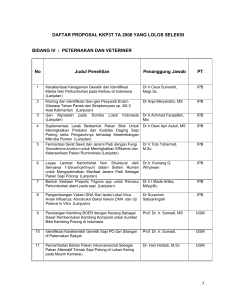
Studi keragaman fenotipik dan genetik beberapa sub populasi
advertisement

STUDI KERAGAMAN FENOTIPIK DAN GENETIK BEBERAPA SUB POPULASI KAMBING LOKAL INDONESIA DAN STRATEGI PEMANFAATANNYA SECARA BERKELANJUTAN ARON BATUBARA SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2011 SURAT PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI Dengan ini saya menyatakan bahwa disertasi yang berjudul “Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan” adalah karya saya dengan arahan Komisi Pembimbing, dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini. Bogor, … …Juli 2011 Aron Batubara NRP. D161070041 ABSTRACT ARON BATUBARA. Study of Phenotypics and Genetics Diversity of Some Indonesian Local Goat Sub Populations and Its Strategies for Sustainable Use. Supervised by RONNY RACHMAN NOOR, ACHMAD FARAJALLAH and BESS TIESNAMURTI. The genetic resources of local goats are very important because it was well adapted to the local environment with low input management systems. Local goats are very potential for breeding stock program which suitable to the Indonesia local conditions. The study was carried out in order to describe the phenotypic and genetic of 6 sub-populations of Indonesian local goats based on the analysis of the morphometrics, mitochondrial DNA, Y Chromosome and GDF9 gene. The sampling was conducted in four of the Provinces in Indonesia, namely North Sumatra Province (Samosir goat n=42 in the Samosir District, Muara goat n=34 in the North Tapanuli district, Kacang goat n=217 in the Deli Serdang District), Central Java Province (Jawarandu goat n=94 in the Blora District); Province of South Sulawesi (Marica goat n=60 in the Maros District, Makassar City, Jeneponto District) and Nusa Tenggara Timur Province (Bengala goat n=96 in the Kupang area, Sikka District, Ende District). All of them were measured for morphometric and was bleeding to collect blood as DNA sources. The blood sample was preserve with absolute ethanol. DNA extracted was analyzed by PCR-RFLP methods for D-loop region, SRY gene and GDF gene on Indonesian local goat does. If there were polymorphism then the procedure followed by the sequencing methods. The results showed that the morphometric discriminant factor variables between local goat were canon circumference, tail width and body length was 0,7 and than rump width, ear widths, wither height, skull height, skull width, body weight, chest girth, chest width and chest depth was 0,5. The colours and colour patterns of Benggala goat was dominant with black colour, Samosir goat with white colour and then the Kacang, Muara, Jawarandu and Marica goats were very high varieties of the combination almost all colours were observed. There were found 50 polymorphic sites and 19 unique haplotypes of the D-loop sequence. The average genetic diversity region of mt DNA was very high (0.014±0.002) and was significantly different between each sub-population with clustering indexs 63-99. The average distance of Y chromosome was lower (0.004±0.002) and to be founded 6 polymorphic sites with 4 unique haplotypes. The origin of local goats reveal to the maternal ancestors was including haplogroups lineage B, but the paternal origin ancestor was classified in to 4 groups; Kacang and Jawarandu one haplotype, Marica and Samosir one haplotype, Muara one haplotype and Benggala goat one haplotype. Based on the phenotypics, mitochondrion and Y chromosom analysis showed that the sub populations of Indonesian local goat as adistinct breed. Identification of the GDF9 gene promotor were polymorphic and have related with the prolific traits on the twinning does of Kacang and PE goats, but the GDF9 gene promotor were monomorphic on the Samosir and Muara goats. Key Words: Indonesian local goat, morphometric and genetic characterization, mitochondrion, Y chromosome, fecundity gene RINGKASAN ARON BATUBARA. Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan. Dibimbing oleh RONNY RACHMAN NOOR, ACHMAD FARAJALLAH, dan BESS TIESNAMURTI Keragaman sumberdaya genetik ternak kambing merupakan modal dasar untuk meningkatkan produktivitas dan mengembangkan ternak kambing. Populasi kambing lokal bisa terancam habis antara lain disebabkan oleh program persilangan atau penggantian dengan bibit unggul eksotis. Pelestarian dan konservasi kambing lokal penting karena telah beradaptasi baik dengan lingkungan setempat, umumnya lebih bertahan hidup pada kondisi pedesaan. Kambing lokal berpotensi besar untuk dimanfaatkan menjadi sumber pembentukan bibit unggul yang adaptif terhadap kondisi lokal di Indonesia. Data dan informasi tentang karakterisasi fenotipik dan genetik ternak kambing lokal Indonesia sampai saat ini masih sangat terbatas. Tujuan penelitian ini adalah untuk memperoleh informasi keragaman karakteristik fenotip dan genetik 6 sub-populasi kambing lokal Indonesia yang mencakup karakteristik morfometrik dan warna dominan; keragaman genetik berdasarkan DNA mitokondria, koromosom Y dan keragaman gen GDF9. Penelitian dirancang dengan metode survei dan analisis di laboratorium. Metode survei menggunakan purposive sampling, yaitu dengan memilih lokasi yang diduga masih terdapat populasi kambing lokal yang diamati. Penelitian lapangan untuk koleksi data fenotipik dilakukan di empat Propinsi, yaitu Propinsi Sumatera Utara (Kambing Samosir n=42 ekor di Kabupaten Samosir , Kambing Muara n=34 di Kabupaten Tapanuli Utara, Kambing Kacang n=217 di Kabupaten Deli Serdang); Propinsi Jawa Tengah (Kambing Jawarandu n=94 di Kabupaten Blora); Propinsi Sulawesi Selatan (Kambing Marica n=60: Kabupaten Maros, Kota Makassar, Kabupaten Jeneponto) dan Propinsi Nusa Tenggara Timur (Kambing Benggala n=96 di Kota Kupang, Kabupaten Kupang, Kabupaten Sikka, Kabupaten Ende). Dilakukan pengambilan data warna tubuh dominan dan warna belang kambing, penimbangan, pengukuran bagian-bagian tubuh dan pengambilan sampel darah. Analisis morfologi dengan pendekatan tehnik diskriminan dan kanonikal untuk menduga hubungan filogenik antar kambing lokal. Analisis diskriminan dilakukan pada bobot badan, bagian-bagian tubuh dan warna dominan. Kegiatan laboratorium terdiri dari ekstraksi DNA mengikuti metode Sambrook yang dimodifikasi, menganalisis DNA mitokondria, kromosom Y dan gen fekunditas (gen GDF9). Hasil analisis varians menunjukkan bahwa bobot badan dan ukuran bagian-bagian tubuh dari Kambing Muara berbeda nyata (P<0.05) lebih tinggi dibandingkan dengan sub populasi kambing lokal lainnya. Berdasarkan analisis morfometrik jarak genetik paling dekat adalah antara Kambing Marica dengan Samosir (11.207) dan paling tinggi adalah antara Kambing Muara dengan Benggala (255.110). Nilai kesamaan antar individu pada setiap sub populasi paling tinggi ditemukan pada Kambing Kacang (99.28%) dan paling rendah pada Kambing Samosir (82.50%). Nilai faktor pembeda variabel ukuran morfometrik yang dominan adalah lingkar kanon, lebar ekor dan panjang badan (masingmasing 0.7), dan variabel lingkar pinggul, lebar telinga, tinggi pundak, tinggi tengkorak, lebar tengkorak, bobot badan, lingkar dada dan dalam dada (masingmasing 0.5), yang dapat digunakan untuk karakteriasi fenotipik kambing lokal di Indonesia. Hasil fenogram menunjukkan dari 6 sub populasi kambing yang berbeda di Indonesia dikelompokkan kedalam kelompok-kelompok terpisah yaitu (1) Kambing Muara (2) Kambing Jawarandu (3) Kambing Kacang (4) Kambing Benggala (5) Kambing Samosir dan (6) Kambing Marica. Dari 879 bp runutan ruas D-loop ditemukan 50 situs polimorfik dengan nilai keragaman 0.014±0.002 dan terdapat 19 kelompok haplotip. Asal-usul secara maternal keenam sub populasi kambing lokal Indonesia yang diteliti termasuk dalam kelompok garis keturunan (lineage) B mengelompok dengan kambing Asia Timur, Afrika Selatan dan Afrika Utara, Asia Selatan, Cina, Mongolia, Malaysia, Pakistan dan India. Sebanyak 13 sampel DNA kambing jantan dianalisis dan diperoleh masing-masing situs nukleotida sepanjang 773 pb. Ditemukan 6 situs polimorfik dengan nilai keragaman 0.004±0.002 dan terdapat 4 kelompok haplotip unik yaitu Kambing Kacang dan Jawarandu, Marica dan Samosir, Muara serta Benggala. Hasil identifikasi keragaman gen GDF9 ruas promotor pada induk beranak kembar bersifat polimorfik dan diduga berhubungan dengan sifat prolifik pada Kambing Kacang dan Peranakan Etawah (PE), akan tetapi bersifat monomorfik pada Kambing Samosir dan Muara. Berdasarkan persyaratan kualitatif, kuantitatif, pemetaan genetik yang diperoleh dalam penelitian ini, maka hasil penelitian ini dapat dipertimbangkan sebagai salah satu bahan dalam penetapan dan standarisasi 6 rumpun/galur kambing lokal Indonesia yang saling berbeda di tingkat lapangan atau peternak dalam rangka pengembangan dan pelestarian sumberdaya genetik kambing lokal Indonesia secara berkelanjutan. Kata kunci : Kambing lokal, karakteristik morfometrik dan genetik, DNA mitokondria, kromosom Y, gen fekunditas. © Hak Cipta milik Institut Pertanian Bogor, tahun 2011 Hak Cipta dilindungi Undang-undang Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB. Dilarang menggunakan dan memperbanyak sebagian atau seluruh Karya Tulis dalam bentuk apapun tanpa izin IPB STUDI KERAGAMAN FENOTIPIK DAN GENETIK BEBERAPA SUB POPULASI KAMBING LOKAL INDONESIA DAN STRATEGI PEMANFAATANNYA SECARA BERKELANJUTAN ARON BATUBARA Disertasi Sebagai salah satu syarat untuk memperoleh gelar Doktor pada Program Studi Ilmu Produksi dan Teknologi Peternakan SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2011 Penguji pada Ujian Tertutup : Prof. (R) Dr. Ir. Ismeth Inounu, M.S. Dr.Ir. Dedy Duryadi Solihin, DEA. Penguji pada Ujian Terbuka : Dr.Ir. Riwantoro, MM. Dr.Ir. Simon Petrus Ginting, M.Sc. HALAMAN PENGESAHAN Judul Disertasi : Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan Nama NRP Program Studi : Aron Batubara : D161070041 : Ilmu Produksi dan Teknologi Peternakan Disetujui Komisi Pembimbing Prof.Dr.Ir. Ronny Rachman Noor, M.Rur.Sc. Ketua Dr.Ir. Achmad Farajallah, M.Si. Anggota Dr.Ir. Bess Tiesnamurti, M.Sc. Anggota Mengetahui Ketua Program Studi/Mayor Ilmu Produksi dan Teknologi Peternakan Dekan Sekolah Pascasarjana Dr.Ir. Rarah R.A. Maheswari, DEA. Dr.Ir. Dahrul Syah, M.Sc.Agr. Tanggal Ujian : 25 Juli 2011 Tanggal Lulus: PRAKATA Dengan mengucapkan puji dan syukur kepada Tuhan Yang Maha Kuasa, karena berkat karunia-Nya penulis dapat menyelesaikan penelitian dan penyusunan disertasi dengan judul “Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan”. Disertasi ini sebagai salah satu syarat untuk memperoleh gelar Doktor pada Program Studi Ilmu Produksi dan Teknologi Peternakan, Sekolah Pascasarjana Institut Pertanian Bogor. Penelitian ini dilakukan atas dasar bahwa kambing lokal Indonesia sebagai salah satu kekayaan plasma nuftah ternak di Indonesia perlu di eksplorasi identitas, karakteristik dan potensi genetiknya sebelum terlanjur hilang. Informasi tentang karakteristik secara fenotipik dan genetik berguna untuk menginventarisasi kekayaan sumberdaya genetik kambing Indonesia. Dengan semakin meningkatnya kebijakan pencapaian produksi daging nasional di berbagai daerah penggunaan bibit unggul eksotis terus meningkat, sehingga pemanfaatan bibit ternak lokal semakin kurang mendapat perhatian. Oleh sebab itu perlu dilakukan penelitian karakterisasi fenotipik dan keragaman genetik yang merupakan tahap awal sebelum penelitian potensi produktivitas dan upaya pembentukan bibit unggul yang berbasis lokal, serta upaya konservasi dan pemanfaatan ternak secara berkelanjutan. Program pendidikan S3 ini didanai oleh APBN melalui program pembinaan dan pengembangan tenaga di Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian. Ketika mulai merencanakan untuk melanjutkan studi ke jenjang S3, berbagai pihak terlibat baik langsung maupun tidak langsung. Masing-masing telah memberikan sumbangsihnya, baik berupa semangat, arahan dan motivasi, sumbangan pemikiran dan materi kepada penulis. Penelitian dan disertasi ini dapat diselesaikan tentunya atas bantuan dan bimbingan dari Komisi Pembimbing. Dalam kesempatan ini, penulis menyampaikan ucapan terimakasih yang tak terhingga kepada Prof.Dr.Ir. Ronny Rachman Noor, M.Rur.Sc selaku Ketua Komisi Pembimbing, Dr.Ir. Achmad Farallah, M.Si., dan Dr.Ir. Bess Tiesnamurti, M.Sc., masing-masing selaku anggota Komisi Pembimbing yang telah meluangkan waktu mendampingi penulis dengan penuh kesabaran, memberikan saran, koreksi, arahan, bimbingan dan semangat selama penelitian hingga selesai penulisan disertasi ini. Ucapan terimakasih yang sebesar-besarnya juga disampaikan kepada Kepala Badan Penelitian dan Pengembangan Pertanian dan Sekretaris Badan Penelitian dan Pengembangan Pertanian selaku Ketua Komisi Pembinaan Tenaga, yang telah memberikan kesempatan kepada penulis untuk mengikuti Program S3, Kepala Pusat Penelitian dan Pengembangan Peternakan dan Dr.Ir. Simon Petrus Ginting, M.Sc. selaku Kepala Loka Penelitian Kambing Potong Sei Putih, yang memberikan motivasi, dorongan dan restu kepada penulis untuk melanjutkan ke jenjang Program S3 di Sekolah Pascasarjana, Institut Pertanian Bogor. Ucapan terimakasih yang sama penulis sampaikan kepada Dr.Ir. Rarah Ratih Adjie Maheswari, DEA, selaku Ketua Program Studi/Mayor Ilmu Produksi dan Teknologi Peternakan (IPTP), Prof.Dr.Ir. Cece Sumantri, M.Agr.Sc selaku Ketua Departemen Ilmu Produksi dan Teknologi Peternakan Fakultas Peternakan Institut Pertanian Bogor yang sekaligus sebagai penguji luar Komisi pada Ujian Kualifikasi Doktor dan Prof.Dr.Ir. Muladno, MSA sebagai penguji luar Komisi pada Ujian Kualifikasi Doktor, Rektor Institut Pertanian Bogor, Dekan Sekolah Pasca Sarjana IPB, Dekan Fakultas Peternakan IPB Bogor. Terima kasih yang sebesar-besarnya juga disampaikan kepada Kepala Dinas Peternakan dan Perikanan Kabupaten Tapanuli Utara, Kepala Dinas Pertanian, Peternakan dan Perikanan Kabupaten Samosir, Kepala Dinas Peternakan Kabupaten Blora Jawa Tengah, Dr. Ir. Jacob Nulik, M.Sc. sebagai Kepala BPTP Nusa Tenggara Timur, Kepala BPTP Sulawesi Selatan, Ir. Matius Sariubang MS, Ir. Daniel Pasambe, Ir Bonggas Pasaribu, Ir. Rosianna Tarigan, Ir. Deborah Kana Hau, M.Si., Drh. Wasito M.Si., Imanianto, Wagimin yang telah banyak membantu selama pengambilan data dan sampel di lapangan, Wildan Muttaqin S.Si., M.Si dan Muhammad Rizal Hasan S.Si., M.Si yang telah banyak membantu selama proses analisis molekuler di Laboratorium Fungsi Hayati dan Perilaku Hewan Departemen Biologi IPB. Rekan-rekan seperjuangan Ir. Suryana M.Si., Ir. Eko Handiwinawan M.Si, drh. Bambang Ngaji Utomo M.Sc., Ben Juvarda, SPt., M.Si., yang selalu memberikan semangat dan motivasi kepada penulis. Ucapan terimakasih penulis sampaikan kepada Tim Pengelola Beasiswa DIPA Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian yang telah memberikan beasiswa selama studi. Ucapan terimakasih juga penulis sampaikan atas sebagian bantuan dana penelitian dari kegiatan Kerjasama Kemitraan Penelitian Pertanian dengan Perguruan Tinggi (KKP3T) tahun 2010 dengan judul “Identifikasi Tiga Gen Fekunditas pada Empat Jenis Kambing Lokal (Kacang, Peranakan Etawah, Samosir dan Muara)”. Kepada istri saya tercinta Helena Lydia Tobing, S.Pt dan ketiga putri saya tersayang Ruth Amelia Batubara, Desy Margaretta Batubara dan Joice Deatri Batubara dan seluruh keluarga besar saya, terimakasih atas perhatian, kesabaran, pengertian, dorongan, pengorbanan dan doa yang diberikan selama ini kepada Penulis, serta kepada semua pihak yang telah membantu dan tidak dapat penulis sebutkan satu persatu. Akhirnya Penulis berharap semoga disertasi ini memberikan sumbangsih dan bermanfaat bagi perkembangan ilmu pengetahuan dan pembangunan peternakan di Indonesia. Bogor,……Juli 2011 Penulis RIWAYAT HIDUP Penulis dilahirkan di Desa Hutatonga, Kecamatan Sipirok (saat ini Kecamatan Arse), Kabupaten Tapanuli Selatan, Sumatera Utara pada tanggal 22 Mei 1968 sebagai anak kedelapan dari sepuluh bersaudara dari pasangan Ali Musa Batubara (alm) dan Mince Ritonga (alm). Pendidikan sarjana ditempuh pada Program Studi Produksi Ternak, Fakultas Peternakan, Universitas HKBP Nommensen Medan, lulus tahun 1992. Kesempatan untuk melanjutkan ke jenjang program magister pada Program Studi Produksi Ternak pada Departemen of Animal Production and Health, Institute of Tropical Medicine, Antwerp, Belgia tahun 1995 dan lulus tahun 1997. Pada tahun 2007 penulis melanjutkan ke jenjang program Doktor pada Program Studi/Mayor Ilmu Produksi dan Teknologi Peternakan (IPTP) dengan minat Ilmu Pemuliaan dan Genetika Ternak, Sekolah Pascasarjana Institut Pertanian Bogor, dengan beasiswa dari DIPA Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian. Penulis memulai karir bekerja sebagai staf peneliti junior pada Proyek Small Ruminant – Collaborative Research Support Programe (SR-CRSP) Pengembangan usaha ternak Domba di daerah Perkebunan Karet di Sei Putih, Medan, Sumatera Utara pada tahun 1993-1994. Pada tahun 1995 penulis diangkat menjadi Calon Pegawai Negeri Sipil pada Sub Balai Penelitian Ternak Sei Putih. Pada tahun 1997-1998 penulis bekerja sebagai Staf Peneliti pada BPTP Sumatera Utara, pada kelompok Peneliti Sumberdaya Pertanian. Pada tahun 1999-2003 bekerja sebagai Staf Peneliti pada kelompok Peneliti Budidaya Pertanian di BPTP Riau. Pada tahun 2004-2006 sebagai Staf Peneliti pada kelompok Peneliti Pemuliaan dan Genetika Ternak Loka Penelitian Kambing Potong Sei Putih. Pada tahun 2000-2003 penulis menjabat sebagai Pimpinan Proyek Agricultural Research Management Project (ARMP-II) Riau. Pada tahun 2004-2007 penulis menjabat sebagai Koordinator Perencanaan dan Program Penelitian pada Loka Penelitian Kambing Potong Sei Putih, Pusat Penelitian dan Pengembangan Peternakan, Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian. Pada tahun 2007 sebagai tenaga Pemandu PRIMATANI Badan Litbang Pertanian di Kabupaten Ogan Ilir, Propinsi Sumatera Selatan. DAFTAR ISI Halaman ABSTRAK……………………………………………………………………….. iii RINGKASAN..………………………………………………………………….. iv DAFTAR ISI…………………………………………………………………….. xvi DAFTAR TABEL……………………………………….………....................... xix DAFTAR GAMBAR…………………………………………………................. xx DAFTAR LAMPIRAN…………………………………………….…………….. xxi PENDAHULUAN Latar Belakang…………………………………………….…………….. Tujuan Penelitian…………………………………………….………….. Manfaat Penelitian……………………………………..……...... ……... Kerangka Pemikiran…….………………………………....................... 1 3 3 4 TINJAUAN PUSTAKA Sejarah Budidaya Ternak Kambing……………………………………. Keragaman Genetik Ternak……………………………………............ Pelestarian Sumberdaya Genetik Ternak…………………………….. Sumberdaya Genetik Kambing Indonesia………………..…………... Sifat Kuantitatif dan Kualitatif…………………………………………… Penanda Genetik…………….………………………………………….. DNA Mitokondria………………………………………………………… DNA Kromosom Y……………………………………………………….. Gen yang Berhubungan dengan Sifat Prolifik…… ………………….. 6 7 8 9 12 13 14 16 17 KARAKTERISTIK MORFOMETRIK DAN ANALISIS FILOGENI PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA Pendahuluan……………………………………………………………... Bahan dan Metode………………………………………………………. Waktu dan Tempat Penelitian………………………………………. Pengumpulan Sampel Kambing……………………………………. Analisis Statistik……………………………………………………... Analisis Morfometrik…………………………………………………. Hasil dan Pembahasan Bobot Badan …………………………………………………………. Parameter Ukuran Tubuh…………………………………………... Plot Penyebaran Kambing menurut Ukuran Fenotipik…………... Nilai Campuran Fenotipik antar Kelompok……………………….. Penentuan Jarak Genetik dan Pohon Fenogram………………... Peubah Pembeda Rumpun Kambing……………………………... Pola Warna Tubuh…………………………………………………... Simpulan……………………………………………………………….…. KARAKTERISASI MOLEKULER ENAM SUB POPULASI KAMBING LOKAL INDONESIA BERDASARKAN ANALISIS SEKUEN DAERAH D-LOOP DNA MITOKONDRIA Pendahuluan…………………………………….……………................ 20 21 21 22 24 25 26 27 34 36 37 39 40 43 44 Bahan dan Metode Sampel Darah Kambing Penelitian…………………………………. Ekstraksi DNA………………………………………………………… Amplifikasi DNA..……………………………………………............. Perunutan DNA………………………………………………………. Analisis Data………………………………………………................ Hasil dan Pembahasan Polimorfisme Segmen daerah D-loop DNA Mitokondria……….... Keragaman Runutan Nukleotida………………………………….... Jarak Genetik Kambing Penelitian dengan Kambing Lainnya Simpulan…………………………………………………………………… KARAKTERISASI MOLEKULER PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA BERDASARKAN ANALISIS DNA KROMOSOM Y Pendahuluan……………………………………………….……………… Bahan dan Metode Sampel Darah Kambing Penelitian……………………………….… Ekstraksi DNA………………………………………………………… Amplifikasi DNA …………………………..………………………….. Perunutan DNA………………………………………………….……. Analisis Data……………………………………………………….…. Hasil dan Pembahasan Polimorfisme DNA Kromosom Y Segmen gen SRY…………….... Frekuensi Nukleotida dan Jarak Genetik…………………………... Simpulan…………………………………………………….……………... 45 45 45 46 46 47 50 50 57 58 58 58 58 59 59 60 62 65 IDENTIFIKASI KERAGAMAN GEN GDF9 DAN HUBUNGANNYA DENGAN SIFAT PROLIFIK PADA KAMBING LOKAL INDONESIA Pendahuluan……………………………………………………………… Bahan dan Metode……………………………………………………….. Pengambilan Sampel Darah Kambing …………………………….. Ekstraksi DNA…………………………………………………………. Amplifikasi Gen GDF9…....………………………………………….. Genotiping PCR-RFLP dan sekuensing……………….…………… Analisis Data………………..…………………………………………. Hasil dan Pembahasan…………..……………………….……………… Simpulan……………………………….…………………….……………. 66 67 67 68 68 69 69 69 72 PEMBAHASAN UMUM…………………………………………………….…... 73 SIMPULAN UMUM DAN SARAN Simpulan…………………………………………………………………… Saran……………………………………………………………………….. 79 81 DAFTAR PUSTAKA……………………………………….…………………….. 82 LAMPIRAN……………………………………………………………………….. 94 DAFTAR TABEL Halaman 1 Rataan, simpangan baku dan kisaran bobot hidup dewasa 6 sub populasi kambing lokal…………………………………………………… 26 2 Rataan dan simpangan baku ukuran tinggi pundak, panjang badan, 28 lebar dada, dalam dada, dan lingkar dada kambing jantan dan betina 6 sub populasi kambing lokal ……………………………………………. 3 Rataan dan simpangan baku ukuran panjang tengkorak, lebar tengkorak, dan tinggi tengkorak jantan dan betina dewasa 6 sub populasi kambing lokal …………………….…………………………… 30 4 Rataan dan simpangan baku panjang dan lebar ekor jantan dan betina 6 sub populasi kambing lokal……………………………….…… 31 5 Rataan dan simpangan baku panjang telinga dan lebar telinga jantan dan betina dewasa 6 sub populasi kambing lokal……………. 32 6 Rataan dan simpangan baku lingkar kanon jantan dan betina 6 sub populasi kambing lokal ………………………..…………………………. 34 7 Persentase nilai kesamaan dan campuran 6 sub populasi kambing lokal ………………………………………………………………………… 36 8 Jarak genetik berdasarkan ukuran tubuh antar 6 sub populasi kambing ……..…………………………………………………………..… 37 9 Struktur kanonikal kelompok kambing dari 6 sub populasi kambing lokal berdasarkan ukuran fenotipik……………………………………... 39 10 Persentase pola warna tubuh dominan dan warna belang pada 6 sub populasi kambing lokal …………. …..………………..….............. 40 11 Jumlah haplotip berdasarkan runutan nukleotida D-loop mtDNA setiap sub populasi kambing lokal Indonesia ………………………….. 47 12 Mutasi nukleotida sebagai penciri kelompok kambing lokal Indonesia dibandingkan dengan Capra hircus (GenBank no. akses NC_005044) ………………………………………………………..…….. 49 13 Keragaman nukleotida D-loop mtDNA pada 6 kambing lokal Indonesia ………………………………………………………………….. 50 14 Jarak genetik berdasarkan runutan nukleotida pada 6 sub populasi kambing lokal Indonesia ………….………………………………….….. 52 15 Matrik keragaman nukleotida gen SRY kromosom Y pada kambing lokal Indonesia..…………………………………..…………………….… 63 16 Distribusi kambing prolifik dan non prolifik……………………………. 67 17 Jenis-jenis mutan gen GDF9 pada ternak domba dan kambing……. 71 DAFTAR GAMBAR Halaman 1 Diagram alir kerangka pemikiran penelitian………………………………. 5 2 Skema genom daerah D-loop kambing……………………………………. 16 3 Skema genom gen SRY kromosom Y dan kromosom X.………………. 17 4 Peta lokasi pengambilan sampel dan data penelitian karakterisasi enam sub populasi kambing lokal Indonesia……….…………………….. 22 5 Titik pengukuran morfometrik kambing ..…………………………..…….. 24 6 Plot penyebaran kelompok kambing berdasarkan ukuran-ukuran fenotipik pada 6 sub populasi kambing lokal …..…………………………. 35 Pohon fenogram penyebaran kelompok kambing lokal Indonesia berdasarkan ukuran fenotipik…………………...................................... 38 8 Pola warna dominan dan belang pada 6 sub populasi kambing lokal … 42 9 Struktur genom mitokondria yang diapit oleh primer AF23 dan AF22. Nomor mengacu pada Capra hircus (GenBank no. akses AF 533441).. 7 47 10 Polimorfisme runutan nukleotida daerah D-loop DNA mitokondria pada 6 sub populasi kambing lokal mengacu pada Capra hircus (GenBank no. akses AF 533441) (tiga baris pertama dibaca secara vertikal merupakan posisi nukleotida)………………………………………………. 48 11 Dendogram 6 sub populasi kambing lokal Indonesia berdasarkan ruas D-loop mtDNA ………….…………………………………………………….. 54 12 Posisi 6 sub populasi kambing lokal Indonesia dalam dendogram kambing-kambing di dunia berdasarkan ruas D-loop ………..…………... 55 13 Median joining network dari 19 haplotip nukleotida daerah D-loop DNA mitokondria pada 6 sub populasi kambing lokal Indonesia…….............. 57 14 Struktur ruas gen SRY yang diapit oleh AF126 dan AF127 pada 6 sub populasi kambing lokal Indonesia…………………………………………… 61 15 Polimorfisme nukleotida gen SRY pada 6 sub populasi kambing lokal Indonesia………………………………………………………………… 62 16 Dendogram Neigbour Joining berdasarkan runutan nukleotida gen SRY antara 6 sub populasi kambing lokal Indonesia…………………………... 64 17 Dendogram median-joining network dari 4 haplotip kromosom Y segmen gen SRY dari 6 sub populasi kambing lokal Indonesia : Kacang (K 1 ), Marica (M 1 ), Samosir (S 1 ), Jawarandu (J 1 ), Muara (R 1 ) dan Benggala (B 1 )………………………………………………………............... 65 18 Mutasi nukleotida gen GDF9 ruas promotor pada kelompok induk prolifik (mutan) dan induk non-prolifik (wild) pada Kambing Kacang dan Peranakan Etawah ………………………………………………………….. 70 19 Dendogram kambing lokal Indonesia berdasarkan runutan nukleotida ruas promotor gen GDF9 metode NJ bootstrap 1000x………………….. 72 DAFTAR LAMPIRAN Halaman 1 Pola warna dominan dan warna belang 6 sub populasi kambing lokal………………………………………………………………………. 94 2 Form daftar karakter morfologi yang diukur……………...…..……… 3 Lokasi pengambilan data dan sampel darah kambing lokal (Kambing Kacang, Samosir, Marica, Jawarandu, Muara dan 101 Benggala)……………………………………………………………….. 4 Nama/bangsa kambing, nomor akses dan asal sampel yang digunakan dari GenBank untuk membentuk pohon filogeni ………. 5 Pensejajaran berganda nukleotida ruas D-loop DNA mitokondria pada kambing lokal…………………………….……………………….. 6 Pensejajaran berganda nukleotida gen SRY pada kambing lokal … 7 Pensejajaran berganda nukleotida gen GDF 9 pada kambing lokal Indonesia……………………….………………..………………………. 