau lebih embrio terdapat dalam satu kantong lembaga. 2. Poliembrioni palsu Embrio terdapat dalam kantong embrio, pada satu ovulum (Fragaria) atau plasenta (Loranthaceae). Gambar 8.37. Poliembrioni pada Eulophia epidendraea A. zigot membentuk kelompok sel, 3 diantaranya membelah membentuk embrio yang bebas B-C. dan embrio yang terbentuk tumbuh tunas (cabang) pada bagian sisi embrio dan masing-masing cabang akan tumbuh menjadi embrio.