100 102 103 112 116 PENDAHULUAN Latar Belakang Sumberdaya genetik ternak dihadapkan pada dua tantangan yang saling bertolak belakang yaitu pemanfaatan ternak unggul eksotis untuk memenuhi kebutuhan daging dan susu, sementara disisi lain sumberdaya genetik ternak lokal terus berkurang. Permintaan produksi asal ternak terus meningkat di negara-negara sedang berkembang. FAO memperkirakan kebutuhan daging akan meningkat dua kali lipat pada tahun 2030 dibandingkan dengan kebutuhan pada tahun 2000 dan kebutuhan susu meningkat menjadi lebih dari dua kali lipat. Selain itu, ada kecenderungan penggunaan ternak unggul eksotis untuk mencapai target produksi ternak secara maksimal sehingga perhatian terhadap pengembangan ternak lokal termarjinalkan. Dampak negatif peningkatan ternak unggul eksotis adalah penurunan populasi sumberdaya genetik ternak lokal secara cepat terutama di negara-negara yang sedang berkembang (FAO 2000; Cardellino & Boyazoglu 2009). Sekitar 190 dari 7600 rumpun ternak di dunia yang telah tercatat akan punah selama 15 tahun terakhir, dan 1500 rumpun diantaranya berada dalam status beresiko mendekati kepunahan. Sekitar 60 rumpun terdiri atas sapi, kambing, babi, kuda dan unggas telah punah/hilang (lost) selama 5 tahun terakhir. Tingkat kehilangan rumpun ternak lokal paling tinggi dijumpai di negaranegara sedang berkembang (FAO 2007). Konservasi dan pengembangan rumpun ternak lokal sangat penting, sebab rumpun lokal dapat memanfaatkan pakan berkualitas rendah, lebih tahan terhadap tekanan perubahan iklim dan serangan penyakit, serta sebagai sumber gen-gen yang secara ekonomi menguntungkan untuk peningkatan kesehatan dan performan sifat-sifat produksi pada rumpun ternak unggul komersial (Cardellino 2006; FAO 2007; Ruane 2000). Salah satu komoditas kekayaan plasma nutfah nasional di sub sektor peternakan adalah ternak kambing. Kambing menyebar di berbagai daerah dengan iklim yang berbeda dan terpisah dalam jangka waktu yang lama. Faktor lingkungan dan perlakuan seleksi yang sangat bervariasi mengakibatkan laju perubahan genetik yang sangat beragam (Rout et al. 2008). Di Asia Tenggara khususnya di Indonesia, ternak kambing memegang peranan penting pada petani ternak kecil untuk meningkatkan pendapatan dan 2 juga sebagai sumber daging, pupuk, pengoptimalan tenaga kerja keluarga dan status sosial serta dibutuhkan dalam aspek budaya (Subandriyo 2008). Hampir 99% ternak ruminansia kecil di Indonesia merupakan skala usaha ternak kecil (Soedjana 2008). Kontribusi ternak ruminansia kecil dalam usahatani sangat berperanan penting. Ternak kambing dapat mengkonversi hijauan berkualitas rendah menjadi protein hewani, sebagai sumber pupuk kandang serta sebagai tabungan. Usahatani ternak kambing merupakan bagian dari sistim usahatani secara umum di Indonesia. Sampai saat ini, tampilan morfologi masih umum digunakan secara praktis untuk mengkarakterisasi dan menyeleksi ternak. Penampilan morfologi ini banyak dipengaruhi oleh faktor lingkungan luar seperti ketersediaan pakan dan iklim. Hal ini menjadikan seleksi ternak berdasarkan morfologi membutuhkan waktu lebih lama (Mabrouk et al. 2008; Nsoso et al. 2004; Lanari et al. 2003). Perkembangan ilmu pengetahuan dan teknologi biologi molekuler akhir-akhir ini, khususnya yang berhubungan dengan penggunaan penanda molekuler telah mempercepat karakterisasi sifat-sifat yang bernilai ekonomi tinggi, daya tahan terhadap penyakit, asal-usul dan kekerabatan suatu individu atau rumpun ternak tertentu (Nijman et al. 2003). Penanda molekuler yang populer saat ini, antara lain DNA-mitokondria dan DNA mikrosatelit. Penanda DNA-mitokondria menggambarkan karakteristik yang diturunkan melalui garis induk (maternal) (Fan-Bin 2007). Penanda DNA mikrosatelit menggambarkan karakter rekombinasi (Wimmers et al. 2000; Tadelle 2003; Zhang et al. 2002). Selain itu penanda molekuler DNA kromosom Y menggambarkan karakteristik yang diwariskan melalui pejantan (paternal) (Verkaar et al. 2003). Sejalan dengan perkembangan yang pesat lalu lintas ternak antar kawasan maka lama kelamaan terjadilah proses adaptasi terhadap agroekosistem yang spesifik sesuai dengan lingkungan dan manajemen pemeliharaan yang ada ditempat tertentu. Dengan demikian terjadi evolusi yang membuka kemungkinan munculnya rumpun/galur/ras kambing yang baru. Balai Penelitian Ternak Ciawi sejak tahun 1995 sudah memulai mengkarakterisasi Kambing Kacang, Peranakan Etawah, Kosta dan Gembrong pada tahun 1997 (Setiadi et al. 1995; 1997). Diperkirakan masih banyak lagi rumpun kambing lokal Indonesia lainnya yang belum dikarakterisasi dan sebagian mungkin sudah hampir langka atau jumlah populasinya sudah mendekati punah. 3 Perwujudan tujuan pelestarian plasma nutfah dan spesifikasi ternak menurut bangsa dapat dimulai dengan sebutan khusus menurut wilayah, misalnya Kambing Marica yang ada di Sulawesi Selatan, Kambing Benggala di Nusa Tenggara, Kambing Lakor di Maluku, Kambing Samosir dan Kambing Muara di Sumatera Utara, Kambing Jawarandu di Jawa Tengah dan jenis kambing lainnya di daerah Indonesia. Penelitian tentang karakterisasi kambing lokal Indonesia perlu dilakukan untuk mendapatkan data dan informasi potensi sumberdaya genetik kambing lokal Indonesia. Tujuan Penelitian 1. Menginventarisasi karakteristik fenotip secara kualitatif dan kuantitatif ciri-ciri morfologi pada kambing lokal Indonesia. 2. Mengkarakterisasi keragaman genetik daerah D-loop DNA Mitokondria pada kambing lokal Indonesia untuk mengetahui asal usul menurut garis keturunan maternal. 3. Mengkarakterisasi keragaman gen SRY ruas DNA kromosom Y pada kambing lokal Indonesia untuk mengetahui asal usul menurut garis keturunan paternal. 4. Mengidentifikasi keragaman gen GDF9 yang berhubungan dengan sifat prolifik pada kambing lokal Indonesia. Manfaat Penelitian Hasil penelitian ini diharapkan dapat memberikan sumbangan pemikiran yang bermanfaat untuk 1. Mengembangkan kambing lokal Indonesia dengan memperhatikan keragaman populasi yang ada. 2. Menyediakan informasi yang dapat digunakan Pemerintah Daerah khususnya Dinas Peternakan sebagai pedoman dalam menyusun perencanaan pembangunan peternakan, khususnya pengembangan kambing lokal Indonesia dalam rangka pelestarian plasma nutfah kambing lokal Indonesia yang bertujuan berkelanjutan. untuk pengembangan dan pemanfaatannya secara 4 3. Menyediakan informasi tentang pemanfaatan ciri-ciri fisik kambing lokal Indonesia yang dapat digunakan sebagai sumber keragaman genetik untuk pembentukan galur bibit kambing unggul. Kerangka Pemikiran Keragaman sumberdaya genetik meningkatkan produktivitas kambing. merupakan modal dasar untuk Data dan informasi tentang karakter fenotipik dan genetik ternak kambing lokal Indonesia sampai saat ini masih sangat terbatas. Penelitian ini perlu dilakukan untuk mendapatkan data fenotifik dan genotip beberapa kambing lokal Indonesia. Penelitian dirancang dengan metode survei dan laboratorium. Metode survei menggunakan purposive sampling, yaitu dengan memilih lokasi yang diduga masih bisa ditemukan populasi kambing lokal terutama daerah yang paling padat populasinya. Lokasi dipilih berdasarkan informasi dari instansi terkait di Propinsi kemudian menuju Kabupaten dan selanjutnya menuju Kecamatan dan Desa. Sampel kambing diusahakan diambil dari lokasi yang berjauhan. Pada lokasi terpilih dilakukan pengambilan sampel darah, penimbangan, pengamatan warna dan pola warna kambing serta ukuran-ukuran tubuh diukur dengan pita, jangka dan tongkat ukur. Kegiatan laboratorium terdiri atas ekstraksi DNA dari darah mengikuti metode Sambrook yang dimodifikasi, menganalisis DNA mitokondria, DNA kromosom Y dan gen GDF9. Diagram alir kerangka pemikiran penelitian Studi keragaman fenotipik dan genetik beberapa sub populasi kambing lokal Indonesia dan pemanfaatannya secara berkelanjutan dapat dilihat pada Gambar 1. strategi 5 KAMBING LOKAL INDONESIA POTENSI • Sebagai sumberdaya genetik ternak daerah/ nasional • Kambing penghasil daging, susu, pupuk • Berkontribusi signifikan terhadap pendapatan peternak dan status sosial PERMASALAHAN • Data dan informasi karakter morfometrik kambing lokal masih terbatas • Data dan informasi pemetaan genetik tentang pengelompokan dan asal usul secara maternal dan paternal masih terbatas • Data dan informasi potensi keragaman gen fungsional yang berhubungan dengan sifat prolifik pada kambing lokal masih terbatas. ANALISIS MORFOMETRIK Data kualitatif (warna dan pola warna tubuh) dan kuantitatif (ukuran tubuh) ANALISIS DNA MITKONDRIA Keragaman genetik DNA mitokondria untuk mengiden tifikasi hubungan asal usul kambing lokal secara maternal ANALISIS DNA KROMOSOM Y Keragaman genetik DNA kromosom Y untuk mengiden tifikasi hubungan asalusul secara paternal ANALISIS KERAGAMAN GEN GDF9 Identifikasi keragaman gen GDF9 pada kambing lokal Status pemetaan keragaman fenotipik dan identitas genetik enam sub populasi kambing lokal Indonesia Data karakteristik sumberdaya genetik dapat digunakan sebagai salah satu bahan pertimbangan dalam menetapkan dan menentukan arah strategi pemanfaatan sumberdaya genetik kambing lokal secara berkelanjutan Gambar 1 Diagram alir kerangka pemikiran penelitian TINJAUAN PUSTAKA Sejarah Budidaya Ternak Kambing Ternak kambing (Capra hircus) sering diartikan sebagai ternak yang dapat membantu memecahkan masalah kemiskinan di kalangan peternak, karena kemampuannya dalam memanfaatkan hijauan dalam jumlah terbatas seperti pada lingkungan yang kritis dan kering/lahan marjinal (MacHugh & Bradley 2001). Kambing merupakan hewan pertama yang didomestikasi, diduga berasal dari Kambing liar Capra aegargus. Pada awalnya sekitar 10 000-11 000 tahun yang silam di daerah Kawasan Timur Tengah manusia zaman Neolithic mulai memelihara kambing dalam jumlah kecil untuk mendapatkan susu, daging dan kotorannya sebagai bahan bakar, juga sebagai bahan untuk pakaian dan bangunan yang terbuat dari bulu, tulang, kulit dan urat daging (MacHugh et al. 2001; Zeder et al. 2000). Saat ini lebih dari 300 rumpun ternak kambing yang hidup di berbagai iklim dan ketinggian, mulai dari dataran tinggi sampai ke daerah dataran rendah. Ahli arkeologi melaporkan dua tempat yang berbeda sebagai asal dari pertama kali proses domestikasi kambing dilakukan, yaitu; Lembah Sungai Eupharate di Nevali Cori, Turki (11 000 B.C.) dan di Pegunungan Zagros di Garj Dareh, Iran (10 000 B.C.). Kemungkinan situs yang lain adalah Indus Basin, di daerah Mehgarh, Pakistan (9 000 B.C.) dan kemungkinan di Pusat Anatolia dan bagian utara Levant. Situs arkeologi yang lain yang penting menunjukkan adanya proses domestikasi kambing di Cayonu, Turki (8 500 - 8 000 B.C.), Tell Abu Hureyra, Syria (8 000 – 7 400 B.C.), Jerico, Israel (7 500 B.C.) dan Ain Ghazal, Jordan (7 600 – 7 500 B.C.) (Hirst 2008). Kambing yang dipelihara (Capra aegagrus hircus) berasal dari 3 kelompok kambing liar yang telah dijinakkan, yaitu Bezoar goat atau kambing liar Eropa (Capra aegagrus), kambing liar India (Capra aegagrus blithy), dan Makhor goat atau Kambing Makhor di pegunungan Himalaya (Capra falconeri). Sebagian besar kambing yang diternakkan di Asia berasal dari keturunan Bezoar, termasuk Kambing Gunung Sumatra (Caprinae sumatraensis) atau disebut juga Kambing Gurun (Maddox & Cockett 2007). Kambing biasanya dibedakan berdasarkan letak geografis, karakteristik morfologi, dan performan produksi. Kambing berdasarkan ukuran tubuh (karakteristik morfologi) dibedakan atas tiga tipe yaitu; kambing tipe besar, tipe 7 sedang dan tipe kecil. Berdasarkan performan produksi kambing dibedakan atas kambing tipe perah, tipe pedaging dan tipe dwi guna (dual purpose). Saat ini usaha ternak kambing juga sangat berperan mendukung kebutuhan akan ternak Qurban bagi yang beragama Islam yang merupakan agama mayoritas di Indonesia dan juga pada aliran kepercayaan seperti agama Parmalim di Pulau Samosir dan daerah sekitar Danau Toba. Saat ini, usaha ternak secara komersial sudah berkembang di beberapa daerah di Indonesia untuk memproduksi susu kambing, dimana kualitas susu kambing mempunyai beberapa keunggulan jika dibandingkan dengan jenis susu ternak lainnya. Usaha ternak kambing perah berperan sekaligus menghasilkan ternak kambing potong. Keragaman Genetik Ternak Keragaman genetik terjadi tidak hanya antar rumpun tetapi juga di dalam satu rumpun yang sama, antar populasi maupun di dalam populasi. Pada spesies ternak domestik suatu identifikasi tingkat keragaman, terutama pada lokus-lokus yang mempunyai sifat bernilai penting mempunyai keterkaitan dengan seleksi dalam program pemuliaan (Handiwirawan & Subandriyo 2004; Abdullah 2008). Salah satu alat bantu yang dapat digunakan untuk mendeteksi keragaman populasi adalah DNA mitokondria dan DNA mikro satelit (Muladno 2006; Yuwono 2006) dan DNA kromosom Y segmen gen SRY. Keragaman genetik dalam populasi merupakan modal dasar aplikasi teknologi pemuliaan dalam pemanfaatan hewan. Keragaman genetik populasi yang digambarkan dalam keragaman penampilan hewan adalah refleksi informasi genetik yang dimilikinya. Perbedaan penampilan disebabkan selama proses domestikasi tipe atau rumpun-rumpun hewan terpisah secara genetik karena adanya proses adaptasi dengan masing-masing lingkungan lokal dan kebutuhan komunitas lokal sehingga dihasilkan rumpun yang berbeda (Muladno 2006). Adanya kemampuan adaptasi hewan disebabkan hewan memiliki kemampuan menghasilkan lebih dari satu alternatif bentuk morfologi, status fisiologi, dan atau tingkah laku sebagai reaksi terhadap lingkungan (Noor 2008). Lebih dari 12 000 tahun yang lalu terdapat 12 spesies ternak telah didomestikasikan dan berevolusi sehingga menjadi rumpun (breed) yang secara genetik unik dan berbeda, beradaptasi terhadap lingkungan dan komunitas setempat. Saat ini terdapat sekitar 6 000 – 7 000 rumpun ternak domestik dari 8 spesies yang telah terdomestikasi, bersama dengan lebih dari 80 spesies kerabat liarnya yang merupakan sumberdaya genetik ternak di bumi ini yang berperan penting untuk pangan dan produksi pertanian. Berbagai rumpun ternak yang telah berkembang dalam berbagai sistem dan lingkungan yang ada saat ini telah menghasilkan berbagai kombinasi gen yang unik. Gen-gen ini tidak hanya menentukan kualitas sifat produksi dari masing-masing rumpun, tetapi juga terhadap kemampuan adaptasinya pada perubahan kondisi lingkungan lokal termasuk makanan, ketersediaan air, iklim dan hama penyakit (FAO 2001). Berbagai macam kebutuhan manusia sehari-hari dipenuhi dari spesies ternak, dalam bentuk pangan maupun kebutuhan lainnya. Namun hanya sebagian kecil dari total keragaman genetik ternak dan kerabat liarnya, yakni sekitar 40 spesies yang memenuhi sebagian besar proporsi dari produksi ternak global. Keragaman ternak di dalam genetik ternak dan beberapa kerabat lainnya telah menjadi sumber keragaman dari rumpun dan populasi ternak. Keragaman genetik ini penting dalam pembentukan ternak modern dan akan terus berkelanjutan di masa mendatang (Subandriyo & Setiadi 2003). Sumberdaya genetik ternak sedikitnya memiliki empat manfaat, yaitu (1) keberlanjutan dan peningkatan produksi pangan; (2) memaksimumkan produktivitas lahan dan sumberdaya pertanian; (3) pencapaian pertanian berkelanjutan untuk memberikan keuntungan masa kini dan generasi rumpun ternak yang akan datang; (4) pemenuhan keanekaragaman baik yang telah maupun yang belum diketahui manfaatnya bagi kehidupan sosial masyarakat. Ketersediaan keanekaragaman genetik ternak, termasuk kambing akan mempengaruhi keberhasilan strategi pemuliaan untuk masa yang akan datang (FAO 2007). Pelestarian Sumberdaya Genetik Ternak Keragaman genetik ini penting dalam pembentukan rumpun dan populasi ternak modern dan akan terus berlanjut untuk masa mendatang. Punahnya keragaman plasma nutfah ternak tidak akan dapat diganti meskipun dengan kemajuan bioteknologi, paling tidak sampai saat ini. Negara-negara sedang berkembang pada umumnya berada pada iklim dengan perubahan temperatur yang ekstrim antara musim panas dan hujan. Pada kondisi seperti ini akan terbentuk rumpun ternak yang beradaptasi. Walaupun produktivitasnya rendah apabila dibandingkan dengan dengan rumpun yang terdapat di daerah temperate (eksotik), rumpun ternak ini memiliki daya tahan terhadap berbagai macam 9 penyakit; tahan terhadap fluktuasi ketersediaan dan mutu pakan dan air; tahan terhadap perubahan temperatur, kelembaban dan pengaruh iklim ekstrim lainnya. Rumpun ternak ini juga beradaptasi terhadap pemeliharaan yang kurang baik sehingga memiliki nilai yang sangat berharga untuk mengantisipasi berbagai perubahan alam dan lingkungan diwaktu yang akan datang (FAO 2007). Dengan demikian, pelestarian terhadap sumberdaya genetik ternak lokal sebagai bagian dari komponen keanekaragaman hayati adalah penting untuk memenuhi kebutuhan pangan, pertanian dan perkembangan sosial masyarakat di masa yang akan datang. Ada beberapa alasan untuk ini, antara lain: (1) lebih dari 60 persen dari rumpun-rumpun hewan ternak di dunia berada di negaranegara sedang berkembang, (2) konservasi rumpun ternak lokal tidak menarik bagi petani, (3) secara umum tidak ada program monitoring yang sistematis dan tidak tersedianya informasi deskriptif dasar sebagian besar sumberdaya genetik hewan ternak, serta (4) sedikit sekali rumpun-rumpun hewan ternak asli yang telah digunakan dan dikembangkan secara aktif (FAO 2001; 2007). Pelestarian sumberdaya genetik ternak pada dasarnya dapat dilakukan salah satu atau gabungan dari: (1) mempertahankan populasi ternak hidup baik dalam bentuk in-situ maupun ex-situ pada satu tempat tertentu, (2) penyimpanan beku (cryogenic), dan (3) penyimpanan dalam bentuk DNA. Dalam beberapa hal, mempertahankan populasi merupakan metode yang lebih praktis. Pelestarian pada ternak hidup mempunyai beberapa keuntungan antara lain; rumpun-rumpun ternak yang dilestarikan secara bertahap dapat merespon terhadap perubahan pengaruh eksternal dan memungkinkan dilakukan evaluasi kinerjanya (FAO 2007). Sumberdaya Genetik Kambing Indonesia Sumberdaya ternak kambing di Indonesia saat ini terdiri dari tiga kelompok, yakni: (1) ternak asli, (2) ternak impor, dan (3) ternak yang telah beradaptasi dalam jangka waktu lama sehingga membentuk karakteristik tersendiri (ternak lokal). Pentingnya nilai konservasi pada kelompok hewan ternak ini, beberapa rumpun ternak ini perlu dijadikan target konservasi sekaligus pemanfaatannya (Utoyo 2002). Rumpun ternak kambing di Indonesia dan rumpun kambing lainnya adalah merupakan hasil domestikasi sekitar 10000 tahun yang lampau. Kambing eksotis masuk ke Indonesia melalui daratan India 10 terus melalui Khyber Pass, kemudian menyebar melalui pulau Sumatra, Kalimantan, Sulawesi, Jawa sampai Indonesia bagian Timur. Kambing Kacang merupakan rumpun kambing asli Indonesia, bentuk badannya kecil dengan tinggi pundak sekitar 50-60 cm serta prolifik. Introduksi rumpun kambing impor Benggala dari India dimulai oleh orang-orang Arab dan kambing-kambing tersebut didatangkan melalui pelabuhan pantai utara Pulau Jawa. Mulai pada tahun 1911-1931 didatangkan rumpun-rumpun Kambing Kashmir, Angora (Montgomey), Benggala dan Etawah untuk stasiun ternak kambing atau stasiun peternakan di Keresidenan Kedu, Solo, Yogyakarta, Banyumas, Pekalongan, Pangalengan, Padang Mangatas, Wlingi (Blitar), Sumba, dan Sumbawa. Disamping dari India pada tahun 1928 pernah pula diimpor dari Negeri Belanda yaitu “Hollandse Edelgeiten” (Kambing Belanda Murni). Rumpun kambing dari India selanjutnya disilangkan dengan rumpun kambing lokal Indonesia dengan cara digaduhkan atau menempatkan pejantan Etawah murni atau persilangan dengan proporsi darah Etawah yang cukup tinggi di desa-desa yang akan dikembangkan peternakan kambingnya. Hasil persilangan tersebut dikenal dengan nama Peranakan Etawah, yang proporsi darah Etawahnya sangat beragam. Selain itu juga terdapat rumpun kambing lain yang berkembang di daerah tertentu yang merupakan kambing lokal tradisional diantaranya Kambing Gembrong (di Bali), Kambing Kosta (di Banten), Kambing Bligon, Kambing Jawarandu (di Jawa Tengah dan Daerah Istimewa Yogyakarta) dan beberapa jenis kambing lokal lainnya yang belum diidentifikasi secara ilmiah. Setelah zaman kemerdekaan diimpor atau diintroduksikan beberapa rumpun kambing, baik dalam bentuk hidup atau mani beku. Rumpun kambing yang pernah dintroduksikan antara lain Kambing Saanen dan Kambing Anglo Nubian. Bahkan akhir-akhir ini telah diintroduksikan pula Kambing Boer dari Australia yang dipersilangkan dengan Kambing Kacang atau Peranakan Etawah dalam bentuk pejantan hidup atau mani beku (Subandriyo 2004). Terjadinya persilangan antara kambing impor dengan kambing asli Indonesia (Kacang) serta adanya aklitimasi dan isolasi selama puluhan bahkan ratusan tahun di suatu lokasi tertentu dapat menyebabkan terbentuknya kelompok kambing lokal atau sub populasi dengan komposisi genetik yang unik pula. Terbentuknya galur/kelompok kambing bisa juga disebabkan terisolasinya suatu lokasi, sehingga menyebabkan terjadinya perbedaan genetik akibat 11 adanya penghanyutan genetik (random genetic drift) seperti dilaporkan Freeland (2005). Kambing Muara dijumpai di daerah Kecamatan Muara, Kabupaten Tapanuli Utara. Berdasarkan pada penampilannya kambing ini nampak gagah, tubuh kompak dan sebaran warna bervariasi antar warna bulu coklat kemerahan, putih dan ada juga yang berwarna bulu hitam. Bobot Kambing Muara ini lebih besar dari pada Kambing Kacang dan diduga mempunyai potensi sebagai ternak prolifik. Kambing Benggala menurut cerita dari peternak diduga merupakan hasil persilangan Kambing Black Bengal dengan kambing lokal yang diduga dibawa pendatang/pedagang dari India, Bangladesh dan Arab ke daerah sekitar Pulau Timor dan Pulau Flores di Propinsi Nusa Tenggara Timur sebelum zaman penjajahan Hindia Belanda. Selang waktu yang sudah ratusan tahun persilangan kambing tersebut mengalami proses adaptasi dengan lingkungan setempat (Batubara et al. 2007). Kambing Benggala secara umum lebih besar dari Kambing Kacang, umumnya didominasi warna hitam dan sedikit berwarna kecoklatan. Menurut FAO (2000) rumpun adalah bagian kelompok tertentu (subspecific group) dari ternak domestik dengan karakteristik eksternal yang dikenal dengan penilaian visual atau kelompok yang dipisahkan oleh geografi dan budaya secara fenotipik. Rumpun berkembang menurut perbedaan geografi dan budaya untuk memenuhi kebutuhan yang serupa dan telah diterima sebagai identitas yang terpisah. Berdasarkan adaptasi terhadap kondisi lokal rumpun dibedakan atas rumpun lokal dan rumpun introduksi. Rumpun lokal dapat dibedakan lagi atas rumpun asli (indigenous breed, native breed) adalah ternak yang berdasarkan sejarah terbukti berasal dari negara tersebut dan rumpun tradisional (rumpun lokal) adalah ternak yang sejarahnya tidak terbukti berasal dari negara tersebut tetapi selama 30-50 tahun telah diternakkan di negara tersebut, terbukti mempunyai catatan silsilah selama lima generasi. Rumpun introduksi (rumpun asing, exotic, alocthonous) yang tidak berasal dari suatu negara atau tidak secara kontinu diternakkan di suatu negara lebih dari 50 tahun (Sapi, kuda) dan 30 tahun untuk ternak lainnya (FAO 2007). Penetapan dan pengakuan rumpun/galur ternak di Indonesia dilakukan oleh Pemerintah melalui Kementerian Pertanian. Pemerintah menyusun tatacara mengenai pengujian, penilaian, penetapan dan pengakuan, pemberian nama dan 12 pelepasan rumpun/galur ternak. Istilah “penetapan” adalah sebagai bentuk pengakuan dari negara terhadap rumpun ternak yang telah ada dan dibudidayakan secara turun temurun oleh peternak dan menjadi milik masyarakat (rumpun lokal atau rumpun asli). Istilah “pengakuan” adalah suatu bentuk pengakuan negara terhadap rumpun dan/atau galur ternak hasil pemuliaan/ introduksi/rekayasa genetik (Puslitbangnak 2007). Sifat Kuantitatif dan Kualitatif Penampilan individu yang nampak dari luar disebut sebagai fenotipik, yang dapat dibedakan menjadi sifat kuantitatif dan kualitatif (Hardjosubroto 2001). Mabrouk et al. (2008) mengemukakan bahwa karakter kuantitatif adalah ciri-ciri dari mahluk hidup yang dapat diukur, dihitung atau diskor, misalnya ukuran-ukuran tubuh. Karakter ini ditentukan oleh banyak pasang gen (poligenik) dan sangat dipengaruhi oleh lingkungan. Beberapa sifat kuantitatif pada ternak mempunyai hubungan satu sama lain, hubungan ini secara statistik disebut dengan korelasi. berkorelasi menjadi penting karena seleksi terhadap Sifat-sifat yang satu sifat akan menyebabkan kemajuan atau kemunduran bagi sifat lain yang berkorelasi dengan sifat tersebut (Nsoso et al. 2004). Berlawanan dengan karakter kuantitatif, karakter kualitatif adalah karakter yang pada umumnya dijelaskan dengan kata-kata atau gambar. Sifat ini sedikit sekali atau bahkan tidak ada hubungannya dengan kemampuan produksi, namun sifat ini mungkin penting sebagai penciri bagi rumpun atau tipe ternak tertentu, misalnya warna dan pola warna tubuh. Sifat ini diatur oleh satu atau beberapa pasang gen saja, dan sedikit sekali dipengaruhi oleh lingkungan (Noor 2008). Sponenberg (2004) mengemukakan bahwa tipe dasar tatawarna bulu dapat dibedakan menurut: pertama, warna yang meliputi seluruh permukaan tubuh sehingga membentuk warna seragam atau homogen atau warna tunggal; dan kedua, heterogen atau campuran. Warna heterogen ini memiliki dua tipe yang berbeda, yaitu: (a) komposit, apabila pada tubuh ditemukan bidang-bidang warna yang berbeda (spotted); dan (b) campuran, apabila bulu-bulu dari warna yang berbeda tampak secara bergantian satu dengan lainnya. Warna pada kambing umumnya diklasifikasikan kedalam warna tunggal (unicoloured) dan terpola (patterned). 13 Penanda Genetik Penanda adalah karakter yang dapat diwariskan dan berasosiasi dengan genotip tertentu dan digunakan untuk mengkarakterisasi genotip. Potensi penggunaan penanda sebagai alat untuk melakukan karakterisasi genetik telah dikenal sejak puluhan tahun yang lalu. Penanda ini dikategorikan atas penanda morfologi, sitologi, dan yang terbaru adalah penanda molekuler (Simianer 2006). a. Penanda Morfologi Penanda morfologi (fenotipik) merupakan penanda yang telah banyak digunakan baik dalam program genetika dasar maupun dalam program praktis pemuliaan, karena penanda ini paling mudah untuk diamati dan dibedakan. Pengukuran parameter tubuh biasa digunakan untuk menduga asal usul rumpun ternak. Ukuran-ukuran tubuh sangat berguna untuk menentukan asalusul dan hubungan filogenetik antar spesies, rumpun dan tipe ternak yang berbeda. Beberapa penelitian telah dilakukan menggunakan ukuran-ukuran tubuh untuk membedakan kelompok Kambing Tswana (Nsoso et al. 2004). Mabrouk et al. (2008) juga telah melakukan penelitian menggunakan beberapa ukuran tubuh (bobot badan, panjang badan, tinggi pundak, dalam dada, lingkar dada, tinggi pinggul, lingkar pinggul, dalam pinggul, panjang ekor, lebar ekor dan tebal ekor) sebagai peubah pembeda kelompok pada beberapa kelompok kambing lokal di Tunisia. Dossa et al. (2007) mengemukakan bahwa kesamaan fenotipik dapat menunjukkan identitas genetik, walau terdapat beberapa batasan, antara lain: fenotipik yang identik dapat disebabkan olel alel-alel yang berbeda atau oleh gen-gen pada lokus yang berbeda. Dalam hal tertentu, mungkin terdapat perbedaan dalam daya ekspresi (derajat manifestasi pada satu individu) atau oleh gen dominan (frekuensi satu sifat diekspresikan relatif terhadap sejumlah pembawa gen tertentu yang diketahui dalam satu populasi). Kemiripan fenotipik dapat juga disebabkan oleh fenokopi, yakni kemiripan satu fenotip yang diakibatkan satu genotip tertentu oleh aksi lingkungan pada genotip lainnya. Namun demikian, penanda ini memiliki kelemahan karena dipengaruhi oleh lingkungan, memperlihatkan sifat menurun dominan/resesif dan banyak yang hanya dapat diamati pada tingkat umur tertentu. 14 b. Penanda Molekuler Menurut Cardellino dan Boyazoglu (2009) aplikasi penanda molekuler yang paling penting adalah untuk pembuatan peta genetik, yang dapat digunakan untuk memeriksa lokasi suatu gen yang bertanggung jawab terhadap suatu sifat yang sederhana, misalnya resistensi terhadap penyakit atau sifat kuantitatif yang komplek pada kromosom. Penanda molekuler ini ada pada tingkat DNA, maka penanda ini bebas dari pengaruh-pengaruh epistasis, lingkungan dan fenotip sehingga dapat menyediakan informasi genetik yang defenitif untuk digunakan dalam mempelajari keragaman genetik, mendeteksi gen-gen major dan mempelajari sifat-sifat genetik yang komplek. Tehnik ini sangat membantu pemulia dalam melakukan studi genetik dengan ketepatan yang tinggi. Untuk mendapatkan informasi genetik dapat dilakukan dengan menggunakan penanda molekuler, seperti isozim, RFLP (restriction fragment length polymorphism), RAPD (random amplified polimorphic DNA), AFLP (amplified fragment length polymorphism) dan lain-lainnya. Penanda molekuler terbaru yang relatif mudah diamati adalah DNA mikrosatelit. DNA Mitokondria Organisme eukariot termasuk ternak domestik, sumber DNA dapat diperoleh oleh organel-organel sitoplasmik antara lain DNA mitokondria. DNA mitokondria memiliki karakteristik sebagai molekul DNA yang diturunkan secara utuh tanpa adanya rekombinasi, memiliki molekul dengan ukuran kecil/pendek yang susunannya berbeda dengan DNA inti (Lewin 2000) dan memiliki variasi basa nukleotida yang lebih tinggi dibandingkan dengan DNA inti. Tingginya variasi basa nukleotida disebabkan DNA mitokondria memiliki laju perubahan 510 kali lebih tinggi dibandingkan DNA inti (Muladno 2006; Fan-Bin 2007). DNA mitokondria terutama daerah D-loop, sangat baik digunakan untuk analisis keragaman hewan, baik di dalam spesies maupun antar spesies (Muladno 2006). Setiap sel mengandung satu hingga ratusan DNA mitokondria. DNA mitokondria merupakan DNA utas ganda yang berbentuk sirkuler (Freeland 2005), mengandung sejumlah gen penting untuk respirasi dan pembentukan energi sel tubuh dan fungsi lainnya, sehingga relatif lebih mudah untuk mengisolasi nukleotidanya dari genom (MacHugh & Bradley 2001). Genom mitokondria hewan berukuran relatif kecil dan terdapat dalam jumlah banyak, maka eksplorasi rumpun dan penelaahannya lebih mudah. 15 DNA mitokondria (mtDNA) mempunyai beberapa kelebihan yang menjadikannya banyak digunakan untuk mengidentifikasi keanekaragaman genetik dan dinamika populasi. Beberapa kelebihan tersebut adalah (1) memiliki ukuran yang kompak dan relatif kecil (16 000 – 20 000 pasang basa), tidak sekomplek DNA inti sehingga dapat dipelajari sebagai satu kesatuan yang utuh; (2) berevolusi lebih cepat dibandingkan dengan jelas perbedaan antara populasi dan hubungan kekerabatannya; (3) hanya sel telur yang menyumbangkan material mitokondria sehingga mitokondria DNA hanya diturunkan dari induk betina; dan (4) bagian-bagian dari genom mitokondria berevolusi dengan laju yang berbeda, sehingga dapat berguna untuk studi sistematika dan penelusuran kesamaan asal-usul. DNA mitokondria telah banyak digunakan sebagai penanda molekul untuk studi genetika populasi, penelusuran asal-usul dan pelacakan beberapa penyakit degeneratif, penuaan serta kanker (Wandia 2001). DNA mitokondria telah dikarakterisasi dengan lebih baik pada sebagian besar ternak dan telah digunakan untuk studi evolusi (Freeland 2005). Tingkat evolusi dari suatu gen atau bagian DNA yang berbeda merupakan faktor penting yang menentukan penggunaan penanda DNA dalam studi sistematika dan biogeografi. Umumnya, gen-gen yang terkonservasi dengan baik (berevolusi lambat) dapat dijadikan dasar penelusuran asal-usul atau filogeni. Sebaliknya, gen-gen yang tidak terkonservasi dengan baik (berevolusi cepat) dapat digunakan untuk perbandingan galur-galur baru (Chen et al. 2005). DNA mitokondria hewan secara umum memiliki jumlah dan jenis gen yang sama yaitu 13 daerah yang mengkode protein (URF1, URF2, URF3, URF4, URF5, URF6, URF6L, URF4L, Cytochrome Oxidase unit I, Cytochrome Oxidase unit II, Cytochrome Oxidase unit III, Cytochrome –b dan ATPase 6); 2 gen pengkode rRNA yaitu 12S rRNA dan 16S rRNA; 22 gen pengkode tRNA (Freeland 2005). Perkembangan sekarang ini ke-8 URF adalah diidentifikasi menjadi gen-gen 7 sub unit NADH-dehidrogenase (ND 1-6 dan ND 4L) dan sisa ATPase 8 (Lewin 2000). Daerah bukan pengkode, hanya terdiri atas daerah kontrol (control region) yang memegang peranan penting dalam proses transkripsi dan replikasi genom mitokondria. Pada mamalia, daerah bukan pengkode meliputi daerah bukan pengkode utama yang merupakan tempat awal replikasi H strand (OH). Daerah bukan penyandi utama terletak pada wilayah dislacement–loop (D-loop region). Bagian lainnya adalah daerah bukan 16 pengkode segmen minor yaitu tempat awal replikasi L strand (OL) yang terletak pada gugus gen tRNA antara gen CO I dan ND 2. Gambar 2 Skema genom daerah D-loop kambing (Sumber : Freeland 2005) DNA Kromosom Y Penentuan jenis kelamin laki-laki pada manusia tergantung dari aktivitas yang disebut testis determining factor yang terdiri dari exon tunggal dan dikodekan oleh gen SRY. Karakteristik dari daerah kotak SRY-HMG merupakan target ideal untuk pengembangan dari uji penentuan jenis kelamin berdasarkan DNA (Prashant et al. 2008). Mutasi pada gen SRY berperan menentukan XY Gonadal Dysgenesis (XYGD). Fenotip XYGD yang telah dilaporkan antara lain pada manusia (Cohen dan Shaw 1965), kuda (Power 1986), sapi (Kawakura et al. 1996) dan kerbau rawa (Iannuzzi et al. 2001) 17 Sampai saat ini hanya sekitar 15-20% kasus yang ditemukan bermutasi (McElreavey 1996) sementara sebagian besar lainnya masih belum diketahui faktor apa yang mempengaruhinya (Veiteia et al. 2001). Gen SRY bersifat nonrekombinan pada bagian Y kromosom, sehingga sekuen gen ini dapat digunakan untuk menganalisis dan menyelidiki proses evolusi dan asal usul dari individu secara paternal atau menurut garis keturunan pejantan (Parma et al. 2004; Prashant et al. 2009). Gen SRY yang terletak di komosom Y bertanggung jawab untuk menentukan jenis kelamin pada mammalia (Sinclair et al. 1990) dan mengkodekan protein sebanyak 204 asam amino. Gen SRY terpusat di daerah High Mobility Groups (HMG) yang mempunyai variasi tinggi sehingga ideal digunakan untuk menguji garis keturunan paternal berdasarkan DNA (Prashant et al. 2008). Penelitian tentang Kromosom Y pada kambing telah dilakukan pada Kambing Sardinian dan Maltese (Sechi et al. 2009), Kambing Sangamneri (Prashant et al. 2009). Skema genom gen SRY kromosom Y dan kromosom X dapat dilihat pada Gambar 3. Gen SRY Sentromer Gambar 3 Skema genom gen SRY kromosom Y dan kromosom X (Sumber: Seli & Sakkas 2005). Gen yang Berhubungan dengan Sifat Prolifik Sifat prolifik pada ternak ruminansia bisa dibedakan menjadi 2 jenis yaitu: Superovulasi yang bersifat multizigous, yaitu jumlah anak sekelahiran bisa lebih 18 dari 2 bahkan bisa sampai beranak lima ekor dalam satu periode kelahiran, biasanya dijumpai pada ternak ruminansa kecil seperti domba dan kambing, dan yang bersifat monozigous, yaitu pada ternak ini ada kemungkinan satu sel telur berkembang dengan cara membelah menjadi dua, biasanya ini dijumpai pada ternak ruminansia besar. Folikel merupakan titik awal yang perlu disoroti sebagai salah satu ”pabrik” penghasil sel telur dan penghasil hormon conseptus (kebuntingan) maupun mammogenic (kelenjar susu), bahkan secara genetik dapat digunakan sebagai alat seleksi untuk memperoleh ternak unggul berdasarkan jumlah anak yang dilahirkan. Selama ini dibidang fisiologi reproduksi telah terjadi inefisiensi pemanfaatan folikel pada induk betina. Kedua ovarium betina mengandung 40 000 – 300 000 folikel bahkan lebih tergantung pada jenis ternak. Pada ruminasia (sapi,domba,kambing) berkisar 50 000 – 150 000 folikel (Hafez 1993), namun selama hidupnya ternyata hanya beberapa folikel yang berovulasi, sehingga ratusan ribu sisanya tidak termanfaatkan. Folikel baik sebelum maupun sesudah ovulasi menjadi corpus luteum merupakan organ transitor yang memiliki peran regulator di dalam proses reproduksi, terutama sebagai sumber penghasil sel telur maupun hormon conseptus endogen (Gemmell 1995). Kematian anak dari tipe kelahiran 3 atau lebih sangat tinggi (Inounu et al. 1993), sehingga potensi keuntungan dari betina dengan jumlah anak banyak ini tidak terlihat. Betina-betina yang merawat anak banyak membutuhkan tingkat nutrisi yang sangat baik pada fase sebelum dan sesudah kelahiran dan juga memerlukan perawatan serta perhatian yang lebih banyak, jika persentase anak yang hidup ingin dicapai. Tehnik laparoskopi dapat dilakukan pada ternak kambing betina untuk mengamati produksi sel telur (laju ovulasi) dengan cara menghitung jumlah corpus luteum (CL) yang dihasilkan. Batas atas keragaan reproduksi adalah jumlah sel telur yang dihasilkan oleh seekor induk per satu siklus birahi, yang diamati dengan cara menghitung jumlah corpus luteum (badan kuning) dari kedua indung telur pada hari ke 3-10 setelah birahi. Faktor lingkungan merupakan faktor pendukung apakah batas atas tersebut dapat dicapai. Laju ovulasi adalah rataan jumlah sel telur yang dihasilkan oleh seekor induk setiap siklus birahi. Di dunia ini ada beberapa rumpun kambing yang sangat prolifik, ditandai dengan laju ovulasi dan jumlah anak sekelahiran tinggi yang bisa melahirkan anak 3-5 ekor anak per kelahiran. Karakterisasi genetik terhadap sifat-sifat prolifik ini telah banyak dilaporkan pada ternak Domba Jawa 19 ekor tipis, Merino, Thoka, Lacaune, Cambridge (Davis et al. 2002); Awassi dan Assap (Gootwine et al. 2008); Han ekor pendek (Chu et al. 2006); Rumpun Aragonesa (Rovo 2008), sedangkan untuk jenis ternak kambing baru dilaporkan pada Kambing Jining Grey (Chu et al. 2007). Meskipun demikian, kedua jenis ternak tersebut seringkali ditemukan mempunyai jumlah anak per kelahiran yang lebih dari satu (Odubute et al. 1992). Lan (2007) melaporkan bahwa laju ovulasi dan litter size pada Domba Booroola dari Merino Australia disebabkan alel FecBB dari major gene yang disebut FecB. Ditemukan pula bahwa gen utama tersebut berada pada ovine chromosome 6 yang juga merupakan posisi dari gen yang menyandikan salah satu anggota growth transforming factor-β, yaitu bone morphogenetik protein receptor 1B (BMPR1B). Hanrahan et al. (2004) kemudian menambahkan bahwa ada juga jenis gen lain yang berasosiasi dengan laju ovulasi dan litter size, yaitu Oocyte- growth derived factors GDF9 dan bone morphogenetik protein 15 pada Domba Cambridge dan Berclare. Dalam perkembangan berikutnya, ternyata ketiga jenis gen tersebut diketahui beraksi sebagai gen-gen utama yang mengatur tingkat laju ovulasi pada kisaran taksa mamalia yang luas, mulai dari manusia, sapi, tikus, dan kucing (Davis 2004; 2005) yang kemudian diikuti dengan penamaan gen yang bersinonim. Jadi dalam keluarga transforming growth factor-β yang berasosiasi dengan sifat-sifat prolifik adalah bone morphogenetik protein receptor type 1B (EU 581862 BMPR1B, activin-like kinase 6, atau FecB) pada ovine chromosome 6, growth differentiation factor 9 (= GDF9, Oocyte-derived growth factors, atau FecG) pada ovine chromosome 5, dan bone morphogenetik protein 15 (BMP15, atau FecX) pada ovine chromosome X. Selain ketiga gen fekunditas yang populer pada domba, gen prolactin receptor juga diduga berasosiasi dengan sifat prolifik pada Kambing Jining Grey (Zhang et al. 2007) dan pada manusia dan sapi. Berbagai analisis terhadap pola-pola mutasi dari gen-gen fekunditas dan asosiasinya dengan laju ovulasi, litter size dan berbagai efisiensi reproduksi lainnya banyak menarik perhatian para pemulia ternak (Davis 2005). Akibatnya bisa dipastikan bahwa pola-pola mutasi nukleotida pada gen-gen fekunditas diatas memunculkan berbagai penanda molekular yang diasosiasikan dengan berbagai sifat prolifik. KARAKTERISASI MORFOMETRIK DAN ANALISIS FILOGENI PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA Pendahuluan Berdasarkan Statistik Tahun 2010 jumlah populasi ternak kambing di Indonesia sebanyak 16 841 149 ekor, paling tinggi di provinsi Jawa Tengah (3 491 073 ekor), Jawa Timur (2 822 534 ekor), Jawa Barat (1 825 748 ekor), Lampung (1 206 000 ekor), Banten (854 522 ekor), NAD (886 468 ekor), Sumatera Utara (621 492 ekor), NTT (556 190 ekor) dan Sulawesi Selatan (442 297 ekor). Hampir 99% ternak ruminansia kecil di Indonesia merupakan skala usaha ternak kecil (Soedjana 2008). Sekitar 95% penduduk Indonesia adalah Muslim, ruminansia kecil mempunyai peranan penting pada kegiatan keagamaan terutama perayaan Idul Adha. Ternak kambing dapat mengkonversi hijauan berkualitas rendah menjadi protein hewani, sebagai sumber pupuk kandang serta sebagai tabungan. Parameter fenotipik merupakan metode yang paling mudah digunakan untuk mengidentifikasi karakterisitik ternak ruminansia (Alade et al. 2008; Khan et al. 2006; Dossa et al. 2007; Jimmy et al. 2010). Perbedaan penampilan disebabkan selama domestikasi tipe-tipe atau rumpun-rumpun hewan terpisah fenotipik secara genetik karena adanya proses adaptasi (ekpresi gen) dengan lingkungan lokal dan kebutuhan komunitas lokal sehingga dihasilkan rumpun yang berbeda. Adanya kemampuan adaptasi hewan disebabkan hewan memiliki kemampuan menghasilkan lebih dari satu alternatif bentuk morfologi, status fisiologi dan atau tingkah laku sebagai reaksi atau upaya adaptasi terhadap perubahan lingkungan berupa pengaturan ekspresi gen dan perubahan bentuk fenotip (Riva et al. 2004; Mansjoer et al . 2007; Noor 2008; Karna et al. 2001). Mendukung upaya pelestarian dan pemanfaatan ternak kambing lokal secara berkelanjutan maka perlu diketahui karakteristik fenotipik dan potensi produksi ternak yang ada di Indonesia. Tujuan penelitian ini adalah untuk mendapatkan data dan informasi tentang karakteristik morfometrik dan jarak genetik kambing lokal di Indonesia. 21 Bahan dan Metode Penelitian ini menggunakan beberapa sub populasi kambing lokal Indonesia yang disebut Kambing Samosir, Kambing Muara, Kambing Marica, Kambing Jawarandu, Kambing Benggala dan Kambing Kacang. Penentuan sampel kambing dengan metode purposive sampling, yaitu pertama menentukan Kabupaten daerah sentra produksi di setiap Propinsi, baru kemudian ditentukan Kecamatan dan Kelompok Desa. Setiap sub populasi diambil sekitar 34 - 217 ekor kambing sebagai sampel sesuai dengan ketersediaan populasi ternak yang bisa ditemui di lapangan dan diusahakan diambil dari desa yang jauh kekerabatan/keturunan sampel kambing dengan sampel pada lokasi desa pengambilan lainnya. Peralatan penelitian yang digunakan yaitu tongkat ukur ketelitian 0.1 cm, pita ukur ketelitian 0.1 cm, jangka sorong stainless steel buatan Jerman, timbangan gantung (shelter) dengan ukuran kg dengan tingkat ketelitian 50 gram, kamera digital Nikon F-9. 8 mega pixel, dan tali rapiah pengikat kambing. Waktu dan Tempat Penelitian Penelitian lapangan untuk koleksi data fenotipik dilakukan pada bulan Maret 2009 sampai Maret 2011 di empat Propinsi yaitu: 1. Propinsi Sumatera Utara : Kabupaten Samosir, Kabupaten Tapanuli Utara, Kabupaten Deli Serdang. 2. Propinsi Jawa Tengah; Kabupaten Blora. 3. Propinsi Sulawesi Selatan: Kabupaten Maros, Kota Makassar, Kabupaten Jeneponto. 4. Propinsi Nusa Tenggara Timur : Kota Kupang, Kabupaten Kupang, Kabupaten Sikka dan Kabupaten Ende. 22 •KACANG (n=217) Kab. Deli Serdang, • MUARA (n=34) Kab. Tap.Utara, •SAMOSIR (n=42) Kab. Samosir PROPINSI SUMATERA UTARA JAWARANDU (n=94) Kab. Blora PROPINSI JAWA TENGAH MARICA (n=60) Kab. Maros, Kab. Jeneponto, Kota Makassar PROPINSI SULAWESI SELATAN BENGGALA (n=96) Kab. Kupang, Kab. Sikka, Kab. Ende, PROPINSI NUSA TENGGARA TIMUR Gambar 4 Peta lokasi pengambilan sampel dan data penelitian karakterisasi enam sub populasi kambing lokal Indonesia. Pengumpulan Sampel Kambing Teknik pengambilan sampel ternak kambing dilakukan secara acak pada 543 ekor kambing, yaitu 96 ekor Kambing Benggala (betina=89, jantan=7), 94 ekor Kambing Jawarandu (betina=72, jantan=22), 60 ekor Kambing Marica (betina=48, jantan=12), 217 ekor Kambing Kacang (betina=193, jantan=24), 34 ekor Kambing Muara (betina=28, jantan=6) dan 42 ekor Kambing Samosir (betina=36, jantan=6). Pengumpulan data fenotipik dilakukan bersamaan dengan pengambilan sampel darah. Koleksi data dimulai dengan pencatatan jenis kelamin dan umur kambing serta nama pemiliknya. Umur kambing penelitian ditentukan berdasarkan minimal sudah terdapat 1 pasang gigi seri yang permanen. Parameter fenotipik yang digunakan dalam analisis data meliputi : 1) lingkar dada (LIDA), diukur melingkar tepat di belakang scapula, dengan menggunakan pita ukur dalam cm; 23 2) lebar dada (LEDA), diukur antara tuberitas humeri sinister dan dexter, dengan menggunakan tongkat ukur dalam cm; 3) dalam dada (DADA), diukur dari bagian tertinggi pundak sampai dasar dada, dengan menggunakan tongkat ukur dalam cm; 4) tinggi pundak (TIPU), diukur dari bagian tertinggi pundak melalui belakang scapula tegak lurus ke tanah, dengan menggunakan tongkat ukur dalam cm; 5) tinggi pinggul (TIPI), diukur dari bagian tertinggi pinggul secara tegak lurus ke tanah, dengan menggunakan tongkat ukur dalam cm; 6) lebar pinggul (LEPI), diukur dengan jarak lebar antara kedua sendi pinggul dengan menggunakan tongkat ukur dalam cm; 7) panjang badan (PABA), diukur dari tuber ischii sampai tuberitas humeri, dengan menggunakan tongkat ukur dalam cm; 8) lingkar kanon (LIKA), diukur tepat melingkar pada bagian tulang canon kaki belakang sebelah kiri dengan menggunakan pita ukur dalam cm; 9) lebar kanon (LEKA) diukur jarak antar tulang kering lutut dengan tulang kanon; dengan menggunakan pita ukur dalam cm 10) panjang ekor (PAEK), diukur pada pangkal sampai ujung ekor, dengan menggunakan pita ukur dalam cm 11) lebar ekor (LEEK), diukur lebar ekor pada bagian paha ekor, dengan menggunakan jangka sorong dalam cm; 12) tebal ekor (TEEK), diukur tebal pada bagian pangkal ekor, dengan menggunakan jangka sorong dalam cm; 13) panjang telinga (PATEL), diukur pada pangkal telinga sampai ujung telinga; dengan menggunakan pita ukur dalam cm 14) lebar telinga (LETEL) diukur lebar telinga pada bagian paling lebar, dengan menggunakan jangka sorong dalam cm; 15) panjang tengkorak (PATEK), diukur pada posisi tengah kepala diantara dua tanduk sampai ke bagian mulut menghitam, menggunakan pita ukur dalam cm; 16) lebar tengkorak (LETEK), diukur dengan jarak kedua sisi tulang pipi, dengan menggunakan pita ukur dalam cm; 17) tinggi tengkorak (TITEK), diukur mulai dari sudut rahang bawah sampai bagian atas sisi paling atas tegak lurus, dengan menggunakan pita ukur dalam cm; 24 18) panjang tanduk (PATA), diukur pada pangkal tanduk sampai ujung tanduk mengikuti arah pertumbuhan tanduk dengan menggunakan pita ukur dalam cm (Lanari et al. 2003 ; Abdullah 2008). Penimbangan berat badan dilakukan sebelum pengukuran ukuran tubuh, dengan menggunakan timbangan gantung (shelter) dengan ukuran kg (tingkat ketelitian 50 gram), dilaksanakan pada pagi hari sebelum makan. Sifat-sifat fenotip kualitatif yang diamati yaitu warna dominan, warna belang tubuh yang dikelompokkan menurut lokasi dan jenis kelamin. Pengamatan bentuk tanduk dengan cara mengamati arah pertumbuhannya berawal dari kepala sampai ujung tanduk. Setiap individu dicatat arah pertumbuhannya dan dibuat sketsa dari pertumbuhan tanduk tersebut. Bagian-bagian permukaan tubuh kambing yang diukur (cm) dapat dilihat pada Gambar 5. Keterangan: 1. Lingkar dada 2. Lebar dada 3. Dalam dada 4. Tinggi pundak 5. Tinggi pinggul 6. Lebar pinggul 7. Panjang badan 8. Lingkar kanon 9. Panjang telinga 10.Panjang ekor 11.Lebar ekor 12.Tebal ekor 13.Panjang tengkorak 14.Lebar tengkorak 15.Tinggi tengkorak 16.Panjang tanduk 17.Lebar telinga 9 Gambar 5 Titik pengukuran morfometrik kambing Analisis Statistik Analisis ragam (ANOVA) digunakan untuk mempelajari pengaruh ukuranukuran tubuh antar lokasi dengan model matematis menurut Mattjik dan Sumertajaya (2006) sebagai berikut: Y ij = µ + τ i + Ɛ ij 25 Keterangan: Y ij = respon peubah yang diamati µ = pengaruh genotip ke-I (i=1, 2, 3,….) τ i = pengaruh galat perlakuan ke-i ulangan ke-j Ɛ ij = respon peubah yang diamati = rataan umum Data dikoreksi untuk menghilangkan pengaruh jumlah lebih kecil sampel kambing jantan tidak seimbang jumlahnya dibandingkan dengan jumlah sampel betina. Analisis nilai rataan, simpangan baku dan analisis ragam (ANOVA) dilakukan dengan menggunakan perangkat lunak statistik Minitab V.21. Jika hasil analisis berbeda nyata, maka dilakukan uji lanjut Duncan. Analisis Morfometrik Fungsi diskriminan sederhana dilakukan untuk penentuan jarak genetik (Traore et al. 2008). Fungsi diskriminan yang digunakan melalui pendekatan jarak Mahalanobis seperti yang dijelaskan oleh Mailund et al. (2008), dimana matriks ragam peragamam antara peubah dari masing-masing tipe kambing yang diamati digabungkan (pooled) menjadi sebuah matriks. Matriks pooled dapat dijelaskan ke dalam bentuk berikut: c11 c 21 C= c 31 cp1 c12 c 22 c 32 cp 2 c1 p c2 p c3 p cpp .... .... .... .... Mendapatkan jarak kuadrat genetik minimum digunakan rumus sesuai dengan petunjuk Everitt et al. (2001) dan Quinn et al. (2002) sebagai berikut: D2 ( i, j ) = ( i - j ) C-1 ( i - j ) Keterangan: D2 ( i, j) = Nilai statistic Mahalanobis sebagai ukuran jarak kuadrat genetik antara dua rumpun/genotip kambing (antara genotip ke-i terhadap genotip ke-j). C-1 i = Kebalikan matrik gabungan ragam peragam antar peubah. = Vektor nilai rataan pengamatan dari genotip kambing ke-i pada masing-masing peubah kuantitatif. j = Vektor nilai rataan pengamatan dari genotip kambing ke-j pada masing-masing peubah kuantitatif. 26 Analisis statistik Mahalanobis dengan menggunakan paket program SAS versi 9.1 (SAS Inst. 2005) prosedur PROC CANDISC dan PROC DISCRIM. Dari hasil perhitungan jarak kuadrat tersebut, kemudian dilakukan pengakaran terhadap hasil kuadrat jarak, agar jarak genetik yang didapat bukan dalam bentuk kuadrat. Hasil pengakaran dianalisis lebih lanjut dengan program MEGA versi 4.0 seperti petunjuk Tamura et al. (2007) untuk mendapatkan pohon fenogram. Analisis kanonikal (Crepaldi et al. 2001) dilakukan untuk penentuan peta penyebaran kambing dan nilai kesamaan dan campuran di dalam dan di antara kelompok kambing. Hasil dan Pembahasan Bobot Badan Rataan bobot badan betina paling tinggi pada Kambing Muara (37.46±5.42 kg) berbeda nyata (P<0.05) jika dibandingkan dengan 5 sub populasi kambing lokal lainnya. Rataan bobot badan Kambing Samosir betina adalah 25±5.24 kg hampir sama dengan Kambing Benggala dan Jawarandu, dan berbeda nyata (P<0.05) lebih tinggi jika dibandingkan dengan Kambing Kacang dan Marica. Rataan dan simpangan baku bobot hidup kambing pengamatan diperlihatkan pada Tabel 1. Tabel 1 Rataan, simpangan baku dan kisaran bobot hidup dewasa 6 sub populasi kambing lokal. Sub Betina Jantan Gabungan populasi n n n kk (%) ± s (kg) ± s (kg) ± s (kg) b c b B 89 24.73 ± 8.69 7 16.00 ± 3.87 96 24.09 ± 8.72 36.20 bc c c J 72 23.11 ± 7.87 22 16.36 ± 4.79 94 21.15 ± 7.79 36.19 c b bc K 193 21.61 ± 5.86 24 24.67 ± 6.09 217 21.95 ± 5.95 27.12 c bc c M 48 20.88 ± 6.61 12 19.17 ± 5.27 60 20.53 ± 6.36 30.98 a a a R 28 37.46 ± 11.01 6 49.00 ± 26.87 34 38.23 ± 12.10 31.64 b bc b S 36 25.00 ± 5.42 6 22.00 ± 8.10 42 24.57 ± 5.86 23.82 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0.05); K= Kacang; B= Benggala; S= Samosir; J= Jawa randu; M= Marica; R= Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Rataan bobot badan jantan paling tinggi didapatkan juga pada Kambing Muara (49±26.87 kg) berbeda nyata lebih tinggi jika dibandingkan dengan 5 sub populasi kambing lainnya. Rataan bobot badan Kambing Kacang jantan 24.67±6.09 kg hampir sama dengan Kambing Samosir dan Marica berbeda nyata. Rataan bobot badan jantan pada sub populasi Kambing Kacang dan Muara lebih tinggi jika dibandingkan dengan bobot badan kambing betina. Pada 27 sub populasi Kambing Benggala, Jawarandu, Marica dan Samosir terdapat sebaran data sampel bobot badan yang tidak normal karena rataan bobot badan kambing jantan lebih rendah jika dibandingkan dengan bobot badan betina, hal ini disebabkan pada keempat sub populasi kambing tersebut kambing jantan biasanya sering dijual oleh peternak lebih cepat. Jumlah kambing jantan yang dipelihara sangat terbatas dan umurnya relatif masih muda, sehingga data yang diperoleh sangat terbatas jumlahnya. Berdasarkan kondisi data ini diduga para peternak tradisional di daerah pedesaan masih kurang memperhatikan perlunya bibit pejantan kambing yang baik untuk sistim perkawinan. Rataan bobot badan betina dan jantan pada pada Kambing Kacang dan Jawarandu pada penelitian ini lebih rendah jika dibandingkan dengan hasil penelitian Setiadi et al. (1997) di Kabupaten Purworejo yaitu 23.83 kg dan 26.88 kg untuk Kambing Kacang, 28.74 kg dan 30.91 kg untuk Kambing Jawarandu. Rataan bobot badan Kambing Jawarandu sangat berbeda diduga karena kualitas bibit dan cara pemeliharaan di Kabupaten Purworejo lebih baik dibandingkan dengan di Kabupaten Blora yang pada umumnya kambing dilepas atau diumbar pada siang hari dan pada malam hari dikandangkan, sistim pemberian pakan pada umumnya hanya mengandalkan rumput alam saja. Jika digabungkan rataan bobot badan betina dan jantan menunjukkan bahwa bobot badan Kambing Muara (38.23 ± 12.10 kg) berbeda nyata (P<0.05) paling tinggi jika dibandingkan dengan 5 sub populasi kambing lokal lainnya. Rataan bobot badan Kambing Samosir (24.57 ± 5.86 kg) hampir sama dengan Kambing Benggala (24.09 ± 8.72 kg) dan Kacang (21.95±5.95 kg), berbeda nyata (P<0.05) lebih tinggi jika dibandingkan dengan Kambing Jawarandu (21.15 ± 7.79 kg) dan Marica (20.53 ± 6.36 kg). Tingkat keragaman bobot badan keenam kambing lokal yang diamati sangat beragam, dapat dilihat berdasarkan koefisien keragaman rataan bobot badan berkisar 23.82 %-36.20 %. Parameter Ukuran Tubuh Rataan dan simpangan baku ukuran tinggi pundak, panjang badan, lebar dada, dalam dada dan lingkar dada jantan dan betina dewasa 6 sub populasi kambing lokal disajikan pada Tabel 2. Hasil pengukuran menunjukkan rataan parameter ukuran-ukuran tubuh kambing antar sub populasi kambing secara umum berbeda nyata (P<0.05). Ukuran tinggi pundak Kambing Muara betina (65.29 ± 3.82 cm) berbeda nyata (P<0.05) paling tinggi jika dibandingkan dengan 28 kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang, Kambing Benggala dan Samosir. Tabel 2 Rataan dan simpangan baku ukuran tinggi pundak, panjang badan, lebar dada, dalam dada, dan lingkar dada kambing jantan dan betina 6 sub populasi kambing lokal Ukuran Tubuh Tinggi Pundak Sub Betina Jantan populasi n kk (%) n kk (%) ± s (cm) ± s (cm) b c B 55.30 ± 7.05 89 12.76 47.71 ± 4.89 7 10.25 cd c J 52.47 ± 7.69 72 14.65 48.91 ± 6.88 22 14.07 b b K 55.62 ± 4.22 193 7.58 56.33 ± 4.44 24 7.88 d bc M 51.42 ± 5.15 48 10.02 51.17 ± 5.86 12 11.45 a a R 65.29 ± 3.82 28 5.58 78.00 ± 11.31 6 14.50 bc bc S 54.50 ± 4.35 36 7.97 50.56 ± 5.09 6 10.04 bc bc Panjang B 61.56 ± 9.12 89 14.81 51.41 ± 3.98 7 7.77 d c Badan J 53.06 ± 11.29 72 21.29 46.36 ± 6.51 22 14.03 c b K 58.87 ± 5.58 193 9.47 58.00 ± 3.01 24 5.18 d b M 54.92 ± 5.09 48 9.26 58.67 ± 14.33 12 24.43 a a R 72.82 ± 6.99 28 9.60 76.50 ± 14.85 6 19.41 b b S 63.44 ± 5.07 36 8.00 59.33 ± 7.89 6 13.30 b ab Lebar B 11.48 ± 4.83 89 42.03 17.71 ± 3.77 7 21.30 b c Dada J 12.60 ± 3.64 72 28.87 10.36 ± 3.72 22 35.92 b b K 11.61 ± 2.14 193 18.40 15.00 ± 2.64 24 17.58 a b M 16.25 ± 3.19 48 19.60 15.83 ± 3.10 12 19.57 a a R 15.11 ± 3.37 28 27.54 21.00 ± 3.49 6 53.03 a b S 15.25 ± 2.29 36 14.98 14.50 ± 3.51 6 24.19 a b Dalam B 20.56 ± 5.56 89 27.05 27.71 ± 3.77 7 13.62 b b Dada J 27.33 ± 4.07 72 14.90 24.82 ± 4.20 22 16.94 c b K 25.61 ± 2.14 193 8.34 29.00 ± 2.64 24 9.09 b b M 27.25 ± 3.19 48 11.69 26.83 ± 3.10 12 11.15 a a R 30.25 ± 3.37 28 11.15 34.00 ± 8.49 6 24.96 bc b S 26.25 ± 2.29 25.50 ± 3.51 13.75 36 8.70 6 b bc Lingkar B 68.35 ± 7.81 89 11.43 57.86 ± 5.52 7 9.54 c c Dada J 64.28 ± 9.62 72 14.97 54.73 ± 7.34 22 13.42 c b K 63.15 ± 7.03 193 11.13 66.67 ± 5.16 24 7.75 c bc M 63.71 ± 6.81 48 10.69 61.33 ± 7.35 12 11.99 a a R 79.93 ± 8.19 28 10.24 85.50 ± 17.68 6 20.68 bc bc S 66.00 ± 6.13 36 9.29 59.83 ± 12.77 6 21.34 c b Lebar B 15.90 ± 2.88 89 18.09 16.86 ± 2.91 7 17.27 c c Pinggul J 15.28 ± 2.08 72 13.60 13.82 ± 1.62 22 11.74 d c K 12.21 ± 2.28 193 18.69 13.67 ± 0.96 24 7.05 c b M 16.75 ± 2.11 48 12.59 16.33 ± 1.87 12 11.48 a a R 20.00 ± 2.51 28 12.55 19.50 ± 0.71 6 3.63 b a S 18.97 ± 2.34 36 12.31 19.83 ± 2.48 6 12.52 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0.05); K= Kacang; B= Benggala; S= Samosir; J= Jawa randu; M= Marica; R= Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Tinggi pundak paling rendah dijumpai pada Kambing Jawarandu dan Marica. Tinggi pundak jantan dewasa paling tinggi juga pada Kambing Muara (78.00 ± 11.31 cm) berbeda nyata jika dibandingkan dengan kelima sub populasi 29 lainnya, kemudian disusul Kambing Kacang, Marica dan Samosir. Tinggi pundak paling rendah pada Kambing Benggala dan Jawarandu. Rataan panjang badan betina paling tinggi pada Kambing Muara (72.82 ± 6.99 cm) berbeda nyata (P<0.05) dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Samosir, Benggala dan Kacang. Panjang badan betina paling rendah pada Kambing Jawarandu dan Marica. Panjang badan jantan paling tinggi pada Kambing Muara (76.50 ± 14.85 cm), disusul Kambing Samosir, Marica, Kacang dan Benggala. Panjang badan jantan paling rendah pada Kambing Jawarandu (46.36 ± 6.51 cm). Lebar dada betina paling tinggi pada Kambing Marica (16.25 ± 3.19 cm), disusul Kambing Samosir dan Kambing Muara, yang berbeda nyata (P<0.05) dengan Kambing Jawarandu, Kacang dan Benggala. Lebar dada jantan dewasa paling tinggi pada Kambing Muara dan Benggala, kemudian disusul Kambing Marica, Kacang dan Samosir. Lebar dada jantan paling rendah pada Kambing Jawarandu (10.36 ± 3.72 cm). Dalam dada betina paling tinggi pada Kambing Muara dan Benggala berbeda nyata (P<0.05) dengan keempat sub populasi kambing lainnya, kemudian disusul Kambing Jawarandu, Marica dan Samosir. Dalam dada betina paling rendah pada Kambing Kacang. Dalam dada jantan paling tinggi pada Kambing Muara yang berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi lainnya. Dalam dada Kambing Samosir tidak berbeda nyata (P>0.05) jika dibandingkan antara Kambing Marica, Kacang, Jawarandu dan Kambing Benggala. Lingkar dada betina paling tinggi pada Kambing Muara (79.93 ± 8.19 cm) berbeda nyata (P<0.05) dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Benggala dan Samosir. Lingkar dada betina paling rendah pada Kambing Kacang, Marica dan Jawarandu. Lingkar dada jantan paling tinggi pada Kambing Muara (85.50 ± 17.68 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang, Marica, Samosir dan Benggala. Lingkar dada jantan paling rendah pada Kambing Jawarandu (54.73 ± 7.34 cm). Panjang tengkorak betina paling tinggi pada Kambing Kacang (15.68 ± 1.38 cm) dan Kambing Benggala (15.30 ± 1.88 cm) berbeda nyata (P<0.05) jika dibandingkan dengan keempat sub populasi kambing lainnya, kemudian disusul Kambing Muara dan Kambing Jawarandu. Tabel 3 menunjukkan rataan dan 30 simpangan baku ukuran panjang tengkorak, lebar tengkorak, dan tinggi tengkorak jantan dan betina dewasa 6 sub populasi kambing yang berbeda. Panjang tengkorak betina paling rendah pada Kambing Marica dan Samosir. Panjang tengkorak jantan dewasa paling tinggi pada Kambing Muara (16.00 ± 2.83 cm) dan Kacang (15.67 ± 1.40 cm) berbeda nyata jika dibandingkan dengan keempat sub populasi kambing lainnya, akan tetapi Kambing Benggala, Marica, Jawarandu dan Kambing Samosir tidak berbeda nyata (P>0.05). Tabel 3 Rataan dan simpangan baku ukuran panjang tengkorak, lebar tengkorak, dan tinggi tengkorak jantan dan betina dewasa 6 sub populasi kambing lokal Ukuran tubuh Sub Betina Jantan popu n kk (%) n kk (%) ± s (cm) ± s (cm) lasi ab b Panjang B 15.30 ± 1.88 89 12.28 12.86 ± 1.46 7 11.39 cd b tengkorak J 14.08 ± 2.52 72 17.87 12.36 ± 1.81 22 14.67 a a K 15.68 ± 1.38 193 8.78 15.67 ± 1.40 24 8.96 d b M 13.73 ± 1.77 48 12.90 13.00 ± 1.35 12 10.37 bc a R 14.64 ± 1.54 28 10.55 16.00 ± 2.83 6 17.68 d b S 13.69 ± 1.86 36 13.61 13.33 ± 1.97 6 14.75 cd d Lebar B 10.53 ± 1.30 89 12.33 8.71 ± 0.76 7 8.67 de d tengkorak J 10.28 ± 2.09 72 20.35 9.18 ± 1.14 22 12.41 bc b K 10.97 ± 1.19 193 10.81 12.33 ± 0.48 24 3.90 e d M 9.79 ± 1.27 48 12.98 8.67 ± 0.49 12 5.68 a a R 13.79 ± 2.91 28 21.11 13.50 ± 2.12 6 15.71 b c S 11.56 ± 2.45 36 21.24 10.67 ± 0.82 6 7.65 c cd Tinggi B 11.70 ± 1.39 89 11.84 11.29 ± 1.11 7 9.86 c d J 11.14 ± 1.78 72 15.97 10.45 ± 1.65 22 15.82 tengkorak b b K 12.58 ± 1.73 193 13.78 15.00 ± 1.87 24 12.43 c cd M 11.58 ± 1.20 48 10.36 11.33 ± 1.15 12 10.19 a a R 14.36 ± 1.47 28 10.25 17.00 ± 2.83 6 16.64 b c S 12.94 ± 1.19 36 9.22 12.50 ± 1.05 6 8.39 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0.05); K= Kacang; B= Benggala; S= Samosir; J= Jawa randu; M= Marica; R= Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Lebar tengkorak betina paling tinggi pada Kambing Muara (13.79 ± 2.91 cm), yang berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Samosir, Kacang, Benggala dan Jawarandu. Lebar tengkorak betina paling rendah pada Kambing Marica. Lebar tengkorak jantan paling tinggi pada Kambing Muara (13.50 ± 2.12 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang dan Samosir. Lebar tengkorak jantan tiga urutan terendah Kambing Marica, Benggala dan Jawarandu. Tinggi tengkorak betina paling tinggi pada Kambing Muara (14.36 ± 1.47 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi 31 kambing lainnya, kemudian disusul Kambing Samosir dan Kacang. Tinggi tengkorak betina tiga urutan terendah pada Kambing Jawarandu, Marica dan Kambing Benggala. Tinggi tengkorak jantan paling tinggi pada Kambing Muara (17.00 ± 2.83 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang (15.00 ± 1.87 cm), Kambing Samosir (12.50 ± 1.05 cm), Kambing Marica (11.33 ± 1.15 cm) dan Kambing Benggala (11.29 ± 1.11 cm). Ukuran tinggi tengkorak jantan dewasa paling rendah pada Kambing Jawarandu (10.45 ± 1.65cm). Parameter rataan ukuran dan simpangan baku panjang dan lebar ekor jantan dan betina dewasa 6 sub populasi kambing pada sub populasi yang berbeda ditampilkan pada Tabel 4. Tabel 4 Rataan dan Simpangan Baku panjang dan lebar ekor jantan dan betina 6 sub populasi kambing lokal Ukuran Sub Betina Jantan tubuh populasi n kk (%) n kk (%) ± s (cm) ± s (cm) b c Panjang B 10.22 ± 2.79 89 27.30 7.86 ± 4.41 7 56.17 b bc ekor J 11.22 ± 1.79 72 15.98 10.27 ± 2.31 22 22.52 b b K 11.40 ± 6.47 193 56.72 11.50 ± 0.98 24 8.50 b c M 10.13 ± 1.18 48 11.64 9.17 b ± 0.72 12 7.83 a a R 13.96 ± 1.73 28 12.40 16.50 ± 3.54 6 21.43 b bc S 10.08 ± 1.99 36 19.75 9.50 ± 1.05 6 11.04 c c Lebar B 3.92 ± 1.71 89 43.53 2.71 ± 0.49 7 17.98 e c ekor J 1.90 ± 0.70 72 36.54 1.82 ± 0.39 22 21.71 b b K 4.73 ± 0.79 193 16.71 4.33 ± 1.13 24 26.06 d c M 2.35 ± 0.48 48 20.53 2.17 ± 0.72 12 33.13 a a R 5.29 ± 1.21 28 22.94 5.50 ± 0.71 6 12.86 c b S 4.11 ± 0.71 36 17.23 3.67 ± 0.82 6 22.27 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0.05); K= Kacang; B= Benggala; S= Samosir; J= Jawa randu; M= Marica; R= Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Secara umum rataan ukuran panjang dan lebar ekor kambing antara 6 sub populasi kambing berbeda nyata (P<0.05) jika dibandingkan antara satu sama lainnya. Rataan ukuran panjang ekor betina dewasa paling tinggi pada Kambing Muara (13.96 ± 1.73 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya. Ukuran rataan panjang ekor betina dewasa pada Kambing Jawarandu, Marica, Samosir, Kacang dan Kambing Benggala tidak berbeda nyata (P>0.05). Ukuran rataan panjang ekor jantan dewasa paling tinggi dijumpai pada Kambing Muara (16.50 ± 3.54 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang (15.00 ± 1.87 cm), Kambing Jawarandu (10.27 ± 2.31 cm), kambing Samosir (9.50 ± 1.05 cm) dan Kambing 32 Marica (9.17 ± 0.72 cm). Ukuran panjang ekor jantan dewasa paling rendah pada Kambing Benggala (7.86 ± 4.41 cm). Rataan ukuran lebar ekor betina dewasa paling tinggi pada Kambing Muara (5.29 ± 1.21 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang (4.73 ± 0.79), Kambing Samosir (4.11 ± 0.71 cm), Kambing Benggala (3.92 ± 1.71) dan Kambing Marica (2.35 ± 0.48 cm). Ukuran rataan lebar ekor betina dewasa paling rendah pada Kambing Jawarandu (1.90 ± 0.70 cm). Ukuran rataan lebar ekor jantan dewasa paling tinggi dijumpai pada Kambing Muara (5.50 ± 0.71 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang (4.33 ± 1.13 cm) dan kambing Samosir (9.50 ± 1.05 cm). Ukuran lebar ekor jantan dewasa paling rendah pada Kambing Jawarandu (1.82 ± 0.39 cm), Kambing Marica (2.17 ± 0.72 cm) dan Kambing Benggala (2.71 ± 0.49 cm). Tabel 5 Rataan dan simpangan baku panjang telinga dan lebar telinga jantan dan betina dewasa 6 sub populasi kambing lokal Betina Jantan Sub populasi n kk (%) kk (%) ± s (cm) ± s (cm) n c c B 14.63 ± 2.94 89 20.12 11.86 ± 1.46 7 12.35 b b J 15.47 ± 1.75 72 11.32 14.64 ± 2.06 22 14.07 b b K 16.08 ± 1.96 193 12.16 14.00 ± 1.02 24 7.30 d bc M 13.38 ± 1.33 48 9.95 13.50 ± 1.98 12 14.65 a a R 19.14 ± 2.86 28 14.96 21.00 ± 1.41 6 6.73 cd b S 13.92 ± 1.87 36 13.46 13.83 ± 1.17 6 8.45 c c Lebar B 15.51 ± 3.44 89 22.17 10.29 ± 3.99 7 38.77 c b telinga J 15.28 ± 2.08 72 13.60 14.27 ± 1.75 22 12.27 d b K 12.10 ± 2.29 193 18.93 13.50 ± 1.93 24 14.32 b b M 16.83 ± 1.99 48 11.84 15.92 ± 1.73 12 10.87 a a R 20.00 ± 2.51 28 12.55 19.50 ± 0.71 6 3.63 a a S 19.19 ± 2.21 36 11.54 18.83 ± 3.13 6 16.59 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0.05); K= Kacang; B= Benggala; S= Samosir; J= Jawa randu; M= Marica; R= Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Ukuran tubuh Panjang telinga Pada Tabel 5 ditampilkan parameter ukuran rataan dan simpangan baku panjang telinga dan lebar telinga jantan dan betina dewasa 6 sub populasi kambing yang berbeda. Secara umum hasil menunjukkan rataan ukuran telinga kambing antar sub populasi berbeda nyata (P<0.05). Parameter ukuran tubuh panjang telinga betina dewasa paling tinggi pada Kambing Muara (19.14 ± 2.86 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Kacang (16.08 ± 1.96 cm), Kambing Jawarandu (15.47 ± 1.75 cm), Kambing Benggala (14.63 ± 2.94 cm) dan 33 Kambing Samosir (13.92 ± 1.87 cm). Ukuran panjang telinga betina dewasa paling rendah pada Kambing Marica (13.38 ± 1.33 cm). Ukuran panjang telinga jantan dewasa paling tinggi pada Kambing Muara (21.00 ± 1.41 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Jawarandu (14.64 ± 2.06cm), Kambing Kacang (14.00 ± 1.02 cm), Kambing Samosir (13.83 ± 1.17 cm) dan Kambing Marica (13.50 ± 1.98 cm). Ukuran panjang telinga jantan dewasa paling rendah pada Kambing Benggala (11.86 ± 1.46 cm). Parameter ukuran lebar telinga betina dewasa paling tinggi pada Kambing Muara (20.00 ± 2.51 cm) dan Kambing Samosir (19.19 ± 2.21 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Marica (16.83 ± 1.99 cm), Kambing Benggala (15.51 ± 3.44 cm), dan Kambing Jawarandu (15.28 ± 2.08 cm). Ukuran lebar telinga betina dewasa paling rendah pada Kambing Kacang (12.10 ± 2.29 cm). Ukuran lebar telinga jantan dewasa paling tinggi pada Kambing Muara (21.00 ± 1.41 19.50 ± 0.71 cm) dan Kambing Samosir (18.83 ± 3.13 cm) berbeda nyata (P<0.05) jika dibandingkan dengan keempat sub populasi kambing lainnya, kemudian disusul Kambing Marica (15.92 ± 1.73 cm), Kambing Jawarandu (14.27 ± 1.75 cm) dan Kambing Kacang (13.50 ± 1.93 cm). Ukuran lebar telinga jantan dewasa paling rendah pada Kambing Benggala (10.29 ± 3.99 cm). Parameter ukuran Rataan dan simpangan baku lingkar kanon jantan dan betina dewasa 6 sub populasi kambing disajikan pada Tabel 6. Secara umum ukuran lingkar kanon antar sub populasi berbeda nyata (P<0.05). Parameter ukuran tubuh lingkar kanon betina dewasa paling tinggi pada Kambing Muara (16.71 ± 1.46 cm) berbeda nyata (P<0.05) jika dibandingkan dengan kelima sub populasi kambing lainnya, kemudian disusul Kambing Benggala (14.64 ± 1.65 cm), Kambing Samosir (14.11 ± 1.26 cm), Kambing Jawarandu (13.83 ± 1.67 cm) dan Kambing Marica (13.29 ± 1.07 cm). Ukuran lingkar kanon betina dewasa paling rendah pada Kambing Kacang (7.73 ± 0.69 cm). Ukuran lingkar kanon jantan dewasa paling tinggi pada Kambing Muara (19.50 ± 4.95 cm) berbeda nyata (P<0.05) jika dibandingkan dengan keempat sub populasi kambing lainnya, kemudian disusul Kambing Samosir (14.67 ± 0.82cm), Kambing Marica (13.83 ± 0.94 cm), Kambing Benggala (13.43 ± 0.53 cm) dan Kambing Jawarandu (13.27 ± 1.58 34 cm). Ukuran lingkar kanon jantan dewasa paling rendah pada Kambing Kacang (9.17 ± 0.38 cm). Tabel 6. Rataan dan simpangan baku lingkar kanon jantan dan betina 6 sub populasi kambing lokal Ukuran tubuh Lingkar kanon Sub Betina Jantan Populasi n kk (%) n kk (%) ± s (cm) ± s (cm) b b B 14.64 ± 1.65 89 11.24 13.43 ± 0.53 7 3.98 c b J 13.83 ± 1.67 72 12.07 13.27 ± 1.58 22 11.90 e c K 7.73 ± 0.69 193 8.98 9.17 ± 0.8 24 4.15 d b M 13.29 ± 1.07 48 8.06 13.83 ± 0.94 12 6.78 a a R 16.71 ± 1.46 28 8.75 19.50 ± 4.95 6 25.38 c b S 14.11 ± 1.26 36 8.93 14.67 ± 0.82 6 5.57 Huruf superskrip yang berbeda pada kolom yang sama menyatakan berbeda nyata (P<0,05); K=Kacang; B=Benggala; S=Samosir; J=Jawarandu; M= Marica; R=Muara; n= jumlah sampel; = rataan; s= simpangan baku; kk= koefisien keragaman Perbedaan ukuran-ukuran tubuh ini disebabkan laju pertumbuhan ukuranukuran tubuh ternak kambing yang dipengaruhi oleh faktor genetik dan lingkungan. Suparyanto et al. (1999) menyatakan bahwa karakteristik ukuranukuran tubuh dapat menggambarkan ciri khas dari suatu bangsa. Selain perbedaan secara genetik dan lingkungan yang dapat berupa adanya perbedaan iklim, hal lainnya yang dapat mempengaruhi karakteristik ukuran-ukuran tubuh tersebut adalah manajemen pemeliharaan di setiap lokasi yang berbeda-beda. Plot Penyebaran Kambing menurut Ukuran Fenotipik Hasil analisis morfologi menunjukkan bahwa pada keenam sub populasi kambing penelitian memperlihatkan adanya keragaman yang tinggi. Keragaman sifat morfologi dapat terjadi karena adanya proses seleksi (alam dan buatan), perkawinan silang atau bencana alam yang dapat berakibat hilang atau hanyutnya gen tertentu (Anderson 2001). Gambar 6 menunjukkan bahwa kambing dari keenam sub populasi kambing penelitian dibagi menjadi 6 kelompok, yaitu kelompok Kambing Muara (R) ada di kuadran II, kelompok Kambing Jawarandu (J) ada sebagian besar di kuadran II dan kuadran III, kelompok Kambing Kacang ada di kuadran I dan IV, kelompok Kambing Benggala sebagian besar ada di kuadran III dan sebagian kecil ada di kuadran II, kelompok Kambing Marica ada di kuadran II dan kuadran III, dan kelompok Kambing Samosir ada di kuadran III. Sub populasi Kambing Kacang merupakan kelompok yang jauh terpisah bergeser ke kiri di kuadran I dan IV jika dibandingkan dengan sub populasi lainnya. Kambing Kacang diduga mempunyai 35 ukuran-ukuran tubuh relatip lebih kecil, seperti lingkar kanon, lebar pinggul dan lebar dada. Karakteristik ukuran tubuh Kambing Marica di Propinsi Sulawesi Selatan, Kambing Samosir di Kabupaten Tapanuli Utara, Propinsi Sumatera Utara berdekatan, relatip sama. Hal ini diduga karena adanya proses adaptasi terhadap kondisi fisik lingkungan juga kondisi ketersediaan pakannya. Faktor lingkungan sebagai pembatas bagi ternak di daerah ini adalah ketersediaan pasokan pakan yang tersedia bagi ternak, dimana di Propinsi Sulawesi Selatan mempunyai bulan kering antara 6 - 9 bulan dalam 1 tahun dan kondisi tanah yang relatip tipis tanah humusnya. Sedangkan di Kabupaten Samosir walaupun curah hujan relatip tinggi, tetapi kondisi tanah hampir sama dengan kondisi di Sulawesi Selatan yaitu ketebalan tanah humus relatip tipis dan berbatu-batu. I II K. Muara K. Kacang K. Jawarandu K. Marica K. Samosir K. Benggala IV Gambar 6 III Plot penyebaran kelompok kambing berdasarkan ukuran-ukuran fenotipik pada 6 sub populasi kambing lokal. 36 Nilai Campuran Fenotipik antar Kelompok Tabel 7 menyajikan persentase nilai kesamaan dan campuran kelompok sub populasi kambing. Kemungkinan besar proporsi nilai campuran yang mempengaruhi kesamaan suatu rumpun lain didasarkan atas kesamaan ukuran fenotipik (Sumantri et al. 2007). Tabel 7 Persentase nilai kesamaan dan campuran 6 sub populasi kambing lokal Sub populasi Benggala Jawarandu Kacang Marica Muara Samosir Benggala 88.57 2.17 0.00 0.00 0.00 5.00 Jawa Kacang randu 0.00 0.00 91.30 0.00 0.00 99.28 10.00 0.00 0.00 0.00 2.50 0.00 Marica Muara Samosir Total 5.71 6.52 0.73 83.33 0.00 2.50 0.00 0.00 0.00 0.00 93.33 7.50 5.71 0.00 0.00 6.67 6.67 82.50 100.00 100.00 100.00 100.00 100.00 100.00 Hasil analisis nilai kesamaan dan campuran kelompok Kambing Samosir mempunyai nilai kesamaan paling rendah 82.50% karena dipengaruhi nilai campuran dengan Kambing Muara 7.5%, Kambing Benggala 5.71%, Jawarandu 2.5% dan Kambing Marica 2.5%. Kelompok Kambing Marica mempunyai nilai kesamaan 83.33%, karena dipengaruhi nilai campuran Kambing Jawarandu 10% dan Kambing Samosir 6.67%. Kelompok Kambing Benggala mempunyai nilai kesamaan 88.57%, karena dipengaruhi oleh nilai campuran Kambing Marica dan Kambing Samosir dengan nilai masing-masing sub populasi kambing sebesar 5.71%. Kelompok Kambing Jawarandu mempunyai nilai kesamaan 91.30% karena dipengaruhi oleh nilai campuran Kambing Marica 6.52% dan Kambing Benggala 2.17%. Kelompok Kambing Muara mempunyai nilai kesamaan paling tinggi 93.33%, karena hanya dipengaruhi oleh nilai campuran Kambing Samosir 6.67%. Faktor genetik dan lingkungan mempunyai hubungan yang erat, dan untuk mengekpresikan kapasitas genetik individu secara sempurna diperlukan kondisi lingkungan yang ideal. 37 Penentuan Jarak Genetik dan Pohon Fenogram Nilai matrik jarak genetik antar kelompok 6 sub populasi kambing pada sub populasi yang berbeda dapat dilihat pada Tabel 8, digunakan untuk membuat konstruksi pohon fenogram (Gambar 7). Pohon fenogram tersebut menggambarkan jarak genetik keseluruhan kelompok. Hasi analisis pada Tabel 8 memperlihatkan bahwa nilai terkecil didapat pada jarak antara sub populasi Kambing Samosir dengan Kambing Marica yaitu sebesar 11.207. Nilai terbesar diperoleh dari Kambing Muara - Benggala (255.110), kemudian disusul oleh Kambing Muara - Marica (187.865), serta Kambing Kacang - Benggala (139.942) dan Kambing Muara - Kacang (133.471). Nilai matrik jarak genetik yang relatip besar didapatkan dari jarak genetik antara Kambing Muara - semua kelompok, dan jarak genetik Kambing Kacang - Benggala. Tabel 8 Jarak genetik berdasarkan ukuran tubuh antar 6 sub populasi kambing Sub populasi Benggala Jawa randu Kacang Marica Muara Samosir Benggala 0 97.977 139.942 15.339 255.110 22.888 Jawa randu Kacang Marica Muara Samosir 0 66.599 51.890 64.170 57.964 0 98.214 133.471 93.086 0 187.865 11.207 0 162.586 0 Jarak genetik adalah tingkat perbedaan gen antar populasi atau spesies yang diukur oleh beberapa kuantitas numerik (Nei 1987). Metode yang lebih murah dan sederhana yang dapat dilakukan dengan penentuan pola perbedaan sifat fenotipik yang dapat ditemui dalam setiap individu ternak (Hartl 1988). Penggunaan ukuran-ukuran tubuh sebagai penduga terhadap jarak genetik dan peubah pembeda dari 5 kelompok Kambing Andalusia dengan menggunakan analisis diskriminan telah dilaporkan Herera et al. (1996) dan Traore et al. (2008) pada kambing lokal di Burkina Faso serta Suparyanto et al. (1999) dan Sumantri et al. (2007) pada domba di Indonesia. Secara umum hasil analisis matrik jarak berdasarkan data ukuran-ukuran tubuh dengan program MEGA menunjukkan bahwa setiap sub populasi masingmasing menunjukkan indek jarak > 60 % antara satu sub populasi terhadap sub populasi kambing lainnya. Ini menunjukkan bahwa jarak karakteristik morfometrik antara setiap sub populasi berbeda nyata terhadap sub populasi kambing lokal lainnya. Pohon fenogram dari 6 sub populasi kambing lokal menurut ukuran fenotipik dapat dilihat pada Gambar 7. 38 1.6738 0.5013 M 1.6738 3.2989 S 2.1752 B 4.9284 K 4.0053 0.5456 4.0053 0.9232 5 J 4 3 R 2 1 0 K=Kacang; B=Benggala; S=Samosir; J=Jawa randu; M=Marica; R=Muara. Gambar 7 Pohon fenogram dari 6 sub populasi kambing lokal berdasarkan ukuran fenotipik. Matrik jarak menunjukkan bahwa sub populasi Kambing Samosir - Marica dan Kambing Marica - Benggala memiliki ukuran jarak yang relatip dekat yaitu berturut-turut 11.207 dan 15.339, jika dibandingkan dengan jarak berdasarkan ukuran fenotipik antara sub populasi Kambing Samosir - Benggala (22.888) dan Kambing Marica - Jawarandu (51.890). Sub populasi Kambing Muara di Kabupaten Tapanuli Utara, Propinsi Sumatera Utara berdasarkan analisis fenogram terpisah dari kelompok Kambing Benggala, Kacang, Marica dan Kambing Samosir. Jarak sub populasi Kambing Muara menunjukkan cabang kaitan tidak langsung antara Kambing Marica, Samosir, Benggala dan Kambing Kacang. Hasil pohon fenogram sesuai dengan peta penyebaran yang menunjukkan adanya enam kelompok sub populasi terpisah, yaitu; (1) Kambing Muara, (2) Kambing Jawarandu, (3) Kambing Kacang, (4) Kambing Benggala, (5) Kambing Marica dan (6) Kambing Samosir. Hasil peta penyebaran berdasarkan ukuran tubuh dan pohon fenogram memberikan gambaran kelompok ternak kambing sebaiknya kita silangkan. Bourdon (2000) menjelaskan persilangan antar individu yang mempunyai jarak lebih jauh akan memberikan performa yang lebih baik dari rataan para tetuanya, karena adanya peningkatan heterosigositas dan kombinasi gen. 39 Peubah Pembeda Rumpun Kambing Hasil analisis struktur kanonikal disajikan pada Tabel 9 menunjukkan bahwa ukuran fenotipik kambing yang memberikan pengaruh kuat terhadap peubah pembeda kelompok sub populasi kambing adalah lingkar kanon (0.7Kan1), lebar telinga (0.5-Kan2), lebar pinggul (0.5-Kan2), lebar ekor (0.7-Kan3), panjang badan (0.7-Kan3), tinggi tengkorak (0.5 Kan-3), lebar tengkorak (0.5Kan3), tinggi pundak (0.5-Kan3), bobot badan (0.5-Kan3), lingkar dada (0.5Kan3), lebar dada (0.5-Kan4) dan dalam dada (0.5-Kan4). Dari 19 variabel pengukuran yang diamati terdapat 11 variabel ukuran tubuh yang mempunyai nilai kanonikal ≥ 0.5 (data dalam tabel dibulatkan menjadi satu desimal) sehingga lingkar kanon, panjang badan, lebar tengkorak, tinggi tengkorak, lebar ekor dapat digunakan sebagai peubah pembeda kelompok kambing. Tabel 9 Struktur kanonikal kelompok kambing dari 6 sub populasi berdasarkan ukuran fenotipik Variabel Bobot badan Panjang badan Lingkar dada Lebar dada Tinggi Pundak Dalam dada Lingkar pinggul Lebar pinggul Tinggi Pinggul Dalam pinggul Lingkar kanon Panjang tanduk Panjang telinga Lebar telinga Panjang tengkorak Lebar tengkorak Tinggi tengkorak Panjang ekor Lebar ekor Kan-1 0.04 0.02 -0.03 -0.18 0.22 0.33 0.00 -0.45 0.30 0.33 0.67 -0.09 0.40 -0.31 0.24 0.28 0.29 0.17 0.37 Kan-2 0.26 0.03 0.21 0.03 0.11 0.30 0.18 0.46 0.25 0.00 0.67 0.24 0.18 0.46 -0.32 0.11 -0.06 0.04 -0.35 Kan-3 0.48 0.67 0.46 0.07 0.50 0.05 0.37 0.44 0.34 0.07 0.20 0.24 0.26 0.37 0.18 0.52 0.53 0.09 0.71 Kan-4 -0.21 -0.17 -0.28 0.52 -0.29 0.46 -0.23 0.24 -0.05 0.09 -0.11 -0.09 -0.37 0.36 -0.39 -0.14 0.01 -0.07 -0.31 Menurut Traore et al. (2008) analisis variasi kanonikal digunakan untuk mendapatkan kombinasi karakter yang membedakan secara keseluruhan dan dapat digunakan untuk menggambarkan plot skor guna membandingkan di dalam dan diantara variabilitas populasi (kelompok kambing) pada dimensi yang kecil. Semakin rendah angka yang diperoleh dari hasil analisis struktur kanonik, semakin tidak dapat digunakan sebagai peubah pembeda kelompok kambing 40 Pola Warna Tubuh Pola warna dibedakan atas dua kelompok yaitu kelompok warna dominan dan kelompok warna belang. Warna dominan adalah kelompok warna yang paling banyak persentase warna tubuh atau paling tidak diperkirakan diatas atau sama dengan 60%, sedangkan yang dimaksud dengan warna belang adalah warna tubuh yang selain warna dominan. Persentase pola warna tubuh dominan dan warna belang tubuh pada 6 sub populasi kambing lokal Indonesia dapat dilihat pada Tabel 10. Tabel 10 Persentase pola warna tubuh dominan dan warna belang pada 6 sub populasi kambing lokal Pola Warna tubuh Warna dominan Putih Coklat muda Coklat kemerahan Coklat Coklat tua Abu-abu Hitam Kacang n=217 Variasi 28 15 5 6 16 2 28 Samosir n=42 Putih 86 2 0 7 7 0 5 Benggala n=96 Hitam 10 10 10 18 0 0 60 Muara n=34 Variasi 53 0 37 0 0 3 7 Marica Jawarandu n=60 n=94 Variasi Variasi 17 6 30 4 10 0 37 57 0 13 0 2 7 17 Warna belang Putih Coklat muda Coklat kemerahan Coklat Coklat tua Abu-abu Hitam Variasi 43 19 22 8 4 0 4 Variasi 12 2 0 26 10 2 48 Variasi 18 20 15 0 0 0 47 Variasi 37 3 0 37 0 0 23 Hitam 27 3 0 3 0 0 67 Variasi 34 0 2 17 2 2 43 Warna tubuh kambing yang diamati antara lain warna putih, coklat muda, coklat kemerahan (merah bata), coklat, coklat tua kehitaman, abu-abu, dan warna hitam. Warna tubuh dan pola warna kambing sangat bervariasi, ada yang mempunyai pola warna yang dominan tunggal dan ada juga yang sangat beragam (pola warna belang dan totol-totol). Kambing Benggala dan Samosir mempunyai warna tubuh tunggal dominan yang khas yaitu Kambing Benggala didominasi warna tunggal hitam dan Kambing Samosir didominasi warna putih. Kambing Muara walaupun kebanyakan warna putih tetapi dikombinasi belang atau totol-totol berwarna hitam, coklat kemerahan (merah bata) dan warna abu-abu. Kambing Marica bervariasi antara lain warna coklat, coklat 41 muda dan warna putih dengan kombinasi warna belang hitam, putih dan warna coklat muda. Sedangkan Kambing Kacang dan Jawarandu menunjukkan pola warna tubuhnya yang sangat bervariasi, sehingga hampir semua warna yang diamati terdapat pada kedua sub populasi kambing tersebut. Pola warna tubuh Kambing Kacang secara umum sangat bervariasi antara lain warna putih, hitam, coklat dan warna abu-abu keputihan. Dari hasil pengamatan warna tubuh dominan putih dan hitam (masing-masing 28%) yang paling tinggi, kemudian diikuti oleh warna coklat tua (16%), coklat muda (15%), coklat (7%), coklat kemerahan (merah bata) (5%) dan abu-abu (2%). Pola warna belang tubuh juga yang paling tinggi adalah warna putih (43%), kemudian coklat kemerahan (22%) dan warna coklat muda (19%). Kemudian diikuti warna coklat (8%), coklat tua (4%) dan belang warna hitam (4%). Contoh warna dan pola warna dominan 6 sub populasi kambing lokal Indonesia dapat dilihat pada Gambar 8. Warna yang dominan Kambing Samosir adalah warna putih (86%), kemudian warna coklat dan coklat tua, hitam dan coklat muda. Pola warna belang Kambing Samosir paling tinggi adalah warna hitam (48%) dan coklat (26%), kemudian diikuti oleh warna putih (12%), coklat tua (10%), hitam (2,4%) dan coklat muda (2%). Kambing Benggala mempunyai pola warna dominan hitam (60%), kemudian warna coklat (18%), coklat kemerahan, coklat muda dan warna putih (masing-masing 10%). Pola warna belang Kambing Benggala antara lain warna hitam (47%), diikuti warna coklat muda (20%), putih (18%) dan warna coklat kemerahan(15%). Kambing Muara mempunyai warna tubuh dominan bervariasi kebanyakan warna putih yang paling tinggi (53%) dan coklat (37%), kemudian warna hitam (7%) dan abu-abu (3%). Pola warna belang Kambing Muara antara lain putih dan coklat (masing-masing 37%), kemudian hitam (23%) dan coklat muda (3%). Pada Kambing Muara dijumpai pola warna dominan putih dengan warna totol-totol (spotted) warna hitam. Kambing Marica mempunyai warna tubuh dominan bervariasi, mulai warna coklat yang paling tinggi (37%) dan coklat muda (30%), kemudian diikuti warna putih (17%), coklat kemerahan (merah bata) (10%) dan warna hitam (7%). Pola warna belang Kambing Marica antara lain hitam (67%) dan putih (27%). Kemudian diikuti warna coklat dan coklat kemerahan (masing-masing 3%). 42 Kambing Kacang Kambing Benggala Kambing Samosir Kambing Marica Kambing Jawarandu Kambing Muara Gambar 8 Pola warna dominan dan belang pada 6 sub populasi kambing lokal Kambing Jawarandu mempunyai warna dominan sangat bervariasi, kebanayakan warna coklat (57%), kemudian diikuti oleh warna hitam (17%), coklat tua (13%), putih (6%), coklat muda (4%)dan abu-abu (2%). Pola warna belang Kambing Jawarandu antara lain hitam (43%), putih (34%), kemudian diikuti oleh warna coklat (17%), coklat kemerahan, coklat tua dan abu-abu 43 (masing-masing 2 %). Dengan beragamnya warna dominan dan warna belang tubuh kambing yang diamati semakin terbuka peluang untuk melakukan seleksi pembentukan warna-warna tertentu yang khas jika dibutuhkan. Simpulan Keenam sub populasi kambing yang diamati menunjukkan bahwa setiap sub populasi berbeda antara satu kelompok dengan kelompok lainnya, sehingga dapat dibedakan menjadi 6 kelompok kambing yaitu Kambing Muara, Kambing Kacang, Kambing Jawarandu, Kambing Marica, Kambing Samosir dan Kambing Benggala. Kambing Muara memiliki bobot badan dan ukuran tubuh lebih besar jika dibandingkan dengan Kambing Jawarandu, Benggala, Kacang, Samosir dan Kambing Marica. Variabel pembeda untuk karakterisasi dan seleksi berdasarkan morfometrik pada kambing lokal adalah parameter bobot badan hidup, panjang badan, tinggi pundak, lebar dada, dalam dada, lingkar dada, lebar tengkorak, tinggi tengkorak, lebar ekor, panjang ekor, lebar telinga, panjang telinga dan lingkar kanon. Terdapat warna dominan yang khas yaitu warna hitam pada Kambing Benggala dan warna putih pada Kambing Samosir, sedangkan Kambing Kacang, Jawarandu, Muara dan Marica menunjukkan pola warna dominan sangat bervariasi. KARAKTERISASI MOLEKULER ENAM SUB POPULASI KAMBING LOKAL INDONESIA BERDASARKAN ANALISIS SEKUEN DAERAH D-LOOP DNA MITOKONDRIA Pendahuluan Analisis DNA mitokondria (mtDNA) merupakan salah satu metode yang banyak digunakan untuk mempelajari asal-usul ternak domestikasi (MacHugh & Bradley 2001). Pada mamalia DNA mitokondria hanya diturunkan lewat jalur induk (maternal) tanpa rekombinasi. Sekuen nukleotida genom mitokondria telah digunakan untuk mempelajari asal-usul sapi (Troy et al. 2001), babi (Giuffra et al. 2000), domba (Hiendleder et al. 2002), kuda (Villa et al. 2001), anjing (Savolainen et al. 2002), keledai (Beja-Pereira et al. 2004) dan kambing (Joshi et al. 2004; Luikart et al. 2001; Mannen et al. 2001; Sultana et al. 2003; Chen et al. 2005; Naderi et al. 2007; Royo et al. 2009; Zhao et al. 2011). Kambing domestik dapat dikelompokkan menjadi 4 garis kelompok keturunan utama atau Lineage (Joshi et al. 2004). Lineage A merupakan kelompok kambing domestikasi yang paling beragam dan luas penyebarannya di dunia. Lineage B menyebar di daerah Asia Timur dan Asia Selatan termasuk Cina, Mongolia, Afrika Selatan, Afrika Utara, Laos, Malaysia, Pakistan dan India. Lineage C menyebar di sekitar Mongolia, Swiss, Slovenia, Pakistan dan India. Lineage D di daerah Pakistan dan kambing lokal di India. Selain itu Naderi et al. (2007) mengelompokkan kambing menjadi enam haplogroup yaitu menambahkan haplogroup F dan G dengan 4 haplogroup yang dilaporkan oleh Joshi et al. (2004). Lineage F menyebar di daerah Sicilia dan lineage G menyebar di daerah Timur Tengah dan Afrika Utara. Kombinasi pengelompokan molekuler dengan temuan arkeologi menunjukkan bahwa kambing domestikasi mempunyai asal usul beberapa garis keturunan ibu (maternal origins) (Zeeder & Hesse 2000). Dengan penambahan sampel terutama di daerah yang mungkin merupakan asal garis keturunan, maka analisis mtDNA akan mendukung pemahaman pengelompokan kambing lokal di berbagai wilayah (MacHugh & Bradley 2001; Chen et al. 2005). 45 Bahan dan Metode Sampel Darah Kambing Penelitian Pengambilan sampel darah sekitar 2 ml dari setiap ekor kambing penelitian dilakukan pada bagian vena jugularis dengan menggunakan jarum venoject yang disambungkan ke tabung vakum dengan EDTA 5 ml. Semua sampel darah selama di lapangan dengan etanol absolut 2x volume darah dikocok hingga homogen. Ekstraksi DNA Ekstraksi DNA dilakukan dengan memodifikasi metode Sambrook et al. (1989) menggunakan bufer lisis sel 350 µl 1xSTE, dan 40 µl 10% SDS, 20 µl 5 mg/ml proteinase-K. DNA dimurnikan dengan metode fenol-kloroform, yaitu dengan menambahkan 40 µl 5 M NaCl dan 400 µl fenol dan kloroform: iso amil alkohol (24:1). DNA diendapkan dengan menambahkan 40 µl 5 M NaCl dan 800 µl etanol absolute. Endapan dicuci dengan menambahkan 400 µl etanol 70%, Selanjutnya sisa etanol dibuang dan diuapkan dengan menggunakan pompa vakum. DNA kemudian disuspensikan dengan 80 µl 80% bufer TE. Amplifikasi DNA Sebanyak 543 sampel DNA yang telah diekstraksi dipilih sebanyak 60 sampel yang masing-masing 10 sampel DNA per sub populasi. Amplifikasi ruas D-loop genom 5’GCGTACGCAAT mitokondria menggunakan CTTACGATCA-3’ 5’ATGCAGTTAAGTCCAGCTAC-3’. primer dan AF23 AF22 (forward) (reverse) Primer AF23 menempel pada basa ke 14979-14999 dan AF22 menempel pada basa ke 16379-16398 dari mtDNA Capra hircus (GenBank no akses AF 533441). Pasangan primer AF23 dan AF22 mengapit ruas tengah hingga akhir Cyt b, tRNA Pro, tRNA Thr dan juga bagian awal hingga tengah daerah D-loop dengan panjang 1420 pb (Gambar 9). Komposisi reaksi PCR dalam volume 25µl adalah sampel DNA 2µl (10100ng), RBC Bioscience taq polymerasi 1.25 unit beserta sistim bufernya, dNTP mix 0.4 nmol, MgCl 2 0.2 mM, primer AF22 dan AF23 masing-masing 1 nmol. Kondisi PCR yang digunakan untuk proses amplifikasi adalah tahap denaturasi awal pada suhu 94oC selama 3 menit, tahap denaturasi pada suhu 94oC selama 46 45 detik, tahap penempelan primer (annealing) pada suhu 58oC selama 50 menit, dan tahap polimerasi (extension) pada suhu 72oC selama 1 menit yang diulang selama 30 siklus, kemudian reaksi PCR diakhiri dengan polimerasi akhir pada suhu 72oC selama 5 menit. Visualisasi produk PCR dilakukan menggunakan teknik elektroforesis gel poliakrilamid (PAGE) 6% dalam buffer 1xTBE (Tris-HCl 10 Mm, asam borat 1 M, EDTA 0.1 mM). Elektroforesis dijalankan pada kondisi 200 mV selama 50 menit. Proses dilanjutkan dengan pewarnaan sensitif perak (Tegelstrom 1986) yang dimodifikasi. Perunutan DNA Produk amplifikasi yang menunjukkan pita tunggal (sekitar 1400 pb) dimurnikan dan dijadikan cetakan dalam reaksi PCR untuk perunutan nukleotida. Masing-masing kelompok kambing dipilih 5 sampel yang saling berjauhan lokalitasnya. Primer yang digunakan dalam proses penentuan runutan nukleotida sama dengan primer yang digunakan untuk amplifikasi. Reaksi PCR tersebut dilakukan dengan menggunakan metode dideoxi terminator dengan dNTP berlabel (big dye terminator). Perunutan nukleotida menggunakan mesin ABI Prism 3700-Avant Genetic Analyzer di PT Charoen Pockphan dan PT Genetika Sains Indonesia Jakarta. Analisis Data Runutan nukleotida yang diperoleh kemudian diedit secara manual dengan bantuan program Bio Edit versi 6.0.7. Urutan DNA yang telah diedit disejajarkan dengan beberapa runutan DNA dari kelompok Capra hircus yang dipublikasikan dalam GenBank (http://ncbi.nlm.nih.gov). Proses pensejajaran menggunakan ClustalW versi 8.1 yang ada dalam program MEGA 4 (Tamura et al. 2007) yang kemudian diedit lagi secara manual terutama untuk ruas-ruas DNA berulang (Lampiran 4). Analisis yang dilakukan meliputi penghitungan komposisi nukleotida, laju subsitusi, jarak genetik menggunakan program MEGA versi 4.0 (Tamura et al. 2007). Perhitungan nilai jarak genetik dilakukan berdasarkan model subsitusi Kimura 2 parameter (K2P). Rekonstruksi pohon filogeni dilakukan berdasarkan semua nukleotida yang bersifat parsimoni menggunakan metode Neigbour 47 Joining (NJ) dengan uji bootstrap 1000 kali dengan program NETWORK versi 4.6 (Fluxus Technology Ltd. 2010). Hasil dan Pembahasan Polimorfisme Segmen daerah D-loop DNA Mitokondria Panjang ruas D-loop yang saling sejajar antar sampel adalah 879 pb. Dari situs-situs ruas D-loop tersebut ditemukan 50 situs polimorfik yang terdiri dari 21 mutasi insersi dan 29 transversi (Gambar 10). Berdasarkan 50 situs nukleotida yang bersifat polimorfik ke-6 sub populasi kambing lokal bisa dibagi menjadi 19 haplotip. Dari 19 jumlah haplotip, masing-masing sub populasi kambing lokal mempunyai jumlah haplotip khas yang bervariasi antara 2-4 haplotip. Variasi haplotip yang khas tersebut dapat digunakan sebagai penciri genetik pada setiap sub populasi kambing lokal Indonesia (Tabel 11). Tabel 11 Jumlah haplotip berdasarkan runutan nukleotida D-loop mtDNA setiap sub populasi kambing lokal Indonesia Kambing lokal n sample n haplotipe Haplotipe Marica (M) 5 3 1, 2, 3 Kacang (K) 5 4 4, 5, 6, 7 Samosir (S) 5 2 8, 9 Benggala (B) 5 3 10, 11, 12 Jawarandu (J) 5 4 13, 14, 15, 16 Muara (R) 5 3 17, 18, 19 Panjang D-loop (1.212pb) t-RNA CYTB Thr Pro 14.153 14.981 15.292 15.296 15.431 15.365 1 AF23 15.431 16.309 541 D-loop Kambing lokal 1.420 16.643 pb (879 pb) AF22 Nukleotida DNA mt kambing lokal (1.420 pb) Gambar 9 Struktur genom mitokondria yang diapit oleh primer AF23 dan AF22. Nomor mengacu pada Capra hircus (GenBank no. akses AF 533441) 48 2333344444 3556800014 5346058977 C.hircus_NC_005044 T---CAA-AA Marica1 .---...-.G Marica2 .---...-.G Marica3 .---...-.G Marica4 .---...-.G Marica5 .---...-.G Kacang1 .---T.G-GG Kacang2 .---T.G-GG Kacang3 .---T.G-GG Kacang4 .---T.G-GG Kacang5 .---T.G-GG Samosir1 .---...-.G Samosir2 .---...-.G Samosir3 .---...-.G Samosir4 .---...-.G Samosir5 .---...-.G Benggala1 .---.C.-.. Benggala2 .---.C.A.. Benggala3 .---...-.. Benggala4 .---.C.A.. Benggala5 .---.C.-.. Jawarandu1 .---...-.. Jawarandu2 .--T...-.. Jawarandu3 .---...-.. Jawarandu4 .---...-.. Jawarandu5 .---...-.. Muara1 .AC-...-.. Muara2 .AC-...-.. Muara3 .AC-...-.. Muara4 CAC-...-.. Muara5 CAC-...-.. 4445555555 6890223444 7744459389 TAACCCCCTC .G..T-..CT .G..T-..CT .G..T-..CT .G..T-..CT .G..T-..CT CGGTT.TT.. CGGTT.TT.. CGGTT.TT.. CGGTT.TT.. CGGTT.TT.. .G..T-.... .G..T-.... .G..T-.... .G..T-.... .G..T-.... .......... .......... .......... .......... .......... .......... .G........ .G........ .G........ .G........ .......... .......... .......... .......... .......... 5555555555 5556788888 0166204589 CTAGTTCCTA TCGA..TT.. TCGA..TT.. TCGA..TT.. TCGA..TT.. TCGA..TT.. ..GA..T... ..GA..T.CC ..GA..T... ..GA..T... ..GA..T... ....GCT... ......T... ......T... ......T... ......T... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 566666666 901144456 311403977 AATCCGGCG ..CT..... ..CT..... ..CT..... ..CT..... ..CT..... GG..T..T. ......... G......T. G......T. GG.....T. ......... ......... ......... ......... ......... G........ G........ G........ G........ G........ ...T.A... ......... .....A... .....A... .....AA.C ......... ......... ......... ......... ......... 6777777788 8014466700 8891817936 TGATCCCCCT .........C .........C .........C ...C.....C .........C .......... .......... .......... .......... .......... A...TAGG.. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .CG....... .......... ........G. .......... .......... .......... 888888 224566 390656 TTCCCT ...... ...... ...A.. ....TC ...... ...... ...... ...... ...... ...... .CA... ...... ...... ...... ...... ...... ...... ...... ...... ...... ...... ...... ...... ...... ...... C..... C..... C..... C..... C..... Gambar 10 Polimorfisme runutan nukleotida daerah D-loop DNA mitokondria pada 6 sub populasi kambing lokal mengacu pada Capra hircus (GenBank no. akses AF 533441) (tiga baris pertama dibaca secara vertikal merupakan posisi nukleotida) Setiap kelompok kambing juga ditemukan perbedaan antara satu kelompok sub populasi kambing lokal dengan kelompok yang lain, sehingga runutan nukleotida yang khas dapat digunakan sebagai penciri dari setiap sub populasi kambing lokal tersebut (Tabel 12). Perubahan (mutasi) susunan nukleotida berupa subtitusi ditemukan pada situs nukleotida Kambing Kacang, Marica, dan Kambing Muara, sedangkan pada Kambing Benggala, Jawarandu dan Kambing Muara dijumpai mutasi berupa subtitusi dan insersi. Mutasi insersi nukleotida yang ditemukan pada 3 sub populasi yaitu insersi nukleotida AC pada 49 situs ke 353, 354 pada Kambing Muara, insersi nukleotida T pada situs ke 366 pada Kambing Jawarandu dan insersi A pada situs ke 409 pada Kambing Benggala, sedangkan mutasi nukleotida pada 3 sub polpulasi lainnya merupakan mutasi substitusi. Tabel 12 Mutasi nukleotida sebagai penciri kelompok kambing lokal Indonesia dibandingkan dengan Capra hircus (GenBank no. akses AF 533441) Sub populasi Subtitusi n Situs ke- Insersi n Situs ke- T-C 5 - - - T-C 1 548, 551, 611, 806 741 C-T 5 C-A 1 549, 585, 614 856 CT-TC 1 865-866 C-T 5 - - - A-G 5 380, 539, 543, 640 408, 601 T-C 5 467, 588 A-C 1 589 G-T 4 667 T-G 1 572 - - - Marica Kacang Samosir Benggala Jawarandu Muara T-A 1 688, 840 C-T 1 748 C-A 1 761 C-G 1 767, 779 C-G 1 857 T-C 1 580, 829 C-A 1 918 A-C 4 405 A 2 409 C-T 4 721 T 1 366 G-C 1 667, 708 G-A 1 643 A-G 1 719 C-G 1 803 AC 5 353-354 T-C 5 235, 823 Keterangan: n=jumlah situs sekuen 50 Keragaman Runutan Nukleotida Urutan frekuensi nukleotida paling tinggi terdapat pada nukleotida A(33.7), kemudian T (27.2%), C (26.4) dan G (12.6) secara berurutan. Perbandingan rataan frekuensi A dan T (60.9%) lebih tinggi dibandingkan C dan G (39.8%). Perbedaan susunan basa nukleotida paling rendah ditemukan antara Kambing Kacang dengan C. hircus AF 533441 (0.5%), dan tertinggi dijumpai antara Kambing Kacang dengan Muara (10.30%). Rataan keragaman susunan nukleotida secara keseluruhan adalah 0.014±0.002. Keragaman susunan nukleotida antar individu dalam kelompok kambing penelitian yaitu; Marica (0.001), Kacang (0.005), Samosir (0.008), Jawarandu (0.004), Muara (0.001) dan Kambing Benggala tidak terdapat perbedaan antar individu dalam kelompok (0.000). Perbedaan keragaman susunan nukleotida antar kelompok kambing lokal berkisar antara 0.004 – 0.103. Keragaman susunan nukleotida paling tinggi adalah antara Kambing Kacang dengan Muara (0.1030) dan paling rendah dijumpai antara Kambing Samosir dengan Marica (0.004) dan nilai yang sama dijumpai antara Kambing Jawarandu dan Kambing Muara (Tabel 13). Tabel 13 Keragaman nukleotida D-loop mtDNA pada 6 kambing lokal Indonesia C. hircus Genotip Capra hircus* Marica Kacang Samosir Benggala J.randu Muara 0 Marica (M) 0.022 0 Kacang (K) 0.005 0.041 0 Samosir (S) 0.025 0.004 0.043 0 Benggala (B) 0.049 0.008 0.072 0.013 0 Jawarandu (J) 0.061 0.011 0.089 0.016 0.005 0 Muara (R) 0.073 0.016 *GenBank no. akses AF 533441 0.103 0.021 0.004 0.004 0 Jarak Genetik Kambing Penelitian dengan Kambing Lainnya Jarak genetik digunakan untuk melihat kedekatan hubungan genetik ternak yang dipelihara di daerah yang berbeda. Selanjutnya informasi jarak genetik dapat digunakan sebagai petunjuk awal dari stuktur populasi dan diferensiasi suatu rumpun di dalam membuat keputusan program konservasi (Ponzoni 1997). Jarak genetik antar individu dalam kelompok kambing penelitian antara lain; nilai jarak genetik Kambing Samosir 0.004, Kacang dan Jawarandu 0.003, 51 Marica 0.002, Muara 0.001 dan Kambing Benggala tidak ada perbedaan (0.000). Nilai jarak genetik antar individu dan sub populasi ini menunjukkan sampel Kambing Benggala relatif homogen walaupun diambil dari tempat berbeda kabupaten. Nilai jarak genetik antar kelompok kambing lokal berkisar 0.004-0.044 dan rataan jarak genetik keseluruhan adalah 0.014. Jarak genetik tertinggi antara kambing lokal dengan pembanding Capra hircus (AF 533441) ditemukan pada Kambing Muara (0.044) dan terendah dengan Kambing Kacang (0.028). Nilai jarak genetik antar kelompok kambing lokal terkecil antara Kambing Muara dan Benggala (0.004), sedangkan tertinggi merupakan jarak genetik antara Kambing Kacang dan Marica (0.004) (Tabel 16). Jarak genetik Kambing Marica paling dekat dengan Kambing Samosir (0.015) dan paling jauh dengan Kambing Kacang (0.023). Jarak genetik Kambing Kacang paling dekat hubungannya dengan Kambing Samosir (0.017) dan paling jauh hubungannya dengan Kambing Jawarandu dan Muara (0.021). Jarak genetik Kambing Samosir paling dekat hubungannya dengan Kambing Jawarandu (0.008) dan paling jauh hubungannya dengan Kambing Muara dan Benggala (0.009). Jarak genetik Kambing Benggala lebih dekat hubungannya dengan Kambing Muara (0.004) dibandingkan dengan Kambing Jawarandu (0.005). Jarak Kambing Jawarandu dengan Kambing Muara adalah 0.005 (Tabel 14). Hal ini menunjukkan bahwa Kambing Jawarandu masih dekat hubungan kekerabatan dengan Kambing Muara. Pohon filogeni yang dibentuk berdasarkan metode 2 parameter Kimura dalam uji bootstrap 1000 kali pengulangan, diperoleh enam klaster kambing yaitu masing-masing sub populasi membentuk klaster tersendiri yaitu klaster Kacang, Marica, Samosir, Jawarandu, Muara dan Benggala. Kambing Kacang merupakan klaster yang relatif dekat dengan Capra hircus dengan nilai uji bootstrap (65%). Kambing Kacang dikenal sebagai kambing asli Indonesia yang tersebar secara luas hampir di seluruh kepulauan yang ada penduduknya. Kambing Jawarandu, Muara dan Benggala menunjukkan terjadi mutasi perubahan susunan basa nukleotida dalam bentuk subtitusi dan insersi (Tabel 14). Perubahan susunan nukleotida dalam bentuk insersi pada ketiga kambing ini diduga karena merupakan hasil persilangan pejantan kambing dari luar (outgroup) dengan induk Kambing Kacang (hibridisasi) yang telah 52 beradaptasi (berevolusi) dengan kondisi agro-ekosistim lokal dimana kambing tersebut berada. Tabel 14 Jarak genetik berdasarkan runutan nukleotida pada 6 sub populasi kambing lokal Indonesia Sub populasi Capra hircus* C. hircus Marica Kacang Samosir Benggala J.randu Muara 0 Marica (M) 0.03 0 Kacang (K) 0.028 0.023 0 Samosir (S) 0.041 0.015 0.017 0 Benggala (B) 0.041 0.019 0.02 0.009 0 Jawarandu (J) 0.042 0.018 0.021 0.008 0.005 0 Muara (R) 0.044 0.019 *GenBank no. akses AF 533441 0.021 0.009 0.004 0.005 0 Mutasi subtitusi ditemukan pada Kambing Kacang, Marica dan Samosir diduga merupakan tanda adanya proses adaptasi dari Kambing Kacang (sebagai kambing asli) dengan kondisi lokasi baru yang berbeda dengan kondisi di Sumatera, Jawa dan Bali, dimana perubahan susunan basa nukleotida terjadi dalam bentuk substitusi sebagai akibat proses adaptasi terhadap kondisi lingkungan yang sumber pakannya terbatas dan diduga akibat adanya seleksi yang berhubungan dengan tujuan produksi yang diinginkan oleh peternak. Seperti Kambing Samosir, tujuan utama pemeliharaan kambing ditujukan untuk menghasilkan kambing yang berwarna putih, karena nilai ekonomi (harga jual) yang tinggi. Pengguna atau konsumen menggunakan kambing berwarna putih untuk keperluan acara ritual bagi penganut agama/aliran kepercayaan Parmalim di daerah Gunung Pusuk Buhit dan sekitar Kabupaten Samosir. Perubahan mutasi nukleotida pada Kambing Marica diduga disebabkan proses adaptasi dengan kondisi iklim yang berbeda dan ketersediaan bahan pakan terutama pada saat musim kemarau yang rata-rata diatas 6-9 bulan per tahun membuat ketersediaan rumput sangat terbatas. Pada musim pertengahan dan akhir musim kemarau umumnya rumput sudah layu dan kering diakibatkan musim kering yang berkepanjangan seperti pada umumnya di daerah kepulauan Indonesia Bagian Timur, sehingga ketersediaan rumput sangat terbatas. Kemungkinan dalam jangka waktu yang lama Kambing Marica mengalami proses adaptasi dengan kondisi setempat, maka terjadilah proses mutasi 53 subtitusi nukleotida yang secara fenotip Kambing Marica mempunyai performans tubuh yang lebih kecil jika dibandingkan dengan Kambing Kacang. Jika digabungkan dengan sekuen nukleotida daerah D-loop DNA mitokondria kambing dari luar negeri yang terdapat pada GenBank, kelompok kambing lokal yang diamati mempunyai jarak genetik yang jauh dari Capra hircus (AF 533441) karena sampel kambing tersebut berasal dari Eropa (Italia) (Parma et al. 2003) dengan nilai uji bootstrap sangat tinggi (99%) (Gambar 11). Sekuen DNA daerah D-loop kambing lokal yang diteliti, jika disejajarkan dan dibandingkan dengan 26 sekuen DNA yang berasosiasi dari GenBank yang mewakili 6 kelompok utama (haplogroup) kambing di dunia berdasarkan asalusulnya secara maternal (Naderi et al. 2007; Joshi et al. 2004) menunjukkan bahwa keenam sub populasi kambing lokal Indonesia termasuk kedalam kelompok haplotip Lineage B (Gambar 12). Kambing yang termasuk dalam haplogroup B adalah kambing yang berdasarkan garis keturunan ibu (maternal) dan telah dilaporkan menyebar di daerah Asia Timur dan Asia Selatan termasuk Cina, Mongolia, Afrika Selatan, Afrika Utara, Laos, Malaysia, Pakistan dan India. Dari Gambar 12 terdapat nilai uji bootstrap dengan pengulangan 1000 kali pada Kambing Boer sangat rendah (30%) pada Kambing Boer dan (38-65%) Kambing Jawarandu menunjukkan bahwa kedua jenis kambing tersebut masih dekat kekerabatannya, karena Kambing Boer adalah merupakan persilangan Kambing Jamnapari dengan kambing lokal di Afrika Selatan. Nilai uji bootstrap Kambing Kacang dan Marica merupakan paling tinggi (99%) kemudian disusul oleh Kambing Jawarandu (92%), Muara (78%), Samosir (66%) dan Benggala (64-65%). Hasil uji bootstrap 1000 kali ulangan pada analisis Neighbour Joining dengan metode 2 parameter Kimura menunjukkan bahwa keenam sub populasi kambing lokal yang diteliti terbagi kedalam 6 kelompok atau bisa dikelompokkan menjadi 6 rumpun yang berbeda yaitu rumpun kambing pertama Kambing Kacang, kedua Kambing Marica, ketiga Kambing Samosir, keempat Kambing Jawarandu, kelima Kambing Muara dan keenam Kambing Benggala (Gambar 11) dengan nilai uji bootstrap di atas 60 % antara satu kelompok sub populasi kambing lokal yang satu dengan yang lainnya. Hampir sama dengan nilai uji bootstrap Kambing Muara juga rendah (46%), hal ini juga memperkuat dugaan bahwa Kambing Muara juga merupakan hasil persilangan kambing lokal dengan Peranakan Etawah (PE) di Indonesia, 54 saat ini Kambing Muara sudah mempunyai karakteristik morfologi tersendiri dan susunan basa nukleotida mempunyai kekhasan tersendiri. 68 65 Muara4 Muara5 78 Muara2 Muara Muara3 Muara1 Benggala3 Benggala5 64 Benggala2 Benggala 93 67 Benggala1 Benggala4 Jawarandu1 Jawarandu3 Jawarandu 99 Jawarandu4 63 Jawarandu5 Jawarandu2 Samosir2 Samosir3 Samosir Samosir4 67 Samosir1 Samosir5 Marica1 Marica2 Marica Marica5 99 Marica3 Marica4 Kacang2 71 Kacang1 Kacang 63 Kacang5 72 Kacang3 49 Kacang4 C.hircus NC_0050441 AF 533441 0.015 0.010 0.005 0.000 Gambar 11 Dendogram 6 sub populasi kambing lokal Indonesia berdasarkan ruas D-loop mtDNA (bootstrap 1000x) Kambing Muara bentuk ukuran tubuh hampir sama dengan PE, tetapi lebar dada relatif lebih panjang dan panjang telinga lebih pendek dan pola warna lebih bervariasi. Kambing Jawarandu bentuk tubuh lebih kecil, telinga pendek 55 dan warna bulu tubuh relatip lebih bervariasi jika dibandingkan dengan Kambing Etawah dan Kambing Peranakan Etawah. 33 11 10 34 1465 21 52 65 30 20 23 85 32 70 97 59 99 68 54 99 96 85 42 74 55 96 80 97 17 19 51 74 41 27 62 26 29 43 93 99 76 99 0.06 0.05 0.04 0.03 0.02 0.01 Muara2 Muara4 Muara5 BOER-AFRIKA SELATAN-(GQ141235) Muara3 Muara1 Jawarandu2 Jawarandu3 Jawarandu4 Jawarandu1 Benggala3 Benggala5 Benggala4 Benggala1 Benggala2 LAOS-NATIVE-(AB044303) Samosir5 Samosir3 Samosir2 Samosir4 AZERI-AZERBAIJAN-(EF617706) Jawarandu5 MONGOLIAN-GOAT-(AJ317833) MATOU-CHINA-(DQ121578) Samosir1 Marica4 Marica1 Marica5 Marica2 Marica3 Kacang2 Kacang1 Kacang5 Kacang3 Kacang4 TAIHANG-CHINA-(DQ188893) PASHMINA-INDIA-(AY155952) PINQAU-AUSTRIA-(EF617701) GURCU-TURKEY-(EF618535) BALADI-EGYPT-(EF617727) NUBIAN-ITALY-(FJ571542) ALPINE-FRANCE-(EF617779) BANJIAO-CHINA-(DQ121491) PUNJAB-GOAT-PAKISTAN-(AB162215) BLACK_BENGAL-INDIA-(AY155721) IRANIAN-GOAT-(EF617945) JAMNAPARI Capra hircus -Vietnam (AF533441) ANGORA-CHINA-(GQ141232) Capra_hircus_(AF533441) MALTESE-ITALY-(FJ571532)_ BARBARI_INDIA-_(AY155708) SWITZERLAND-GOAT-(AJ317838) SPANISH-GOAT-(EF618413) GIRGENTARA)-SICILY-(DQ241349 GIRGENTARA-SICILY-(DQ241351) B D G A C F 0.00 Gambar 12 Posisi 6 sub populasi kambing lokal Indonesia dalam dendogram kambing-kambing di dunia berdasarkan ruas D-loop mtDNA 56 Kambing Benggala mempunyai nilai bootstrap cukup jauh (65%) dari Kambing Kacang. Berdasarkan performans karakteristik tubuh dan warna bulu Kambing Benggala diduga merupakan persilangan Kambing Black Bengal dengan kambing lokal yang telah beradaptasi dengan kondisi agro-ekosistim di Pulau Timor dan Pulau Flores (Propinsi Nusa Tenggara Timur). Kambing Jawarandu, Muara dan Benggala merupakan persilangan kambing dari luar dengan kambing lokal, yang kemudian terjadi proses adaptasi dengan kondisi agro-ekosistim lokal sehingga penampilan produksi dan karakteristik kambing yang diteliti telah berubah jika dibandingkan dengan karakteristik rumpun asal kambing itu sendiri. Seperti Kambing Benggala jika dibandingkan dengan Kambing Black Bengal tubuh Kambing Benggala relatip lebih kecil, tetapi bentuk telinga dan warna bulu sama-sama warna hitam dan coklat tua pada umumnya. Berdasarkan analisis Median joining network dari 30 sekuen nukleotida daerah D-loop kambing yang diamati terdapat 50 situs yang bersifat polimorfik, terdapat sebanyak 19 haplotipe runutan DNA yang unik. Aliran gen berasal dari Kambing Kacang sebagai kambing asli di Indonesia, yang kemudian mengalami adaptasi sesuai dengan kondisi agro-ekosistem dan perlakuan manajemen dan terjadi perubahan susunan nukleotida dan perubahan fenotipik yang dikenal dengan Kambing Marica di daerah Sulawesi Selatan dan Kambing Samosir di daerah Kabupaten Samosir. Mutasi susunan nukleotida paling banyak di jumpai antara Kambing Kacang dengan Kambing Marica dan Samosir (Gambar 13). Perubahan mutasi ini terjadi diduga akibat adanya proses adaptasi dari Kambing Kacang sebagai kambing asli Indonesia dengan kondisi lingkungan yang sangat berbeda seperti lamanya musim kering yang tinggi di daerah Sulawesi Selatan yang mengakibatkan pakan hijauan layu dan mengering sehingga sangat terbatas jumlahnya di ujung musim kemarau. Hal ini bisa dilihat dari perubahan (mutasi) susunan nukleotida dalam bentuk subtitusi pada situs–situs tertentu (Tabel 14). Pada Kambing Jawarandu, Benggala dan Muara ditemukan mutasi subtitusi dan insersi nukleotida diduga akibat proses adaptasi lingkungan dan merupakan hasil persilangan (proses hibridisasi) dengan rumpun kambing di luar kambing lokal yang terdapat di daerah setempat. 57 K=Kacang, M=Marica, S=Samosir, J=Jawarandu, R=Muara, B=Benggala Gambar 13 Median joining network dari 19 haplotip nukleotida daerah D-loop DNA mitokondria pada 6 sub populasi kambing lokal Indonesia Simpulan Keragaman genetik 6 kambing lokal Indonesia yang diamati adalah 0.014±0.002 dan ditemukan 50 situs susunan nukleotida yang polimorfik dan terdiri dari 19 haplotip yang unik. Keenam sub populasi kambing lokal Indonesia menunjukkan keragaman susunan nukleotida yang berbeda antara setiap kelompok dengan kelompok sub populasi kambing lainnya. Perbedaan susunan nukleotida yang khas pada setiap rumpun kambing dapat dipakai sebagai penciri DNA antar rumpun kambing. Berdasarkan analisis keragaman genetik pada sekuen kambing penelitian dan sekuen nukleotida dari GenBank diduga asal usul tetua secara maternal kambing lokal yang diamati termasuk kedalam kelompok utama (haplogroup) lineage B. KARAKTERISASI MOLEKULER PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA BERDASARKAN ANALISIS DNA KROMOSOM Y Pendahuluan Variasi genetik dari rumpun ternak lokal sangat penting untuk mempertahankan sumberdaya genetik yang tidak tergantikan dan juga bermanfaat untuk membentuk bibit ternak yang baru. Gen SRY terletak pada kromosom Y yang terdiri dari ekson tunggal. Gen ini bertanggungjawab pada penentuan jenis kelamin pada ternak mamalia (Sinclair et al. 1990, Prashant et al. 2008). Sampai saat ini hasil penelitian tentang keragaman genetik kromosom Y pada kambing lokal Indonesia masih sangat terbatas. Tujuan penelitian ini adalah untuk mempelajari keragaman genetik Kromosom Y bagian gen SRY dan hubungan genetik antar sub populasi menurut garis keturunan secara paternal pada 6 sub-populasi kambing lokal Indonesia. Bahan dan Metode Sampel Darah Kambing Penelitian Sebanyak 18 sampel dipilih dari 77 sampel darah kambing jantan yang dikoleksi yang terdiri masing-masing 3 sampel per kelompok 6 sub populasi kambing. Ekstraksi DNA Perlakuan sama seperti yang dilakukan pada Ekstraksi DNA untuk analisis mitokondria. Amplifikasi DNA Amplifikasi genom mitokondria menggunakan primer AF126 (forward) 5’CCAGATCGATGTAGAGACAT-3’ dan AF127 (reverse) 5’TGCAATTTA CAAAGAGGTGGAA-3’. Primer AF126 menempel pada basa ke 3 925 - 3 947 nt dan AF127 menempel pada basa ke 4 675 - 4 696 nt dari runutan DNA kromosom Y pada sekuen lengkap Capra hircus dengan no akses GenBank EU 59 581862. Komposisi reaksi PCR dalam volume 25µl adalah sampel DNA 2µl (10100ng), RBC Bioscience taq polymerase 1.25 unit beserta sistim bufernya, dNTP 0.4 nmol, MgCl2 0.2 mM, primer AF22 dan AF23 masing-masing 1 nmol. Kondisi PCR yang digunakan untuk proses amplifikasi adalah tahap denaturasi awal pada suhu 94oC selama 3 menit, tahap denaturasi pada suhu 94oC selama 45 detik, tahap penempelan primer (annealing) pada suhu 62oC selama 50 menit, dan tahap polimerasi (extension) pada suhu 72oC selama 1 menit yang diulang selama 30 siklus, kemudian reaksi PCR diakhiri dengan polimerasi akhir pada suhu 72oC selama 5 menit. dilakukan menggunakan Visualisasi produk PCR tehnik elektroforesis gel poliakrilamid (PAGE) 6% dalam bufer 1xTBE (Tris-HCl 10 Mm, asam borat 1 M, EDTA 0.1 mM). Elektroforesis dijalankan pada kondisi 200 mV selama 50 menit, kemudian proses dilanjutkan dengan pewarnaan sensitive perak (Tegelstrom 1986) yang di modifikasi. Perunutan DNA Produk amplifikasi yang menunjukkan pita tunggal kemudian dimurnikan dan dijadikan cetakan dalam reaksi PCR. Primer yang digunakan dalam proses PCR untuk perunutan nukleotida sama dengan primer yang digunakan untuk amplifikasi. Reaksi PCR dilakukan dengan menggunakan metode dideoxi terminator dengan dNTP berlabel (big dye terminator). Perunutan nukleotida menggunakan mesin ABI Prism 3700-Avant Genetic Analyzer. Analisis Data Runutan nukleotida yang diperoleh menggunakan program Bio Edit versi 6.0.7. kemudian diedit dengan Urutan DNA yang telah diedit disejajarkan dengan beberapa runutan DNA dari kelompok Capra hircus yang dipublikasikan dalam GenBank (http://ncbi.nlm.nih.gov). Data yang diambil sebagai pembanding adalah gen SRY kromosom Y Capra hircus dengan no.akses EU 581862. Proses pensejajaran menggunakan ClustalW versi 8.1 yang tertanam dalam program MEGA 4 (Tamura et al. 2007) yang kemudian diedit lagi secara manual. Pengeditan hasil pensejajaran dilakukan dengan unitunit ruas DNA berulang. 60 Analisis yang dilakukan meliputi penghitungan komposisi nukleotida, laju subsitusi, jarak genetik berdasarkan ruas DNA kromosom Y pada segmen SRY. Proses analisis dilakukan dengan menggunakan program MEGA versi 4 (Tamura et al. 2007). Perhitungan nilai jarak genetik dilakukan berdasarkan model subsitusi Kimura 2 parameter (K2P). Rekonstruksi pohon filogeni dilakukan berdasarkan ruas daerah gen SRY untuk semua nukleotida yang bersifat parsimoni. Rekonstruksi pohon filogeni keduanya dilakukan menggunakan metode Neigbour Joining (NJ) dengan bootstrap 1000 kali. Untuk mengetahui jumlah haplotip dan penyebaran mutasi nukleotida digunakan analisis MedianJoining Network versi 4.6 (Fluxus Technology Ltd., 2005). Hasil dan Pembahasan Polimorfisme DNA Kromosom Y Amplifikasi gen SRY pada daerah Kromosom Y menggunakan pasangan primer AF126 dan AF127 yang didesain sendiri berdasarkan sekuen komplit gen SRY pada Capra hircus dari GenBank dengan nomor akses EU 581862. Dari hasil analisis sekuensing setelah diedit secara manual menggunakan program Bio Edit dan kemudian digabungkan antara hasil sekuen forward dengan hasil sekuen reverse maka diperoleh hasil keseluruhan sekuen nukleotida sepanjang 773 pb dengan letak posisi sekuen diperkirakan antara situs ke 3 925 – 4 697 pb (Gambar 14). Setelah saling disejajarkan antara hasil sekuen nukleotida dari keenam sub populasi kambing lokal yang diamati maka ditemukan 6 situs polimorfik (variable site) yang terdiri atas 1 singleton variable site yang ditandai dengan adanya insersi nukleotida T pada sub populasi Kambing Benggala, dan 5 parsimony informative sites yang ditandai dengan adanya mutasi substitusi pada setiap sub populasi kambing lokal lainnya. 61 Gambar 14 Struktur ruas gen SRY yang diapit oleh primer AF126 dan AF127 pada 6 sub populasi kambing lokal Indonesia Jika dibandingkan dengan sekuen nukleotida gen SRY pada Capra hircus (EU 581862) dari GenBank disini dapat dilihat terdapat mutasi yang sama pada keenam sub populasi kambing lokal Indonesia yaitu perubahan mutasi nukleotida dari G berubah menjadi A pada situs ke 398 pada penelitian ini. Runutan sekuen nukleotida yang bersifat polimorfisme pada Kromosom Y keenam sub populasi kambing lokal dibandingkan dengan Capra hircus (EU 581862) dapat dilihat pada Gambar 15. Terdapat 4 haplotipe berdasarkan 6 situs nukleotida yang bersifat polimorfik, dimana situs polimorfisme Kambing Kacang sama dengan Kambing Jawarandu, situs polimorfisme Kambing Marica sama dengan Kambing Samosir, sedangkan Kambing Muara dan Benggala berbeda satu sama lain. Runutan basa-basa nukleotida yang polimorfik terletak pada situs ke 330 - 754 berupa subtitusi dan insersi. Berdasarkan runutan nukleotida yang disejajarkan ditemukan substitusi nukleotida khas yang bisa digunakan sebagai penciri 6 kambing lokal jika dibandingkan dengan Capra hircus (EU 581862), akan tetapi Kambing Shiba dari Jepang sama dengan situs nukleotida C. hircus. Perubahan susunan (mutasi) nukleotida berupa subtitusi ditemukan pada situs nukleotida Kambing Kacang dan Jawarandu di situs ke 398 (G-A), Marica dan Samosir di situs ke 330 (A-T), 398 (G-A), 701 (T-C), Muara di situs 330 (A-T), 398(G-A), 701 (T-C), 718 (A-G), 754 (C-T) dan Benggala di situs 330 (A-T), 1398(G-A), 701(T-C), 718 (A-G), 754 (C-T). Mutasi insersi nukleotida T hanya ditemukan di situs ke 369 pada Kambing Benggala. 62 3333333333 3333333333 3333333334 7777777777 7777777777 7777777777 2222222223 6666666667 9999999990 0000000001 1111111112 5555555556 1234567890 1234567890 1234567890 1234567890 1234567890 1234567890 C.hircus_EU581862 TAATTTTAAA AGAATTTG-G CTCTGTTGAT TTCTAAAGCA CTTTCTGATA TTTCCACCTC Benggala1 .........T ........T. .......A.. C......... .......... ...T...... Benggala .........T ........T. .......A.. C......... .......... ...T...... Benggala3 .........T ........T. .......A.. C......... .......... ...T...... Jawarandu1 .......... ........-. .......A.. .......... .......... .......... Jawarandu2 .......... ........-. .......A.. .......... .......... .......... Jawarandu3 .......... ........-. .......A.. .......... .......... .......... Kacang1 .......... ........-. .......A.. .......... .......... .......... Kacang2 .......... ........-. .......A.. .......... .......... .......... Marica1 .........T ........-. .......A.. C......... .......... .......... Marica2 .........T ........-. .......A.. C......... .......... .......... Muara1 .........T ........-. .......A.. C......... .......G.. ...T...... Muara2 .........T ........-. .......A.. C......... .......G.. ...T...... Samosir .........T ........-. .......A.. C......... .......... .......... Shiba_goat_D82963 .......... ........-. .......... .......... .......... .......... Gambar 15 Polimorfisme nukleotida gen SRY pada 6 sub populasi kambing lokal Indonesia (tiga baris pertama dibaca secara vertikal merupakan posisi nukleotida yang mengacu pada C. hircus GenBank no. akses EU581862 dan Shiba Goat no. akses D82963 Frekuensi Nukleotida dan Jarak Genetik Berdasarkan hasil sekuen DNA gen SRY analisis jarak genetik antara 6 sub populasi kambing lokal yang diamati dibandingkan dengan sekuen nukleotida pada Capra hircus (EU 581862) dari GenBank, maka jarak genetik paling tinggi adalah pada Kambing Muara (0.001), kemudian diikuti Kambing Benggala (0.008), Kambing Marica sama dengan Kambing Samosir (0.006), dan paling rendah terdapat pada Kambing Kacang sama dengan Kambing Jawarandu (0.002). Perbandingan jarak genetik antar sub populasi kambing lokal ditemukan bahwa tidak ada jarak perbedaan antara Kambing Kacang dengan Kambing Jawarandu (0.000), demikian juga antara Kambing Marica sama dengan Kambing Samosir (0.000). Jarak genetik paling tinggi dijumpai perbedaan antara Kambing Muara terhadap Kambing Kacang dan Jawarandu (0.008), kemudian dikuti jarak perbedaan antara Kambing Benggala terhadap Kambing Kacang dan Jawarandu (0.006). Perbedaan jarak genetik Kambing Marica dan Samosir terhadap semua sub populasi kambing lainnya sama (0.004), kecuali terhadap 63 Kambing Benggala (0.002). Jarak genetik berdasarkan susunan nukleotida DNA Kromosom Y Kambing Kacang, Benggala, Jawarandu, Muara, Samosir, Marica dan Capra hircus (EU 581862) dari GenBank dapat dilihat pada Tabel 16. Tabel 15 Matrik keragaman nukleotida gen SRY kromosom Y pada kambing lokal Indonesia C.hircus Kacang Sub populasi Capra hircus* Kacang 0.002 Samosir 0.006 0.004 Marica 0.006 0.004 Jawarandu 0.002 0.000 Muara 0.010 0.008 Benggala 0.008 0.006 *GenBank no. akses EU 581862 Samosir Marica Jawarandu 0.000 0.004 0.004 0.002 0.004 0.004 0.002 0.008 0.006 Muara Benggala 0.002 Keragaman genetik antar individu dalam kelompok sub populasi kambing penelitian yaitu sub populasi Kambing Samosir 0.004, Kambing Kacang dan Kambing Jawarandu 0.003, Kambing Marica 0.002, Kambing Muara 0.001 dan Kambing Benggala tidak ada perbedaan (0.000). Berdasarkan hasil analisis philogenik runutan nukleotida DNA kromoson Y kambing lokal Indonesia dengan nilai bootstrap 89% menunjukkan bahwa Kambing Kacang masih satu kelompok dengan Kambing Jawarandu dan Kambing Marica masih satu kelompok dengan Samosir (21%), sedangkan Kambing Benggala (65%) dan Muara (69%) membentuk kelompok terpisah satu sama lain (Gambar 16). Berdasarkan hasil analisis metode Neighbour Joining pada keenam sub populasi kambing yang diamati dapat dikelompokkan berdasarkan keragaman genetik secara paternal menjadi 4 kelompok, yaitu kelompok Kambing Kacang dan Jawarandu, kelompok Kambing Marica dan Samosir, kelompok Kambing Benggala dan kelompok Kambing Muara. 64 68 Muara1 21 19 64 23 22 Muara2 Benggala1 Benggala3 Benggala2 Marica1 Marica2 Samosir Jawarandu3 Jawarandu2 Kacang1 87 Kacang2 Jawarandu1 C.hircus EU581862 66 Shiba goat D82963 0.0005 Gambar 16 Dendogram Neighbour Joining berdasarkan runutan nukleotida gen SRY antara 6 sub populasi kambing lokal Indonesia Jika digabungkan dengan rumpun kambing out group dari beberapa situs nukleotida gen SRY di GenBank ditemukan bahwa keenam sub populasi kambing lokal Indonesia membentuk kelompok tersendiri terpisah dari kelompok kambing lainnya. Situs nukleotida Capra hircus (EU 581862) satu kelompok dengan Kambing Shiba dari Jepang dengan jarak genetik dengan kelompok kambing lokal Indonesia 63%, sedangkan jarak kambing lokal dengan Kambing Sangamneri dan Capra hircus (D0845) sangat jauh yaitu 100 % berbeda dengan kambing lokal Indonesia (diduga karenan hasil sekuen kromosom Y yang didapat lebih pendek). Sampai saat ini hasil penelitian gen SRY pada ternak kambing yang dilaporkan dan bisa diakses di GenBank masih sangat terbatas, sehingga belum bisa leluasa membandingkan dengan jenis rumpun kambing lainnya. Berdasarkan analisis Median-joining network terdapat 6 situs yang bersifat polimorfik, dan terdapat sebanyak 4 haplotip runutan DNA yang unik (Gambar 17) yaitu Kambing Marica dan Samosir (1 haplotip), Kambing Kacang dan Jawarandu (1 haplotip), Kambing Muara (1 haplotip) dan Kambing Benggala (1 haplotip). 65 M1 J1 Gambar 17 Median-joining network dari 4 haplotip gen SRY dari 6 sub populasi kambing lokal Indonesia: Kacang (K 1 ), Marica (M 1 ), Samosir (S 1 ), Jawarandu (J 1 ), Muara (R 1 ) dan Benggala (B 1 ) Diduga Kambing Marica dan Kambing Samosir berasal dari pejantan Kambing Kacang yang telah mengalami proses adaptasi terhadap lingkungan dan campur tangan perlakuan peternak yang diarahkan untuk tujuan produksi tertentu. Hal ini bisa dilihat dari perubahan (mutasi) susunan nukleotida dalam bentuk substitusi pada situs tertentu (Tabel 15). Sedangkan pada Kambing Jawarandu, Benggala dan Muara ditemukan mutasi subtitusi dan insersi nukleotida diduga akibat proses adaptasi lingkungan dan merupakan hasil persilangan (proses hibridisasi) dengan rumpun kambing diluar kambing lokal yang terdapat di daerah setempat. Simpulan Keragaman genetik 6 kambing lokal Indonesia yang diamati adalah 0.004 ± 0.002 dan ditemukan 6 situs susunan nukleotida yang polimorfik dan terdiri dari 4 haplotip yang unik, yaitu pada Kambing Kacang dan Jawarandu (1 haplotip), Marica dan Samosir (1 haplotip), Muara (1 haplotip) dan Benggala (1 haplotip). Keenam sub populasi kambing yang diamati dikelompokkan menjadi 4 haplotip kambing yang berbeda, dimana terdapat perbedaan susunan nukleotida yang khas sebagai penciri DNA antara satu genotip dengan genotip yang lainnya. Asal usul tetua secara paternal Kambing Jawarandu, Marica, Samosir diduga berasal dari Kambing Kacang. IDENTIFIKASI GEN GDF9 DAN HUBUNGANNYA DENGAN SIFAT PROLIFIK PADA KAMBING LOKAL INDONESIA Pendahuluan Sifat prolifik adalah kemampuan untuk melahirkan dua atau lebih anak sekaligus per periode kelahiran pada induk ternak. Sifat prolifik dikendalikan oleh gen-gen yang dikenal sebagai keluarga gen kesuburan (fecundity genes), yaitu bone morphogenetic protein receptor type 1B (BMPR1B) yang disebut juga dengan nama Fecundity Boorola (FecB) (Souza et al. 2001; Davis 2005, Davis et al. 2006); growth differentiation factor 9 (GDF9), disebut FecG (Hanrahan et al. 2004); bone morphogenetic protein 15 (BMP15) disebut dengan FecX (Hanrahan et al. 2004; Galloway et al. 2000). Tiga gen fekunditas diatas dikelompokkan sebagai TGF-β super family yang telah diidentifikasi pada mamalia. Beberapa mutasi pada gen GDF9 dilaporkan berhubungan dengan peningkatan laju tingkat ovulasi dan litter size pada ternak ruminansia kecil (Gilchrist et al. 2005) Gen GDF9 ini diekspresikan pada oosit dan sel-sel granulosa ovarium sejak tahap folikel primer sampai oosit diovulasikan. Gen GDF9 yang terdapat pada kromosom 5 pada domba, kambing dan sapi telah berhasil dipetakan yang terdiri atas dua ekson dan satu intron. Selain itu, gen GDF9 ini diketahui menyandikan prepropeptida sepanjang 453 residu asam amino. Polipeptida aktif adalah sepanjang 135 residu asam amino (Bodensteiner et al. 1999; Hanrahan et al. 2004). Pengaturan ekpresi GDF9 dan proses pematangan prepropeptida menjadi polipeptida aktif melibatkan ruas-ruas pengaturan ekspresi gen yang terdapat dalam promotor dan intron (Gilchrist et al. 2005). Secara alami ternak ruminansia kecil cenderung bersifat prolifik, termasuk kambing dan domba lokal Indonesia. Kambing Kacang mempunyai rata-rata anak sekelahiran (litter size) antara 1.56 – 1.98 ekor (Sodiq et al. 2003; Hoda 2008), kambing PE sekitar 1.3-1.7 anak per kelahiran (Sodiq et al. 2003), sedangkan Kambing Samosir dan Kambing Muara belum ada laporan yang menyebutkan kemampuan prolifikasinya. Beberapa program pemuliaan saat ini aktif untuk mengembangkan kambing yang mengarah pada peningkatan produksi daging, peningkatan pertumbuhan, daya adaptasi terhadap kondisi lingkungan yang sulit dan peningkatan sifat prolifik. 67 Hasil penelitian identifikasi keragaman gen BMPR1B dan BMP15 di ruas ekson 1, ekson 2 dan intron pada induk Kambing Kacang, PE, Muara dan Samosir dengan sampel yang sama dengan penelitian menunjukkan bahwa sekuen antara kelompok induk beranak kembar hampir sama dengan kelompok induk beranak tunggal atau bersifat monomorfik (Hasan et al. 2011). Pada saat ini informasi yang berhubungan dengan keragaman gen GDF9 pada ruas promotor masih belum ada laporan penelitian. Tujuan penelitian ini adalah untuk mengidentifikasi keragaman gen GDF9 pada dua kelompok induk, yaitu induk yang rata-rata beranak tunggal (nonprolifik) dan kelompok induk yang rata-rata beranak kembar (prolifik) pada Kambing Kacang, PE, Samosir dan Muara. Bahan dan Metode Penelitian ini dilaksanakan mulai bulan Mei 2010 sampai dengan Maret 2011 di Laboratorium Fungsi Hayati dan Perilaku Hewan, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor. Pengambilan Sampel Darah Kambing Pengambilan sampel darah Kambing Kacang dan Kambing Peranakan Etawah (PE) dilakukan di Kandang Percobaan Loka Penelitian Kambing Potong Sei Putih, Sumatera Utara. Sedangkan sampel darah Kambing Samosir diambil dari peternakan rakyat di Kabupaten Samosir dan Kambing Muara di peternakan rakyat di Kabupaten Tapanuli Utara, Propinsi Sumatera Utara. Sampel darah kambing yang diperoleh dari lapangan adalah 149 sampel antara DNA kelompok induk yang rata-rata beranak tunggal (wild type atau normal) disebut dengan kelompok non-prolifik dan dengan kelompok induk yang beranak kembar (mutant type atau mutan) yang disebut dengan kelompok induk prolifik (Tabel 17). Tabel 16 Distribusi kambing prolifik dan non prolifik No. Rumpun kambing Prolifik Non-prolifik 1 Kacang 14 11 2 Peranakan Etawah (PE) 14 15 3 Muara 9 26 4 Samosir 24 36 Total 61 88 Jumlah 25 29 35 60 149 68 Sampel darah diambil dari vena jugularis sekitar 2 ml menggunakan jarum venoject yang dihubungkan dengan tabung vakum. Darah yang diperoleh di lapangan langsung diawetkan dalam alkohol absolut 2x volume darah, kemudian darah dikocok dengan kuat hingga larut dalam alkohol. Ekstraksi DNA Ekstraksi genom DNA dilakukan menggunakan Genomic DNA Mini Kit for Fresh Blood (GeneAid) yang dimodifikasi. Modifikasi yang dilakukan bertujuan untuk menghilangkan alkohol sebelum dilakukan proses ekstraksi DNA. Sampel darah dalam alkohol sebanyak 1mL disentrifugasi 5 000 rpm selama 5 menit. Endapan sel dicuci dengan menambahkan aquades hingga volume total 1,5 mL dan didiamkan selama 20 menit. Pencucian dilakukan sebanyak dua kali. Sel-sel darah yang telah dicuci disuspensikan dengan bufer pelisis (GT bufer) 100µL, kemudian dilisis lebih lanjut dengan enzim Proteinase K 0.01-0.5 µg/mL dan diinkubasi pada suhu 600C selama 30 menit. Langkah selanjutnya yaitu, pemisahan bahan organik non-DNA dan pemurnian molekul DNA dilakukan sesuai dengan prosedur dari perusahaan. Amplifikasi Gen GDF9 Amplifikasi gen GDF9 pada ruas promotor (5’flanking region) dilakukan dengan mesin TaKaRa Thermal Cycler menggunakan primer AF 211 forward CCTCAGTCTTCTCCTCGGTTCC dan AF 212 reverse CTGGAAGTGG GAGAAGTGG yang mengacu pada Dong et al. (2005). Amplifikasi gen GDF9 menghasilkan ruas DNA dengan panjang 1972 pb berdasarkan penempelan primer pada sekuen DNA Capra hircus dengan kode accession number EF446168. Reaksi PCR dilakukan dalam volume 12 µl, yang terdiri atas sampel DNA sekitar 10 ng, primer forward dan reverse masing-masing 1 ng, dan KAPA Taq Ready Mix DNA polymerase (KAPATaq DNA polymerase 1 unit, bufer polimerase dengan Mg2+.25 mM dan setiap dNTP masing-masing 0,4 mM). Kondisi PCR, yaitu predenaturasi 940C selama 5 menit, (denaturasi 940C selama 60 detik, penempelan primer 58 0C selama 90 detik, pemanjangan 720C 90 detik) sebanyak 30 siklus, pemanjangan akhir pada suhu 720 C selama 10 menit, dan penyimpanan dilakukan pada suhu 40C. Amplikon dideteksi dengan 69 elektroforesis gel poliakrilamida 6% yang dilanjutkan dengan pewarnaan perak (Byun et al. 2009). Perunutan DNA Untuk mengetahui posisi dan jenis nukleotida yang mengalami mutasi maka dilakukan perunutan DNA (sekuensing). Beberapa amplikon gen GDF9 pada setiap sub populasi dicampur menjadi satu berdasarkan sifat prolifikasi induk kambing. Campuran amplikon selanjutnya disekuensing menggunakan primer yang sama dengan amplifikasi awal. Teknik ini dilakukan untuk memindai adanya mutasi dengan lebih cepat. Hal ini dilakukan atas dasar laju mutasinya sangat rendah. Analisis Data Runutan nukleotida yang diperoleh kemudian diedit secara manual dengan bantuan program Bio Edit versi 6.0.7. Urutan DNA yang telah diedit disejajarkan dengan beberapa runutan DNA dari kelompok Capra hircus yang dipublikasikan dalam GenBank (http://ncbi.nlm.nih.gov). Proses pensejajaran menggunakan Clustal W versi 8.1 yang ada dalam program MEGA 4 (Tamura et al. 2007) yang kemudian diedit lagi secara manual. Analisis yang dilakukan meliputi penghitungan komposisi nukleotida, laju mutasi delesi dan substitusi. Proses menentukan struktur gen yang diperoleh untuk mencari situs yang homolog dengan program BLASTN versi 2.2.25 dan dianalisis dengan menggunakan program MEGA versi 4. Hasil dan Pembahasan Amplifikasi gen GDF9 menggunakan primer AF 211 dan AF 212 menghasilkan fragmen DNA dengan panjang 1296 pb. Mutasi pada gen GDF9 dapat meningkatkan laju tingkat ovulasi dan litter size. Ruminansia kecil dengan genotip heterozigot carier akan meningkatkan laju ovulasinya rata-rata 1.5 dengan rata-rata litter size 1.0. Sedangkan genotip homozigot carrier akan meningkatkan laju ovulasi rata-rata 3.0 dengan rataan litter size 1.5 (Davis 2005). Jika hasil amplikon gen GDF9 sudah dipotong dengan enzim masih meragukan, dapat diverifikasi dengan metode sekuensing. 70 Hasil sekuensing pada gen GDF9 menunjukkan adanya polimorfisme satu pasang basa nukleotida berupa mutasi substitusi G – A pada posisi basa nukleotida ke 836 pada Kambing Kacang dan PE, serta mutasi substitusi G – C pada Kambing Kacang di situs ke 1019 (Gambar 18). 8888888888 6666666667 1234567890 KACANG-tunggal(08-K1) CAGAGGCAAG KACANG-kembar(07-K2) ..A....... PE-tunggal(04-PE1) .......... PE-kembar(15-PE3) ..A....... MUARA-tunggal(06-R1) .......... MUARA-kembar(05-R2) .......... SAMOSIR-tunggal(14-S1) .......... SAMOSIR-kembar(12-S2) .......... Nt 863 (G-A) 1111111111 0000000000 1111111112 1234567890 ATGAATGAGC ........C. .......... .......... .......... .......... .......... .......... Nt 1019 (G-C) Exon 1 5’UTR Exon 2 Intron 3’UTR Promotor 1nt 1260nt 23nt wild mutan 2170 nt 3294 nt 4258 nt 5515 nt wild mutan Gambar 18 Mutasi nukleotida gen GDF9 ruas promotor pada kelompok induk prolifik (mutan) dan induk non-prolifik (wild) pada Kambing Kacang dan Peranakan Etawah (Tiga baris pertama dibaca secara vertikal merupakan posisi nukleotida). 71 Keragaman gen GDF9 dengan sifat prolifik pada kambing bervariasi dan dipengaruhi oleh rumpun ternak dan posisi ruas DNA yang diidentifikasi. GDF9 ruas ekson 1 dan ekson 2 pada Kambing Black Bengal yang dikenal prolifik adalah monomorfik, tetapi pada Kambing Jining Grey dilaporkan polimorfik (Feng et al. 2010). Hasil penelitian ini hampir sama dengan penelitian Polley et al. (2009). Adanya alel-alel mutasi nukleotida gen GDF9 di ruas promotor pada Kambing Kacang dan PE dapat menambah temuan identifikasi keragaman gen GDF9 untuk keperluan seleksi calon induk yang prolifik pada ternak domba dan kambing, seperti yang tertera pada Tabel 18. Tabel 17 Jenis-jenis mutan gen GDF9 pada ternak domba dan kambing Mutasi Ekson Mutasi asam amino DNA G 260 A 2 Arginin-Histidin G 471 T Tidak ada mutasi G 477 A Tidak ada mutasi G 721 A Glutamin-Lys G 978 G Tidak ada mutasi G 994 A Val - Ile G 1111 A Val - Met G 1184 T Ser - Phe A 562 C 2 Glutamin-Prolin G 26 A G 792 A A 183 C C 336 T 1 2 1 2 Tidak ada mutasi Valine - Isoleusine Tidak ada mutasi Valine - Isoleusine Runutan nukleotida gen Jenis ternak Referensi Domba Domba Domba Domba Domba Domba Domba Domba Kambing Qianbei pockmarked Kambing Jining Grey Kambing Jining Grey Kambing Jining Grey Kambing Jining Grey Kambing Jining Grey GDF9 bersifat Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Hanrahan et al. 2004 Ren et al. 2010 Wu et al. 2010 Wu et al. 2010 Chu et al. 2011 Chu et al. 2011 Feng et al. 2011 polimorfik dan diduga berhubungan dengan pengaturan sifat prolifikasi pada Kambing White goat (Xuqin et al. 2009), Jining Grey (Feng et al. 2010), sementara gen ini juga bersifat monomorfik pada Kambing Black Bengal (Polley et al. 2009), Boer dan Huanghuai Goats (He 2010). Fenomena tersebut juga dijumpai pada penelitian ini, yaitu fenomena keragaman mutasi gen GDF9 di ruas promotor pada keempat sub populasi kambing lokal yang diamati bervariasi. Berdasarkan hasil penelitian tersebut dapat ditarik kesimpulan bahwa ada atau tidaknya polimorfisme nukleotida gen-gen fekunditas pada kambing masih sangat bervariasi antar rumpun-rumpun kambing dan hubungan gen tersebut dengan pengaturan tingkat prolifikasi pada induk kambing. Temuan alel-alel mutasi gen ini diduga dapat 72 digunakan sebagai salah satu parameter atau metode dalam upaya seleksi calon induk bibit kambing yang prolifik untuk meningkatkan produktivitas dan efisiensi pemeliharaan ternak kambing. Berdasarkan kejadian mutasi yang bersifat parsimony runutan nukleotida gen GDF9 ruas promotor (Lampiran 7) menunjukkan bahwa kambing lokal Indonesia berada dalam satu klaster dan lebih dekat dengan Kambing Jining grey dari Cina (Gambar 19). 91 KACANG-tunggal 62 KACANG-kembar 60 26 29 C.hircus PE-kembar MUARA-tunggal MUARA-kembar PE-tunggal SAMOSIR-tunggal 100 SAMOSIR-kembar 0.01 Gambar 19 Dendogram kambing lokal Indonesia berdasarkan runutan nukleotida ruas promotor gen GDF9 metode NJ bootstrap 1000x Simpulan Mutasi gen GDF9 ruas promotor bersifat polimorfik yang mengekpresikan adanya hubungan mutasi gen GDF9 dengan sifat prolifik pada Kambing Kacang dan PE. Keragaman gen GDF9 pada induk Kambing Muara dan Samosir bersifat monomorfik. PEMBAHASAN UMUM Sesuai dengan tujuan penelitian untuk mengidentifikasi karakteristik morphometrik dan genetik pada 6 sub populasi kambing lokal Indonesia dan strategi pemanfaatannya secara berkelanjutan, kambing lokal pada 6 sub populasi yang diamati memiliki karakteristik fenotipik (ukuran-ukuran tubuh, bobot badan dan warna tubuh) yang beragam. Rataan ukuran-ukuran tubuh dan bobot badan berbeda antara satu sub populasi dengan sub populasi kambing lokal lainnya. Karakter panjang badan, tinggi pundak, tinggi pinggul, lebar dada, lingkar dada, lingkar kanon, lebar tengkorak, tinggi tengkorak pada kambing lokal yang diamati menunjukkan nilai koefisien keragaman tinggi. Seleksi juga dapat dilakukan pada warna dan pola warna tubuh kambing lokal yang beragam apabila diinginkan untuk meningkatkan pola warna tertentu. Menurut Astuti (2004) seleksi ternak lokal dengan memanfaatkan keragaman karakteristik sifat produksi dan reproduksi akan meningkatkan produktifitas, disamping memiliki dampak penting terhadap pelestarian sumber daya genetik ternak. Dari keenam sub populasi kambing yang diamati terdapat tiga sub populasi yang sudah menunjukkan pola warna yang spesifik yaitu Kambing Benggala cenderung berwarna hitam, Kambing Samosir sebagian besar berwarna putih, sedangkan Kambing Muara sebagian besar berwarna putih dengan belang hitam ataupun coklat kemerahan (merah bata). Pada Kambing Kacang, Marica dan Jawarandu pola warna masih sangat beragam, sehingga peluang untuk mengarah ke pola warna tertentu untuk membentuk pola warna yang dinginkan masih sangat terbuka. Adanya perbedaan susunan dan komposisi basa-basa nukleotida antara kambing-kambing lokal Indonesia (Kacang, Jawarandu, Marica, Muara, Samosir dan Benggala) berdasarkan analisis D-loop parsial (957 bp) DNA mitokondria, menunjukkan ada perbedaan materi genetik antara kambingkambing tersebut. Demikian juga apabila dibandingkan keenam sub populasi kambing lokal Indonesia yang diamati dengan kambing-kambing Eropa, Afrika, Cina dan kambing dari data GenBank, maka jelas terdapat perbedaan materi genetik Kambing Kacang, Jawarandu, Marica, Muara, Samosir dan Benggala dengan kambing dari kelompok tersebut. Menurut Naderi et al. (2007) berdasarkan keragaman genetik DNA mitokondria pendugaan asal-usul ternak kambing dapat dkelompokkan menjadi enam kelompok utama (haplogroup 74 lineage). Berdasarkan hasil analisis sekuen yang diperoleh keenam sub populasi kambing lokal yang diamati termasuk kedalam lineage B yaitu kelompok kambing yang menyebar di Asia Timur dan Asia Selatan, Cina, Mongolia, Afrika Selatan, Afrika Utara, Laos, Malaysia, Pakistan dan India. Selain 6 sub populasi kambing lokal Indonesia yang diamati dalam penelitian ini, di Indonesia juga dijumpai jenis rumpun kambing lainnya yang termasuk dalam kelompok utama (haplogroup lineage) A yang berupa rumpun-rumpun kambing yang daerah penyebarannya dari Eropa seperti Kambing Saanen, Kambing Nubian dan Jamnapari (Etawah). Susunan nukleotida yang khas (unik) ini dapat dipakai sebagai salah satu penciri untuk mengidentifikasi kambing lokal Indonesia. Penciri khas kambing lokal Indonesia dapat diperoleh setelah dibandingkan dengan nukleotida Capra hircus (AF533441) dari GenBank yaitu perubahan nukleotida pada situs ke 201, 386, 625 (A-G); 113, 633, 697 (C-T); 298 (G-A); 537 (G-C); 597 (T-C) dan 812 (C-A). Sedangkan setiap sub populasi susunan basa DNA yang unik yang dapat dipakai sebagai penciri khas pada Kambing Kacang, Marica, Jawarandu, Muara, Samosir dan Kambing Benggala. Berbeda dengan keragaman susunan genetik yang tinggi pada metode analisis DNA mitokondria, pada analisis DNA kromosom Y pada segmen gen SRY tingkat keragaman susunan genetik relatif lebih rendah. Berdasarkan primer yang didesain sendiri diperoleh hasil analisis sekuen susunan basa nukleotida sepanjang 499 bp dan terdiri dari 6 situs yang bersifat polimorfik dan ditemukan 4 haplotip susunan nukleotida yang khas yang dapat dipakai sebagai salah satu acuan untuk mengetahui asal-usul berdasarkan keturunan secara paternal (menurut garis keturunan ayah, tetua jantan). Berdasarkan haplotip tersebut ditemukan bahwa Kambing Kacang satu kelompok dengan Kambing Jawarandu, disini memperkuat dugaan Kambing Jawarandu merupakan keturunan dari persilangan pejantan Kambing Kacang dengan induk Kambing Peranakan Etawa (PE) atau Etawah, hal ini juga bisa dilihat dari penampilan ciri-ciri fenotipik yang berbeda dengan Kambing PE yang antara lain panjang dan lebar telinga lebih pendek, tinggi pundak lebih rendah sehingga performan Kambing Jawarandu lebih mengarah ke Kambing Kacang dari pada ke arah Kambing Etawah (Jamnapari). Kambing Marica dan Kambing Samosir satu kelompok tetapi keragaman genetiknya tidak berbeda nyata dengan kelompok Kambing Kacang dan Kambing Jawarandu. Diduga Kambing ini berasal dari Kambing Kacang yang 75 telah mengalami adaptasi dengan lingkungan setempat dalam jangka waktu yang lama. Kambing Marica diduga mengalami adaptasi dengan kondisi lingkungan di Sulawesi Selatan dengan masa musim kemarau yang panjang (6-9 bulan per tahun), sehingga mulai dari pertengahan sampai akhir musim kering rumput sudah mengering dan jumlahnya relatif terbatas sehingga kambing beradaptasi, secara fenotip dapat dilihat ukuran tubuh mengecil dan bulu relatif coklat kemerahan dan mengkilat. Pada Kambing Samosir terjadi karena seleksi campur tangan manusia, dimana peternak ingin memelihara kambing yang berwarna putih untuk keperluan ritual keagamaan, sehingga kambing yang berwarna putih dapat dijual dengan harga yang lebih tinggi. Tetapi di beberapa tempat karakteristik ukuran tubuh dan pola warna Kambing Samosir ada yang agak mirip dengan Kambing Saanen, diduga Kambing Samosir mungkin juga merupakan hasil persilangan Kambing lokal dengan Kambing Saanen, tetapi performan tubuh dan bobot tubuh relatif lebih kecil jika dibandingkan dengan Kambing Saanen. Jika ditinjau dari runutan nukleotida daerah D-loop DNA mitokondria dan DNA kromosom Y segmen gen SRY lebih dekat dengan Kambing Kacang dan Kambing Marica. Kambing Muara diduga merupakan kambing persilangan Kambing Peranakan Etawah dengan kambing lokal di daerah Kabupaten Tapanuli Utara yang telah beradaptasi dengan lingkungan setempat. Dengan topografi lingkungan dataran tinggi yang berbukit-bukit, ketersediaan pakan relatif cukup membentuk postur tubuh Kambing Muara relatif besar dan kompak, sangat cocok untuk ternak kambing penghasil daging. Juga berdasarkan pengamatan dari jumlah anak perkelahiran, beberapa induk beranak empat dapat hidup sampai lepas sapih di tingkat petani. Hal ini diduga disamping suplay makanan cukup juga di dukung produksi susu yang relatif baik untuk mendukung pertumbuhan anak kambing sejak lahir. Kambing Benggala umumnya berwarna hitam dan ada coklat tua, telinga separuh di ujung jatuh, tinggi pundak relatif pendek . Kambing ini terpisah jauh dari Kambing Kacang (0.006). Diduga kambing ini merupakan hasil persilangan Kambing Black Bengal dengan kambing lokal di daerah Nusa Tenggara Timur. Diduga ternak tersebut masuk ke daerah NTT melalui para pedagang dari India, Pakistan atau Sri Lanka yang di bawa serta untuk menghasilkan susu dan daging. Masyarakat menyebutnya Kambing Benggala (mungkin berasal dari 76 India dari kata Benggali) atau karena warna kambing tersebut berwarna hitam pada umumnya. Berdasarkan jarak genetik pada ukuran-ukuran tubuh, penampilan, pola warna dominan, keragaman genetik berdasarkan D-loop DNA mitokondria dan kromosom Y segmen gen SRY, diduga setiap populasi kambing lokal Indonesia yang diamati telah membentuk karakteristik morfologi dan karakteristik genetik tersendiri yang khas sehingga bisa dikelompokkan menjadi 6 rumpun atau galur kambing yaitu Kambing Kacang, Kambing Jawarandu, Kambing Marica, Kambing Muara, Kambing Samosir dan Kambing Benggala. Berdasarkan hasil penelitian ini kambing lokal Indonesia dapat dipetakan keragaman genetiknya untuk tujuan konservasi sumberdaya genetik ternak lokal dan pengembangan ternak lokal guna mendukung ketahanan pangan dan peningkatan pendapatan masyarakat peternak kambing. Dengan diperolehnya status dan karakteristik morfologi kambing lokal ini, perlu dilanjutkan karakterisasi produksi, reproduksi dan daya tahannya terhadap penyakit dan perubahan lingkungan di daerah tropis. Sampai saat ini laporan hasil penelitian pendahuluan tentang hubungan ketiga gen fekunditas tersebut masih sangat bervariasi, sebagian dilaporkan ada polimorfisme dan sebagian lagi melaporkan tidak terdapat polimorfisme. Beberapa hasil penelitian juga melaporkan bahwa keragaman Gen GDF9 bersifat monomorfik pada Kambing Boer, Haimen, Huanghuai, Nubi, dan Kambing Matou (Guo-Hua et al. 2008). Hasil penelitian menunjukkan tingkat keragaman gen GDF9 pada kelompok induk yang rata-rata beranak kembar dan kelompok induk yang rata-rata beranak tunggal pada Kambing Kacang, PE, Samosir dan Muara bersifat monomorfik. Alternatif lain yang mungkin dilakukan adalah dengan mengidentifikasi gen-gen lain yang diduga berhubungan dengan sifat fekunditas pada induk kambing yang beranak kembar, seperti beberapa jenis gen yang diduga berhubungan dengan sifat beranak kembar pada kambing antara lain ; gen FSHB yang berhubungan dengan superovulasi pada Kambing Matou (Zhang et al. 2011), gen KiSS-1 diduga berhubungan dengan sifat prolifik pada Kambing Guanzhong dan Xinong Saanen (Huo et al. 2011), gen INHA masih diduga mengontrol sifat prolifik pada Kambing Matou dan Kambing Haimen (GuoHuo et al. 2007). Fenogram berdasarkan data karakter morfometrik, dendogram berdasarkan mitokondria menunjukkan bahwa masing-masing sub populasi berbeda, sehingga dapat dikelompokkan menjadi 6 rumpun kambing lokal. Sedangkan dendogram berdasarkan nukleotida kromosom Y dan gen GDF9 ruas 77 promotor menunjukkan bahwa secara paternal Kambing Kacang dan Jawarandu termasuk satu kelompok, serta Kambing Muara dengan PE satu kelompok. Ini memperkuat dugaan bahwa Kambing PE merupakan tetua jantan dari Kambing Muara. Hasil penelitian ini memperoleh data dan informasi pendahuluan tentang pemetaan keragaman sumberdaya genetik kambing lokal Indonesia khususnya untuk rumpun atau galur Kambing Kacang, Marica, Muara, Jawarandu, Samosir dan Kambing Benggala. Berdasarkan Peraturan Menteri Pertanian Nomor 35/Permentan/OT.140/8/2006, tentang Pedoman Pelestarian dan Pemanfaatan Sumberdaya Genetik, upaya pelestarian dan pengembangan kambing lokal Indonesia penting dilakukan untuk mengantisipasi kehilangan sumberdaya genetik plasma nuftah kambing akibat adanya persilangan dan pemotongan ternak yang tidak seimbang dengan produksinya. Untuk menghindari kehilangan sumberdaya genetik kambing lokal Indonesia perlu dilanjutkan karakterisasi produktivitas dan keunggulan potensi genetiknya dan penelitian pengembangan, kemudian disertai dengan sosialisasi pengenalan ternak kambing lokal tersebut terutama di wilayah dimana populasi aslinya berada (in-situ), sehingga timbul minat untuk memelihara dan mengembangkannya. Sampai saat ini pada umumnya sistim pemeliharaan ternak kambing lokal masih diserahkan kepada kebaikan alam, dengan pemeliharaan secara tradisional (kearifan lokal). Disadari atau tidak ternak kambing lokal tersebut berperan menambah pendapatan keluarga petani di daerah pedesaan dengan sistim pemeliharaan dengan biaya relatip rendah (low input system), sesuai dengan kondisi agro-ekosistem setempat dan ternak dapat hidup dan berproduksi. Jika berhasil ditetapkan secara ilmiah rumpun-rumpun atau galur kambing lokal berdasarkan hasil-hasil penelitian dan sudah dapat ditetapkan, diharapkan kemudian akan dilanjutkan dengan penetapan wilayah pengembangan rumpun atau galur kambing lokal tertentu. Dengan begitu sumberdaya genetik kambing lokal tersebut dapat dipertahankan dan dikembangkan secara berkelanjutkan untuk meningkatkan pendapatan petani dan ikut mendukung ketahanan pangan di daerah-daerah pedesaan. Hal ini mengacu kepada Undang-Undang Republik Indonesia Nomor 18 tahun 2009, tentang Peternakan dan Kesehatan Hewan yang telah mengakomodir kekuatiran pemerintah akan terjadinya penggerusan materi genetik ternak lokal dengan melaksanakan program pewilayahan yang 78 dibagi atas: a) wilayah sumber bibit, yang bersifat mengembangkan secara murni, dengan mempertimbangkan jenis ternak dan rumpun, agroklimat, kepadatan penduduk, sosial ekonomi, budaya serta ilmu pengetahuan dan teknologi, b) wilayah produksi, pengembangbiakan dengan tujuan komersil yang memungkinkan menggunakan teknik-teknik perkawinan silang dan penggemukan, dan c) wilayah konservasi, dengan melakukan penangkaran hewan/ternak asli yang masih ada atau mengembangbiakkan hasil dari suatu wilayah sumber bibit (Departemen Pertanian 2006). Di Indonesia umumnya kambing lokal mempunyai potensi reproduksi yang sangat baik, maka introduksi melalui persilangan merupakan salah satu cara untuk menggabungkan sifat adaptabilitas dengan keunggulan genetik pertumbuhan rumpun kambing yang diintroduksi. Alternatif lain adalah dengan melakukan program seleksi untuk pengembangan ternak kambing lokal yang dianggap cukup potensial untuk menjadi bibit unggul yang berbasis rumpun kambing lokal, seperti galur Kambing PE Kaligesing yang sudah ditetapkan oleh pemerintah. Berdasarkan jumlah sampel selama pengamatan di lapangan jumlah data pejantan sangat terbatas dan sulit dijumpai di daerah pedesaan. Diduga hal ini terjadi karena peternak lebih sering menjual kambing jantan karena harga jual yang lebih tinggi dan kurang memperhatikan kualitas pejantan yang akan digunakan. Hal tersebut akan menyebabkan erosi genetik dan penurunan kualitas genetik ternak. Pada umumnya pengetahuan tentang jenis rumpun dan tingkat kesadaran menjaga kelestarian ternak kambing lokal masih sangat terbatas, sehingga perlu dilakukan sosialisasi dan penyuluhan tentang perlunya mengenal dan mempertahankan sumberdaya genetik kambing lokal Indonesia. SIMPULAN UMUM DAN SARAN Simpulan 1. Analisis morfometrik dan fenotipik keenam sub populasi kambing lokal yang diamati antara setiap sub populasi berbeda nyata terhadap sub populasi kambing lokal lainnya. 2. Faktor peubah pembeda antara kambing lokal Indonesia yang diamati berdasarkan ukuran morfometrik antara lain: lingkar kanon, lebar pinggul, lebar telinga, lebar ekor, panjang badan, tinggi pundak, tinggi tengkorak, lebar tengkorak, bobot badan, lingkar dada, lebar dada dan dalam dada. 3. Warna dan pola warna tubuh Kambing Benggala dominan tunggal warna hitam, Kambing Samosir dominan tunggal warna putih. Kambing Muara, Kambing Marica, Kambing Kacang dan Kambing Jawarandu sangat bervariasi. 4. Rataan jarak genetik berdasarkan nukleotida daerah D-loop DNA mitokondria antara 6 sub populasi kambing yang diamati adalah 0.014±0.002, hal ini menunjukkan keragaman yang sangat tinggi dan berbeda nyata antara setiap sub populasi terhadap sub populasi kambing lokal lainnya. 5. Ditemukan 50 situs nukleotida D-loop DNA mitokondria yang polimorfik dan 19 haplotip unik yang bisa digunakan sebagai penciri khusus keragaman genetik pada kambing lokal Indonesia. 6. Analisis jarak genetik sekuen nukleotida D-loop DNA mitokondria dengan sekuen dari GenBank menunjukkan bahwa diduga asal usul secara maternal dari 6 kambing lokal yang diamati termasuk dalam kelompok utama (haplogroup) lineage B. 7. Rataan jarak genetik berdasarkan nukleotida kromosom Y segmen gen SRY antar 6 sub populasi kambing lokal adalah 0.004±0.002 hal ini menunjukkan tingkat keragaman lebih rendah dibandingkan dengan nukleotida D-loop DNA mitokondria. Dimana terdapat 6 situs gen SRY yang bersifat polimorfik dan 4 haplotip unik, sehingga berdasarkan hasil analisis ini diduga asal-usul secara paternal dari 6 sub populasi kambing lokal yang diamati dapat digolongkan menjadi 4 kelompok yaitu: Kambing Kacang dan Jawarandu (1 haplotip), Kambing Marica dan Samosir (1 80 haplotip), sedangkan Kambing Muara dan Benggala membentuk haplotip sendiri-sendiri. 8. Analisis fenotipik dan analisis genetik berdasarkan nukleotida daerah Dloop DNA mitokondria serta DNA kromosom Y segmen gen SRY menunjukkan bahwa 6 sub populasi kambing lokal yang diamati dapat digolongkan menjadi 6 rumpun kambing yaitu: Kambing Kacang, Kambing Jawarandu, Kambing Marica, Kambing Samosir, Kambing Muara dan Kambing Benggala. 9. Identifikasi keragaman gen GDF9 ruas promotor pada kelompok induk yang beranak kembar dan beranak tunggal bersifat polimorfik yang mengekpresikan adanya hubungan mutasi gen GDF9 dengan sifat prolifik pada Kambing Kacang dan PE. Gen GDF dapat dijadikan sebagai alternatif untuk mengidentifikasi calon induk yang prolifik pada Kambing Kacang dan PE dalam program pemuliaan pada perusahaan pembibitan kambing. Tetapi keragaman gen GDF9 pada induk Kambing Muara dan Samosir bersifat monomorfik. 10. Hasil karakteristik kualitatif, kuantitatif, pemetaan genetik berdasarkan daerah D-loop DNA mitokondria, DNA kromosom Y segmen gen SRY yang diperoleh dalam penelitian ini dapat dipertimbangkan sebagai salah satu bahan dalam perencanaan penelitian program pengembangan ternak lokal lebih lanjut serta dalam rangka penyusunan standarisasi 6 kambing lokal Indonesia di tingkat lapangan atau peternak. 11. Alternatif strategi pemanfaatan kambing lokal Indonesia secara berkelanjutan dapat dilakukan dengan upaya antara lain: 1) penerapan di lapangan oleh para stake holder sesuai dengan UU No. 18 tahun 2009 dengan melaksanakan pewilayahan pengembangan ternak menjadi 3 wilayah yaitu wilayah sumber bibit, wilayah produksi dan wilayah konservasi, 2) peningkatan mutu genetik rumpun kambing lokal melalui kegiatan pemuliaan atau seleksi atau penyediaan semen beku untuk perkawinan dengan inseminasi buatan, 3) penyediaan pejantan unggul rumpun kambing lokal bagi petani di pedesaan, 4) sosialisasi pengenalan rumpun kambing lokal kepada peternak, penyuluh lapangan dan para pengambil kebijakan di daerah habitat asli rumpun kambing lokal, 5) perlu program pengembangan kawasan pusat pembibitan ternak (Village 81 breeding centre) atau kampung ternak berbasis rumpun kambing lokal terutama di daerah habitat aslinya. Saran 1. Tujuan penetapan kambing lokal Indonesia perlu dilanjutkan melalui penelitian karakterisasi produksi sesuai dengan kondisi peternak di pedesaan dan usaha ternak secara intensip pada Kambing Marica, Muara, Benggala dan Samosir. 2. Perintisan data base sumberdaya genetik plasma nuftah kambing lokal Indonesia perlu dilanjutkan melalui karakterisasi kambing-kambing lokal lainnya. 3. Berdasarkan pengamatan di lapangan kondisi dan kualitas pejantan relatip rendah di pedesaan, karena kurang mengetahui dan kurang mendapatkan perhatian dari peternak di pedesaan. Hal ini bisa menjadi salah satu penyebab erosi genetik kambing lokal, sehingga perlu upaya untuk menyediakan pejantan unggul kambing lokal atau penyediaan semen beku. 4. Perlu penelitian lebih lanjut untuk mengidentifikasi keragaman gen fekunditas jenis lain yang berhubungan dengan sifat prolifik pada kambing lokal Indonesia. DAFTAR PUSTAKA Abdullah MNA. 2008. Karakterisasi Genetik Sapi Aceh Menggunakan Analisis Keragaman Fenotipik daerah D-loop DNA Mitokondria dan DNA Mikrosatelit (disertasi). Bogor: Pascasarjana Institut Pertanian Bogor. Anderson L. 2001. Genetic dissection of phenotypic diversity in farm animals. Nature Rev 2:130-1138. Barker JSF, Tan SG, Moore SS, Mukherjee TK, Matheson JL, Selvaraj OS. 2001. Genetic variation within among populations of Asian goats (Capra hircus). J Anim Breed Genet 118:213-233. Batubara A, Doloksaribu M, Tiesnamurti B. 2007. Potensi keragaman sumberdaya genetik kambing lokal Indonesia. Prosiding Lokakarya Nasional Pengelolaan dan Perlindungan Sumberdaya Genetik di Indonesia. Bogor, 20 Desember 2006. Jakarta: Direktorat Kerjasama Multilateral Departemen Perdagangan, Kementerian Lingkungan Hidup, Pusat Penelitian dan Pengembangan Peternakan Departemen Pertanian, Direktorat Kerjasama dan Pengembangan Departemen Hukum dan HAM dan Perhimpunan Ilmu Pemuliaan Indonesia (PERIPI).ISBN 978-9798308-66-6.halaman 245-265. Batubara A, Pamungkas FA, Doloksaribu M, Sihite E. 2007. Potensi beberapa plasma nutfah kambing lokal Indonesia. Loka Penelitian Kambing Potong. Bogor: Pusat Penelitian dan Pengembangan Peternakan. Beja-pereira A, England PR, Ferrand N, Jordan S, Bkhiet AO, Abdalla MA, Makhour M, Jordana J, Teberlet P, Luikart G. 2004. African origin of the domestic donkey. Science 304:1781. Bodensteiner KJ, Clay CM, Moeller CL, Sawyer HR. 1999. Molecular cloning of the ovine GDF-9 and expression of GDF-9 in ovine and bovine ovaries. Biol Reprod 60:381-386. Bodin L, Pasquale ED,Fabre S, Bontoux M, Monget P, Persani L, Mulsant P. 2007. A novel mutation in the bone morphogenetic protein 15 gene causing defective protein secretion is associated with both increased ovuation rate and sterility in Laucane sheep. Endocrinol 148:393-400. Bourdon, RM. 2000. Understanding Animal Breeding. 2nd Ed. New Jersey: Prentice-Hall, USA. Bruford MW, Bradley DG, Luikart G. 2003. DNA markers reveal the complexity of livestock domestication. Natl Revol Genet 4:900-910. Budiharjo, K, Setiadi A. 2003. Analisis Usaha Ternak Kambing pada dua skala pemeliharaan ternak di Kota Semarang. Fakultas Peternakan, Universitas Diponegoro, Semarang. Canon J. 2006. Geographical partitioning of goat diversity in Europe and the Middle East. Anim Genet 37:327-334. Cardellino RA. 2006. Status of the worlds livestock genetic resources : preparation of the first report on state of the world animal genetic resources, In: (Ed: Ruane J & Sounino A) The Role of Biotechnology in Exploring and Protecting Agriculture Genetic Resources. Rome: FAO (Food and Agricultural Organization) of the United Nations. p. 3-9 83 Cardellino RA, Boyazoglu J. 2009. Research opportunities in the field of animal genetic resources. Livest Sci 120:166-173. [CBD] Convention of Biological Diversity. 2008. Conference of the Parties 9, Bonn, Germany. (http://www.cbd.int/cop9). Chen SY, Su YH, Wu SF, Sha T, Zhang YP. 2005. Mitochondrial diversity and phylogeographic structure of Chinese dosmestic goats. Mol Phylogen Evol 37:804-814. Christensen K. 2009. Population genetics. Division of Animal Genetics, Departement of Animal Science and Health. Copenhagen: The Royal Veterinary & Agriculture University, Denmark. Chu MX, Liu ZH, Jiao CL, He YQ, Fang L, Ye SC, Chen GH, Wang JY. 2006. Mutation in BMPR1B and BMP15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J Anim Sci 85:598-603. Chu MX, Jiao CL, He YQ, Wang JY, Liu ZH, Chen GH. 2007. Association between PCR-SSCP of bone morphogenetic protein 15 genes and prolificacy in Jining Grey goats. Anim Biotechnol 18:263-74. Chu MX, Wu ZH, Feng T, Cao GL, Fang L, Di R, Huang DW, Li XW, Li N. 2011. Polymorphism of GDF9 gene and its association with litter size in goats. Vet Res Commun doi 10.1007/s11259-011-9476-8. Cohen MM, Shaw MW. 1965. Two sibling with gonadal dysgenesis and a female phenotype. New Engl J Med 272:1083-1088. Crepaldi P, Negrini R, Milanesi E, Gorni C, Cicogna M, Ajmone, Marsan P. 2001. Diversity in five goat populations of the Lambardy Alps: Comparison of estimates obtained from morphometric traits and molecular markers. J Anim Breed Genet 118:173-180. Davis GH, Galloway SM, Ross IK, Gregan SM, Ward J, Nimbkar BV, Ghalsasi PM, Nimbkar C, Gray GD, Subandriyo, Inounu I, Tiesnamurti B, Martyniuk E, Eythorsdottir E, Mulsant P, Lecerf F, Hanrahan JP, Bradford GE, Wilson T. 2002. DNA tests in prolific sheep from eight countries provide new evidence on origin of the Booroola (FecB) mutation. Biol Reprod 66:1869– 1874. Davis GH. 2004. Fecundity in sheep. Anim Reprod Sci 83:247-253. Davis GH. 2005. Major gene affecting ovulation rate in sheep. Genet Select Evol 37:11-23. Davis GH, Balakrishnan L, Ross IK, Wilson T, Galloway SM, Lumsden BM, Hanrahan JP,Mullen M, Mao XZ, Wang GL, Zhao ZS, Zeng YQ,Robinson JJ, Mavrogenis P, PapachiristoforouC, Peter C, Baumung R, Cardyn P, Boujenane I, Cockett NE, Eythorsdottir E, Arranz JJ, Notter DR. 2006. Investigation of the Booroola (FecB) and Inverdale (FecX(I)) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries. Anim Reprod Sci 92: 87–96. [DEPTAN] Departemen Pertanian. 2006. Peraturan Menteri Pertanian Nomor 35/Permentan/OT.140/8/2006 tentang Pedoman Pelestarian dan Pemanfaatan Sumberdaya Genetik Ternak. Jakarta: Deptan. 84 [DEPTAN] Departemen Pertanian. 2009. Undang-Undang Republik Indonesia Nomor 18 tahun 2009 tentang Peternakan dan Kesehatan Hewan. Departemen Pertanian. Jakarta: Deptan. [DEPTAN] Departemen Pertanian. 2010. Surat Keputusan Menteri Pertanian RI Nomor 2591/Kpts/PD.400/7/2010 tanggal 19 Juli 2010 tentang Penetapan Galur Kambing Kaligesing. Departemen Pertanian. Jakarta: Deptan [DITJENNAK] Direktorat Jenderal Peternakan. 2008. Peraturan Direktur Jenderal Peternakan Nomor 07/PD.410/F/01/2008 tahun 2008 tentang Petunjuk Teknis Penjaringan Bibit Ternak. Direktorat Jenderal Peternakan. Departemen Pertanian. Jakarta: Deptan. [DITJENNAK] Direktorat Jenderal Peternakan. 2009. Statistik Peternakan. Direktorat Jenderal Peternakan. Departemen Pertanian. Jakarta: Deptan. Dodds KG, Mc Ewan JC, Davis GH. 2007. Integration of molecular and quantitative information in sheep and goat industry breeding program. Small Rum Res 70:32-41. Dong CH, Du LX, Li ZZ. 2007. Analysis of fecundity on Jining Grey goat. National centre for molecular genetic and breeding of animal. Institute of animal science (abstract). Dossa LH, Wollny C, Gauly M, 2007. Spatial variation in goat population from Benin as revealed by multivariate analysis of morphological traits. Small Rum Res 73:150-159. Du ZY, Lin JB, Qin C, Wang JF, Ran XQ. 2008. Polimorfism of exon of growth differentiation factor 9 gene in Guizhou White goats. Xumu Yu Shouyi 40:46-48. Duffy DL, Montgomery GW, Hall J, Mayne C, Healey SC, Brown J, Boomsma DI, Martin MG. 2001. Human twinning is not linked to the region of chromosome 4 syntetic with the sheep twinning gene FecB. Am J Med Genet 1100:182-186. Dwiyanto K. 2002. Kebijakan Perbibitan Nasional. Pemanfaatan Bioteknologi dalam Pengelolaan Plasma Nuftah Hewan/Ternak. Makalah pada Seminar Nasional Pemanfaatan dan Pelestarian Plasma Nuftah, Tanggal 3-4 September 2002. Bogor: Kerjasama Pusat Penelitian Bioteknologi IPB dan Komisi Nasional Plasma Nutfah, Departemen Pertanian. Everitt B, Dunn G. 2001. Applied Multivariate Data Analysis. 2nd Ed. Oxford: University Press.pp.342. Excoffier L, Laval G, Schneider S. 2006. Computational and molecular population genetics lab (CMPG). Institute of Zoology, University of Berne, Switzerland. ( http://cmpg.unibe.ch/software/arlequin3), 26 June 2011. Fan-Bin. 2007. Phylogenetic relationships among Chinese indigenous goat breeds inferred from mitochondrial control region sequence. Small Rum Res 73:262-266. [FAO] Food and Agriculture Organization. 2000. World Watch List for Domestic Animal Diversity 3rd Ed. Food and Agriculture Organization of the United Nations (FAO). Rome, Italy. [FAO] Food and Agriculture Organization. 2001. Sustainable Use of Animal Genetic Resources. IDAD-APHD FA. Rome, Italy. 85 [FAO] Food and Agriculture Organization. 2007a. The state of the World’s animal genetic resources for food and agriculture. In: Rischkowsky, B., Pilling, D. (Eds) Food and Agriculture Organization of the United Nations (FAO). Rome : Food and Agriculture Organization of the United Nations (FAO). [FAO] Food and Agriculture Organization. 2007b. The global plan of Action for animal genetic resources and Interlaken Declaration on Animal Genetic Resources. International Technical Conference on Animal Genetic Resources for Food and Agriculture, Interlaken, 3-7 September 2007. Switzerland. Rome: Food and Agriculture Organization of the United Nations (FAO). [FAO] Food and Agriculture Organization. 2008. High-Level Conference on World Food Security: the Challenges of Climate Change and Bioenergy, Rome, Italy. (http://www.fao.org/foodclimate/hlc-home/en/). Felsenstein J. 2007. Phylip (Phylogeny Inference Package) version 3.67. Washington: University of Washington. Feng T, Geng CX, Lang XZ, Chu MX, Cao GL, Di R, Fang L, Chen HQ, Liu XL, Li N. 2010. Polymorphisms of caprine GDF9 gene and their association with litter size in Jining Grey goats. Mol Biol Rep doi 10.1007/s11033-0100669-y. Princeton Univ Press Freeland JR. 2005. Molecular Ecology. London: John Wiley & Sons, Ltd. pp.388. Galloway SM, McNatty KP, Cambridge LM, Laitinen MPE, Juengel JL, Jokiranta TS, McLaren RJ, Luiro K, Dodds KG, Montgomery GW, Beattie AE, Davis GH, Ritvos O. 2000. Mutations in an oocyte derived growth factor gene (BMP15) cause increased ovulation rate and infertility in a dosagesensitive manner. Genetics 25:279–283. Garcia D, Corral N, Canon J. 2005. Combining inter- and intrapopulation information with the Weitzman approach to diversity conservation. J Hered 96:704-712. Gemmell RT, 1995. A Comparative study of the corpus luteum. Reprod Fertiliz Dev 7:303-12 Gilchrist RB, Ritter RJ, Armstrong DT. 2005. Oocyte-somatic cell interaction during follicle development in mammals. Anim Reprod Sci 82:341-377. Giuffra E, Kijas JMH, Amarger V, Carlborg O, Jeon JT, Anderson L. 2000. The origin of the domestic pig: independent domestication and subsequent introgression. Genetics 154:1785-1791. Glowatzki-Mullis ML, Muntwyler J, Baumle E, Gaillard C. 2008. Genetic diversity measures of Swiss goat breeds as decision-making support for conservation policy. Small Rum Res 74:202-211. Gootwine E, Reicher S, Rozov A. 2008. Prolificacy and lamb survival at birth in Awassi and Assaf sheep carrying the FecB (Booroola) mutation. Anim Reprod Sci 108:402–411. Guo-Hua H, Shi-Lin C, Hong-Wei Y, Wei Sheng W, Zhong S, Qi-Kang C, Lin C, Qung-Ling, Li-Guo. 2007. HaeII RFLP of INHA and its relationship to goat litter size. Heredity 29:972-976. 86 Guo-Hua H, Chen SL, Ai JT, Yang LG. 2008. None of polymorphism of ovine fecundity major genes FecB and FecX was tested in goat. Anim Reprod Sci 108:279-286.. Hafez ESE. 1993. Reproduction in Farm Animals, 6th Ed, Filadelphia: Lea and Febiger. Handiwirawan E, Subandriyo. 2004. Potensi keragaman sumberdaya genetik Sapi Bali. Wartazoa 14(3):107-115. Hanrahan JP et al. 2004. Mutation in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both Increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol Reprod 70:900909. Hardjosubroto W. 2001. Genetika Hewan. Jogjakarta: Fakultas Peternakan Universitas Gadjah Mada. Hartl, DL.1988. A Primer of Population Genetics. 2nd Ed. Massachusetts : Sinauer Associates, Sunderland, Inc. Publisher. Hasan MSR, Perwitasari RRD, Tiesnamurti B, Farajallah A. 2011. Fecundities genes (BMPR 1B and BMP15) on three Indonesia local goats. Hayati (In publications process). He YQ, Chu MX, Wang JY, Fang L, Ye SC. 2006. Polymorphism on BMPR15 as candidate gen for prolificacy in six goat breeds. J Anhui Agric Univ 33:6164. He YQ. 2010. Candidate genes polymorphism and its association to prolificacy in Chinese goats. J Agric Sci 2:1. Herera M, Rodero E, Gutierrez MJ, Pena F, Rodero JM. 1996. Application of multifactorial discriminant analysis in the morphostructural differentiation of Andalusian caprine breeds. Small Rum Res 22:39-47 Hiendleder S, Kaupe B, Wassmuth R, Janke A. 2002. Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies. Proc Rur Soc Lond Bull 269:893-904. Hoda A. 2008. Studi karakterisasi, produktivitas dan dinamika populasi Kambing Kacang (Capra hircus) untuk program pemuliaan ternak kambing di Maluku Utara. [Disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Hua GH, Chen SL, Ai JT, Yang LG. 2008. None of polymorphism of ovine fecundity major genes FecB and FecX was tested in goat. Small Rum Res 108:279-286. Hua GH, Yang LG. 2009. A review of research progress of FecB gene in Chinese breeds of sheep. Anim Reprod Sci 116:1–9. Huo JX, An XP, Wang JG, Song YX, Cui YH, Wang YF, Chen QJ, Cao BY. 2011. New genetic polymorphisms of KiSS-1 gene and their association with litter size in goats. Small Rum Res 96:106-110. Iannuzzi L, Di Meo GP, Perucatti A, Di Palo R, Zicarelli L. 2001. 50 XY gonadal dysgenesis (Swyer’s syndrome) in female river buffalo (Bubalus bubalis). Vet Rec 148:634-635. 87 Inounu I, Iniguez L, Bradford GE, Subandryo, Tiesnamurti B. 1993. Production performance of prolifik Javanese ewes. Small Rum Res 12:243-257. Joshi MB, Rout PK, Mandal AK, Tyler-Smith C, Singh L, Thangaraj K. 2004. Phylogeography and origin of Indian domestic goats. Mol Biol Evol 21:454-462. Kawakura K et al. 1996. Deletion of SRY region on the Y chromosome detected in bovine gonadal hypoplasia XY (female) by PCR. Cyto Cell Genet 72:183-184. Karna DK, Kuul GL, Bisht. 2001.Pashmina yield and its association with morphometric traits in Indian Chenghu goats. Small Rum Res 41:271-275. Kimura M. 1980. A simple method for estimating evolutionary rate of base substitution through comparative studies of nucleotida sequences. J Mol Evol 16:111-120. Kumar D. 2005. Population structure, genetic variation and management of Marwari goats. Small Rum Res 59:41-48. Kumar S, Kolte AP, Mishra AK, Arora AL, Singh VK. 2006. Identification of the FecB mutation in Garole x Malpura sheep and its effect on litter size. Small Rum Res 64:305–310. Lan XY. 2007. The novels SNPs of the IGFBP3 gene and their association with litter size and weight traits in goats. Arch Tierz Dummerst 50:223-224. Lanari MR, Taddeo H, Domingo E, Centeno MP, Gallo L. 2003. Phenotypic differentiation of exterior traits in local Criollo goat population in Patagonia (Argentina). Arch Tierz Dummerst 46:347-356. Lewin B. 2000. Genes VII. Oxford: Oxford University Press. Luikart G, Gielly L, Excoffier L, Vigne JD, Bouvet J, Teberlet P. 2001. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proc Natl Acad Sci 98:5927-5932. Mabrouk O, Sghair N, Amor G, Mohamed BA, Amel BAE. 2008. Morphostructural growth according to the sex and birth mode and relationship between body size and body weight of the local kids at the first five months of age in Tunisian arid area. Res J Biol Sci 3:120-127. MacHugh DE, Bradley DG. 2001. Livestock genetic origins: goat buck the trend. Proc Natl Acad Sci 98:5382-5384. Maddox JF, Cockett NE. 2007. An Update on sheep and goat linkage maps and other genomic resources. Small Rum Res 70:4-20. Mahmilia F, Ginting SP, Batubara A, Sianipar J, Tarigan A. 2004. Karakteristik Morfologi dan Performans Kambing Gembrong dan Kambing Kosta. Laporan Hasil Penelitian TA. 2004. Sei Putih: Loka Penelitian Kambing Potong Sungei Putih, Sumatera Utara. Mahmilia F, Tarigan A. 2004. Karakteristik morfologi dan performans Kambing Kacang, Kambing Boer dan persilangannya. Prosiding Lokakarya Nasional Kambing Potong: Kebutuhan Inovasi mendukung Agribisnis yang berdayasaing Bogor, 6 Agustus 2004. Bogor: Pusat Penelitian dan Pengembangan Peternakan, Badan Penelitian dan Pengembangan Pertanian. 88 Mailund T, Brodal GS, Fagerberg R, Pederson CNS, Phillips D. 2008. Recraffting the neighbour-joining method. MBC Bioinform7:29-36. Mannen H, Nagata Y, Tsuji S. 2001. Mitochondrial DNA reveal that domestic goat (Capra hircus) are genetically affected by two subspecies of bezoar (Capra aegagrus). Biochem Genet 39:145-154. Mansjoer SS, Kertanugraha T, Sumantri C. 2007. Estimasi jarak genetik antar domba garut tipe tangkas dengan tipe pedaging. Med Pet 30:129-138. Martinez-Royo A, Jurado JJ, Smulder JP, Marti JI, Alabart JL, Roche A, Fantova E, Bodin L, Mulsant P, Serrano M, Folch J, Calvo JH. 2008. A deletion in the bone morphogenetic protein 15 gene causes sterility and increased prolificacy in Agaronesa sheep. Anim Genet 39:294-301. Mattjik AA, Sumertajaya M. 2006. Perancangan Percobaan dengan Aplikasi SAS dan MINITAB. Institut Pertanian Bogor, Bogor:IPB. Mburu D, Hanotte O. 2005. A practical approach to microsatellite genotyping with special reference to livestock population genetics. Nairobi: International Livestock Research Institute, Nairobi, Kenya. McElreavey K. 1996. Mechanism of sex determination in mammals. Adv Gen Bio 4:304-354. McNatty KP, Smith P, Moore LG, Reader K, Lun S, Hanrahan JP, Groome NP, Laitinen M, Ritvos C, Juengel JL. 2005. Oocyte expressed genes affecting ovulation rate. Mol Cell Endocr 234:57–66. Merkens J, Sjarif A. 1932. Bijdrage tot de kennis van de geitenfokkerij in Nederlandsch Oost Indie. Nederland Bladen voor Diergenekuun 44:36466. (Terjemahan Bahasa Indonesia: Sumbangan pengetahuan tentang peternakan kambing di Indonesia. Dalam: Domba dan Kambing. Terjemahan Karangan mengenai Domba dan Kambing di Indonesia, September 1979, SDE-67). Moioli B, Napolitano F, Orru L, Catillo G. 2006. Analysis of the genetic diversity between Gentile di Puglia, Saprarissana and Sarda sheep breed using microsatellite markers. Ital J Anim Sci 5:73-78. Montaldo HH. 2006. Genetic engineering applications in animal breeding. E J Biotechnol 9:159-170. Moore RK, Otsuka F, Shimasaki S. 2003. Molecular basic of bone morphogenetic protein-15 signaling in granulosa cells. J Biol Chem 278:304-310. Muladno 2006. Aplikasi Teknologi Molekuler dalam Upaya Peningkatan Produktivitas Hewan. Pelatihan Teknik Diagnostik Molekuler untuk Peningkatan Produksi Peternakan dan Perikanan di Kawasan Timur Indonesia. Bogor: Kerjasama Pusat Studi Ilmu Hayati, Lembaga Penelitian dan Pemberdayaan Masyarakat Institut Pertanian Bogor dan Direktorat Jenderal Pendidikan Tinggi Depdiknas. Mulsant P, Lecerf F, Fabre S, Schibler L, Monget P, Lanneluc I, Pisselet C, Riquet J, Monniaux D, Callebaut I, Cribiu E, Thimonier J, Teyssier J, Bodin L, Cognie Y, Chitour N, Elsen JM. 2001. Mutation in bone morphogenetic protein receptor-1B is associated with increased ovulation rate in Booroola Merino ewes. Proc Nat Acad Sci USA 98:5104–5109. 89 Naderi S, Rezaei HR, Taberlet P, Zundel S, Rafat SA, Nagasha HR, El-Barody MAA, Ertugrul O, Pompanon F. 2007. Large-scale mitochondrial DNA analysis of domestic goat reveal six haplogroup with high diversity. PloS ONE 2, e1012. Doi: 10. 1371/journal.pone.0001012. Nei M. 1987. Molecular Evolutionary Genetics. New York: Columbia University Press. Nijman IJ. 2003. Hybridization of Banteng (Bos javanicus) and Zebu (Bos indicus) revealed by mitochondria DNA, satellite DNA, AFLP and microsatellites. Heredity 90:10-16. Noor RR.2002. Gen Kelenturan Fenotipik Ubah Strategi Peternakan. URL Source(http://www.kompas.co.id/kompascetak/0312/15/inspirasi/743375. htm), 25 Mei 2011. Noor RR. 2008. Genetika Ternak. Ed ke-4. Jakarta: Penebar Swadaya, pp.200. Nsoso SJ, Podisi B, Otsogile E, Mokhutshwane BS, Ahmadu B. 2004. Phenotypic characterization of indigenous Tswana goats and sheep breeds in Botswana: Continuous traits. Trop Anim Health Prod 36:789800. Odubute IK, Akinokun JO, Ademosum AA. 1992. Production characteristic of West African Dwarf goats under improved management system in the humid tropics of Nigeria. Proc International Workshop, Ile-Ile, Nigeria: ILRI pp.202-207. Parma P, Feligini M, Greppi G, Enne G. 2003. The complete nucletide sequence of goat (Capra hircus) mitochondrial genome. DNA Seq 14:199-203. Parma P, Feligini M, Greppi G, Enne G. 2004. The complet coding region of river buffalo (Bubalus bubalis) SRY gene. DNA Seq 15:77-80. Pereira B. 2008. A multiplex primer extension assay for the rapid identification of paternal lineages in domestic goat (Capra hircus): Laying the foundations for a detailed caprine Y chromosome phylogeny. Mol Phylogenet Evol 49:663-668. Polley S, Dea S, Batabyalb S, Kaushika R, Yadava P, Aroraa JS, Chattopadhyaayb S, Panc S, Brahmad B, Dattaa TK, Goswami SL. 2009. Polymorphism of fecundity genes (BMPR 1B, BMP15 and GDF9) in the Indian prolific Black Bengal goat. Small Rum Res 85:122-129 Ponzoni RW. 1997. Genetic Resources and Conservation. New York: CAB International. Power MM. 1986. XY sex reversal in mare. Equine Vet J 18:233-236. Prashant, Gour DS, Dubey PP, Jain A, Gupta SC, Joshi BK, Kumar D. 2008. Sex determination in 6 Bovid species by duplex PCR. J Appl Genet 49:379381. Prashant, Gour DS, Dubey PP, Jain A, Nanda DK, Joshi BK, Kumar D. 2009. Complete nucleotide sequencing, SNP identification and characterization of SRY gene in Indian Sangamneri goat. Afric J Biotechnol 8:2939-2942. [PUSLITBANGNAK] Pusat Penelitian dan Pengembangan Peternakan. 2007. Penetapan dan pengakuan rumpun dan galur ternak mendukung sistem perbibitan ternak nasional yang berdaya saing dan berkelanjutan. Pusat 90 Penelitian dan Pengembangan Peternakan, Badan Penelitian dan Pengembangan Pertanian, Bogor: Puslitbangnak. Quinn GP, Keough MJ. 2002. Experimental Design and Data Analysis for Biogist. Cambridge: Cambridge University Press, pp.537. Ren ZZ, Cai HF, Luo WX, Liu RY. 2010. GDF9 and BMP15 genes analysis of genetic variation on Qianbei-pockmarked goats. China Anim Husband Vet Med 7: 2010-2017 Riva J, Rizzi R, Marelli S, Cavalchini LG. 2004. Body measurements in Bergamasca sheep. Small Rum Ress 55:221-227. Rout PK, Joshi MB, Mandal A, Laloe D, Singh L, Tangaraj K. 2008. Microsatellit based phylogeni of Indonesian domestic goats. Bio Medic Cent Genet 9:1-11. Royo LJ, Traore A, Tamboura HH, Alvarez I, Kabore A, Fernandez I, Sanou GO, Toguyeni A, Sawadogo L, Goyache F. 2009. Analysis of mitochondrial DNA diversity in Burkina Faso populations confirms the maternal genetic homogenity of the West African Goat. Anim Genet 40:344-347. Ruane J. 2000. A framework for prioritizing domestic animal breeds for conservation purpose at the national level: a Norwegian case study. Conservat Biol 14:1385-1393. Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning, A Laboratory Manual 3rd Ed. NY: Cold Spring Harbor Laboratory Press. Sambrook, J. and D.W. Russell. 2001. Molecular Cloning, A Laboratory Manual. 2nd Ed. NY: Cold Spring Harbor Laboratory Press. SAS Institute. 2005. SAS/STAT Guide for Personal Computer. Version 9.1 Ed. NC: SAS Institute Cary. USA. Savolainen P, Zhang YP, Luo J, Lunderberg J, Leitner T. 2002. Genetic evidence for an East Asian origin of domestic dogs. Science 298:1610-1613. Sechi T, Miari S, Pirumpun D, Crumpunta L, Mulas G, Carta A. 2009. Genetic variation of goat Y chromosome in the Sardinian population. Ital J Anim Sci 8:159-161. Seli E, Sakkas D. 2005. Spermatozoa nuclear determinants of reproductive outcome: implication for ART. Hum Reprod Update 11:337-349. Setiadi B, Subandriyo, Iniguez LC. 1995. Reproductive performance of small ruminants in an outreach pilot project in West Java. JITV 1:73-80. Setiadi B, Prianto D, Martawidjaja. 1997. Komparatif Morpologi Kambing. Laporan Hasil Penelitian APBN 1996/1997. Bogor: Balai Penelitian Ternak Ciawi-Bogor. Setiadi B, Mathius IW, Sutama KI. 1998. Karakteristik Sumberdaya Kambing Gembrong dan Alternatif Pola Konservasinya. Prosiding : Seminar Nasional Peternakan dan Veteriner, Bogor, Oktober; 1998. Bogor: Pusat Penelitian dan Pengembangan Peternakan. Setiadi B, Priyanto D, Subandriyo. 1999. Karakteristik Morfologi dan Produktivitas Induk kambing Peranakan Etawah di Daerah Sumber Bibit Kabupaten Purworejo. Prosiding Seminar Nasional Kiat Usaha 91 Peternakan, Purwokerto, Agustus 1999. Jember: Universitas Jenderal Sudirman. Setiadi B, Tiessnamurti B, Subandriyo, Sartika T, Adiati U, Yulistiani D, Sendow I. 2002. Koleksi dan Evaluasi Karakteristik Kambing Kosta dan Gembrong Secara Ex-situ. Laporan Hasil Penelitian APBN 2001/2002. Bogor: Balai Penelitian Ternak Ciawi-Bogor. Shrestha JNB, Fahmy MH. 2007. Breeding goats for meat production. 3. Selection and breeding strategies. Small Rum Res 67:113-125. Simianer H. 2006. Use of Molecular marker and other information for sampling germ plasm to create an animal gene bank. In: Ruane J, Sonnino A. (Edit) The Role of Biotechnology in Exploring and Protecting Agricultural Genetic Resources. Rome: Food and Agriculture Organization of the United Nations (FAO). Sinclair AH, Berta P, Palmer MS, Hawkins JR, Griffiths BL, Smith MJ,Foster JW,Frischauf AM, Lovell-Badgeand RP, Goodfellow N. 1990. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 346:240-244 Sodiq A, Taufik ES. 2003. The role and breeds, management system, productivity and development strategies of goats in Indonesia: A Review. J Agric Rur Dev Trop 104:71-89. Soedjana TD. 2008. Goat Development in Indonesia. In: International Seminar on Dairy and Meat goat production, Bogor August 5-6, 2008. Bogor: Indonesian Research Institute for Animal Production. pp:154-157. Solis A, Jugo BM, Meriaux JC, Iriondo M, Mazoke LI, Aquirre AI, Vicario A, Estomba A.2005. Genetic diversity within and among four South European native horse breed based on microsatellite DNA analysis: implication for conservation. J Hered 96:670-678. Souza CJH, MacDougall C, Campbell BK, McNeilly AS, Baird DT. 2001. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1B (BMPR1B) gene. J Endocr 169:1–6. Sponenberg P. 2004. Genetics of goat color. Blacksburg: Virginia-Maryland Regional College of Veterinary Medicine, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061 USA. (http:www.cagba.org/ genetics goat clr 20041.pdf) Steel RGD, Torrie JH. 1985. Prinsip dan Prosedur Statistika: Suatu Pendekatan Biometrik. (Terjemahan: Sumantri B). Edisi kedua. Jakarta : PT. Gramedia. Subandriyo, Setiadi B. 2003. Pengelolaan Plasma Nutfah Hewani sebagai Aset dalam Pemenuhan Kebutuhan Manusia. Makalah disampaikan dalam Lokakarya Pemantapan Pengelolaan Database dan Pengenalan Jejaring Kerja Plasma Nutfah Pertanian. Bogor, 21-28 Juli 2003. Bogor: Komisi Nasional Plasma Nutfah. Subandriyo. 2004. Strategi Pemanfaatan Plasma Nutfah Kambing Lokal dan Peningkatan Mutu Genetik Kambing di Indonesia. Lokakarya Nasional Kambing Potong, Bogor 6 Agustus 2004. Bogor: Pusat Penelitian dan Pengembangan Peternakan, Badan Penelitian dan Pengembangan Pertanian. 92 Subandriyo. 2008. Goat breeding and genetic resources in Indonesia. In Proceedings: International Seminar on Dairy and Meat goat production, Bogor August 5-6, 2008. Bogor: Indonesian Research Institute for Animal Production. p:29-37. Sultana S, Mannen H, Tsuji S. 2003. Mitochondrial DNA diversity of Pakistani goats. Anim Genet 34:417-421. Sumantri C, Einstiana A, Salamena JF, Inounu I. 2007. Keragaman dan hubungan phylogenik antar domba lokal di Indonesia melalui pendekatan analisis morfologi. JITV 12:42-54. Suparyanto A, Purwadaria T, Subandriyo. 1999. Pendugaan jarak genetik dan faktor peubah pembeda bangsa dan kelompok domba di Indonesia melalui pendekatan analisis morfologi. JITV 4:80-87 Swedlow JR, Matteri RL, Parkoff H. 1996. Deglycosylation of gonadothropine with an endoglycosidase. Proc Soc Exp Biol Med 18: 432-437. Tadelle DA. 2003. Phenotypic and genetic characterization of local chicken ecotypesin Ethiopia. PhD thesis. Berlin: University of Humboldt – Berlin, Germany. Takebayashi K, Takakura K, Wang HQ, Kimura F, Fasahara K, Noda Y. 2000. Mutation analysis of the growth differentiation factor 9 and 9B genes in patients with premature ovarian failure and polycustics ovary syndrome. Fertil Steril 74:976-979. Tamura K, Dudley J, Nei M, Kumar S. 2007. Molecular evolutionary genetic analysis (MEGA) software version 4. Mol Biol Evol 24:1596-1599. Tegelstrom H. 1986. Mitochondrial DNA in natural population : An inproved routine for the screening of genetic variation based on sensitive silver staining. Electrophoresis 7:226-229. Tozaki T, Kakoi H, Mashima S, Hirota K, Hasegawa T, Ishida N, Miura N, ChoiMiura NH, Tomita M. 2001. Population study and validation of paternity testing for thoroughbreed horses by 15 microsatellite loci. J Vet Med 63:1191-1197. Traore A et al. 2008. Multivariates analysis on morphological traits of goats in Burkina Faso. Arch Tierz Dummerst 6:588-600. Troy CS, MacHugh DE, Bailey JF, Magee DA, Loftus RT, Cuningham P, Chamberlain AT,Sykes BC, Bradley DG. 2001. Genetic evidence for near-eastern origins of European Cattle. Nature 410:1088-1091. Utoyo DP. 2002. Management of the farm domestic animal genetic resources in Indonesia. In : Animal Genetic Resources. Directorate General of Livestock Service. Jakarta: Ministry of Agriculture Indonesia. Veiteia RA, Salas-Cortex L, Ottolenghi C, Pailhoux E, Cotinot C, Fellous M. 2001. Testis determination in mammals: more questions than answers. Mol Cell Endocr 179:3-16. Verkaar ELC, Vervaecke H, Roden C, Mendoza RL, Barwegen MW, Susilawati T, Nijman IJ, Lenstra JA. 2003. Paternally inherited markers in bovine hybrid populations. Hered 91: 565-569. 93 Vila C, Leonard JA, Gotherstrom A, Marklund S, Sandberg K, Liden K, Wayne RK, Ellergen H. 2001. Widespread origin of domestic horse lineages. Science 291:474-477. Visser C, Hefer CA, Van Marle-Koster E, Kotze A. 2004. Genetic variation of three commercial and three indigenous goat populations in Souh Africa. South Afric J Anim Sci 34: 24-27,Supp 1. Wandia IN. 2001. Genom Mitokondria. J Vet. 2(4) :131-137. Wilson T, Wu XY, Juengel JL, Ross IK, Lumsden JM, Lord EA, Dodds KG, Walling GA, McEwan JC, O’Connell AR, McNatty KP, Montgomery GW 2001. Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetik protein 1B receptor (ALK-6) that is expressed in both oocytes and granulose cells. Biol Reprod 64:1225– 1235. Wimmers K, Ponsuksili S, Hardge T, Valle-Zarate A, Marthur PK, Horst P. 2000. Genetic distinctnes of African, Asian and South American local chickens. Anim Genet 31: 159-165. Womack JE. 2005. Advance in livestock genomics: Opening the barn door. Gen Res 15:1699-1705. Wu ZH, Chu MX, Li XW, Fang L, Ye SC, Liu ZH, Chen GH. 2010. PCR-SSCP analysis on exon 2 of GDF9 gene in goats. Shincuan: College of animal science and technology, Sichuan Agricultural University. Xu-qin R, Jian-bin L, Zhi-yong D, Cheng Q, Jia-fu W. 2009. Diversity of BMP15 and GDF9 genes in white goat of Guizhou Province and evolution of the encoded proteins. CN 53-1040/Q. Beijing: The National Science Foundation of China. Yan C, Wang P, DeMayo J, DeMayo FJ, Elvin J, Carino C, Prumpunad SV, Skinner SS, Dunbar BS, Dube JL, Celeste AJ, Matzuk MM. 2001. Synergistic role of bone mophogenetic protein 15 and growth differentiation factor 9 in ovarian function. Mol Endocr 15:854-866. Zeder MA, Hesse B. 2000. The initial domestication of goats (Capra hircus) in the Zagros Mountain 10,000 years ago. Science 287:2254-2257. Zhang CY, Wu CJ, Zeng WB, Huang KK, Li X, Feng JH, Wang D, Hua GH, Xu DQ, Wen QY, Yang LG. 2011. Polymorphism in exon 3 of follicle stimuating hormon beta (FSHB) sub unit gene and its association with litter traits and superovulation in the goat. Small Rum Res 96:53-57. Zhang GX, Chu MX, Wang JY, Fang L, Ye SC. 2007. Polymorphism of exon 10 of prolactin receptor gene and its relationship with prolificacy of Jining Grey Goats. Yi Chuan 29:329-36. Abstr (Article in Chinese) Zhang X, Leung FC, Chan DK, Chen Y, Wu C. 2002. Comparative analysis of allozyme, random amplified polymorphic DNA, and microsatellite polymorphism on Chinese native chickens. Poult Sci 81: 1093–1098. Zhao Y, Zhang J, Zhao E, Zhang X, Liu X, Zhang N. 2011. Mitochondrial DNA diversity and origins of domestic goats in Southwest China (excluding Tibet). Small Rum Res 95:40-47. 94 Lampiran 1 Pola warna dominan dan warna belang 6 sub populasi kambing lokal a. Kambing Jawarandu 95 b. Kambing Kacang 96 c. Kambing Samosir 97 d. Kambing Marica 98 e. Kambing Benggala 99 f. Kambing Muara 100 Lampiran 2 Form daftar karakter morfologi yang diukur DAFTAR KARAKTERISTIK MORFOLOGIK KAMBING ................... DI PROPINSI .................................... No:.................................... No. URAIAN 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 Panjang badan (cm) Lingkar dada (cm) Lingkar pinggul (cm) Lingkar canon (cm) Tinggi canon (cm) Panjang tanduk (cm) Orientasi tanduk Panjang telinga/Tipe telinga Skor rahang Panjang tengkorak (cm) Lebar tengkorak (cm) Tinggi tengkorak (cm) Gigi/umur (pasang) Jenis kelamin Paritas (untuk betina) Tipe lahir/ Tipe sapih No induk/ no bapak Status fisiologik Tinggi pundak (cm) Dalam dada (cm) Tinggi pinggul (cm) Dalam pinggul (cm) Lebar tungging (cm) Panjang ekor (cm) Tebal/ lebar ekor (cm) Warna tubuh dominan/ warna belang/ warna kepala dominan. Pola warna tubuh Penyebaran belang (%) Garis muka/ Garis punggung Ambing/ puting Bobot ternak (kg) LOKASI :........................................ NOMER TERNAK (SAMPEL) 1 2 3 4 5 101 Lampiran 3 Lokasi pengambilan data dan sampel darah kambing lokal (Kambing Kacang, Samosir, Marica, Jawarandu, Muara dan Benggala) No. Sub populasi/ Kecamatan Desa Provinsi/Kabupaten 1. Kambing Benggala, Provinsi Nusa Tenggara Timur a. Kab. Kupang Kupang Timur Oelatimo, Tuapukan, Naibonat Kupang Barat Sumlili, Tablolong Aesesa Mbay, Ngolonio b. Kab. Ende Kelimutu Kuru Lio Timur Habatua, Detupera c. Sikka Alok Kota Uneng, Nanglimang 2. 3. 4. 5. Kambing Marica, Provinsi Sulawesi Selatan a. Maros Tanralili Bantimurung b. Jeneponto Arungkeke Kelara Bangkala Kambing Jawarandu, Provinsi Jawa Tengah a. Blora Kec. Kradenan Kec. Sambong Kec. Kedung Tuban Kec. Randu Blatung Mendenrejo, Sumber Sambongrejo Ngelandeyan, Tanjung, Bajo Pilang, Randu Blatung Kambing Muara, Provinsi Sumatera Utara a. Kabupaten Tapanuli Kec. Muara Utara Batu Binumbun, Simatupang, Dolok Martumbur, Huta Lontung Kambing Samosir, Provinsi Sumatera Utara a. Kabupaten Samosir Kec. Pangururan Kec. Harian Kec. Simanindo Kec. Onan Runggu Kec. Sianjur Mulamula 6. Borong, Toddo Pulia, Damai Leang-leang, Mattoangin Arung keke, Kampala Bontolebang, Tolo Tombo-tombolo Kalimporo Kambing Kacang, Provinsi Sumatera Utara a. Kabupaten Deli Kec. Galang Serdang Kec. Bangun Purba Kec. Dolok Masihul Lumban Suhisuhi, Pardugul, Saitni huta Turpuk Limbong, Partungko Naginjang, Siparmahan Simarmata, Ambarita Sitamiang, Sitinjak Sianjur Mulamula, Hutaginjang Kotangan, Tanjung Gusti, Paya Sampir, dan Kandang Percobaan Loka Penelitian Kambing Sei Putih Bandar Kuala Bangun Purba Dolok Masihul, Paya Buaya 102 Lampiran 4 Nama/bangsa kambing, nomor akses dan asal sampel yang digunakan dari GenBank untuk membentuk pohon filogeni No. Ruas DNA Rumpun/galur Nomor Lokalitas kambing di GenBank Akses GenBank 1 D-LOOP Capra hircus AF533441 Vietnam Jamnapari AY155816 India Black bengal AY155721 India Alpine EF617779 Prancis Iranian Goat EF617945 Iran Laos native AB044303 Laos Azeri EF617706 Azerbaijan Mongolian Goat AJ317833 Cina Matou DQ121578 Cina Barbari AY155708 India Switzerland Goat AJ317838 Swiss Spanish Goat EF618413 Spanyol Taihang DQ188893 Cina Pashmina AY155952 India Pinqau EF617701 Austria Girgentara DQ241349 Sicilia Girgentara DQ241351 Sicilia Nubian FJ571542 Italia Gurcu EF618535 Turki Baladi EF617727 Egyp Boer GQ141235 Afrika Selatan Banjiao DQ121491 Cina Angora GQ141232 Cina Saanen FJ571552 Italia Maltese FJ571532 Italia Punjab Goat AB162215 Pakistan 2 Gen SRY Capra hircus Shiba Goat EU581862 D82963 Australia Jepang 3 Gen GDF9 Capra hircus Shiba Goat EF446168 D82963 Cina Jepang Lampiran 5 Pensejajaran berganda nukleotida ruas D-loop DNA mitokondria pada kambing lokal [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 1 1234567890 AACCACTATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111112 1234567890 AACCACATCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222223 1234567890 ATTAATATAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333334 1234567890 CCCCAAAAAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444445 1234567890 ATTAAGAGCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555556 1234567890 TCCCCAGTAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666667 1234567890 TAAATTTACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777778 1234567890 AAAAATTTCA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888889 1234567890 AATATACAAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1 ] 9999999990 ] 1234567890 ] ACAAACTTCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 103 104 [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 1111111111 0000000001 1234567890 CACTCCACAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111112 1234567890 GCTTACAGAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 2222222223 1234567890 ATGCCAACAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 3333333334 1234567890 CCCACACGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 4444444445 1234567890 TAAAAACATC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 5555555556 1234567890 CCAATCCTAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 6666666667 1234567890 CCCAACTTAG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 7777777778 1234567890 ATACCCACAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 8888888889 1234567890 AAACGCCAAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111112 ] 9999999990 ] 1234567890 ] ACCACACAAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 2222222222 0000000001 1234567890 GTTACGCGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 1111111112 1234567890 TGCAAGTACA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 2222222223 1234567890 TTACACCGCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 3333333334 1234567890 CGCCTACACA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ....C..... ....C..... 2222222222 4444444445 1234567890 CAAATACATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 5555555556 1234567890 TACTAACATC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 6666666667 1234567890 CATATAACGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 7777777778 1234567890 GGACATACAG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 8888888889 1234567890 CCTTCATATA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222223 ] 9999999990 ] 1234567890 ] GTTTACTATA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 105 106 [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 3333333333 0000000001 1234567890 TATCTACCCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 1111111112 1234567890 ACACATGTGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 2222222223 1234567890 AGTACTAATC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 3333333334 1234567890 CAGCATAAAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 4444444445 1234567890 GTAATGTATG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 5555555556 1234567890 TA--CATTAC ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..--...... ..AC...... ..AC...... ..AC...... ..AC...... ..AC...... 3333333333 6666666667 1234567890 ATTTT-ATGA .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....T.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... .....-.... 3333333333 7777777778 1234567890 TCTACTTCAC .......... .......... .......... .......... .......... .........T .........T .........T .........T .........T .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 8888888889 1234567890 GTGTACGTAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333334 ] 9999999990 ] 1234567890 ] ATAATATTAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 4444444444 0000000001 1234567890 TGTAACAA-G ........-. ........-. ........-. ........-. ........-. .......G-. .......G-. .......G-. .......G-. .......G-. ........-. ........-. ........-. ........-. ........-. ....C...-. ....C...A. ........-. ....C...A. ....C...-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. 4444444444 1111111112 1234567890 GACATAATAT .......... .......... .......... .......... .......... ......G... ......G... ......G... ......G... ......G... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 2222222223 1234567890 GTATATAGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 3333333334 1234567890 CATTAAACGA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 4444444445 1234567890 TTTTCCACAT ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 5555555556 1234567890 GCATATTAAG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 6666666667 1234567890 CACGTATATC .......... .......... .......... .......... .......... ......C... ......C... ......C... ......C... ......C... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 7777777778 1234567890 AGTATTAATG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 8888888889 1234567890 TAATAAAGAC ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... ......G... .......... .......... .......... .......... .......... .......... ......G... ......G... ......G... ......G... .......... .......... .......... .......... .......... 4444444445 ] 9999999990 ] 1234567890 ] ATAATATGTA .......... .......... .......... .......... .......... ...G...... ...G...... ...G...... ...G...... ...G...... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 107 108 [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 5555555555 0000000001 1234567890 TATCGTACAT .......... .......... .......... .......... .......... ...T...... ...T...... ...T...... ...T...... ...T...... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 1111111112 1234567890 TAAACGATCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 2222222223 1234567890 CCCCCATGCA ...T-..... ...T-..... ...T-..... ...T-..... ...T-..... ...T...... ...T...... ...T...... ...T...... ...T...... ...T-..... ...T-..... ...T-..... ...T-..... ...T-..... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 3333333334 1234567890 TATAAGCACG .......... .......... .......... .......... .......... ........T. ........T. ........T. ........T. ........T. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 4444444445 1234567890 TACAATGTCC .......CTT .......CTT .......CTT .......CTT .......CTT ..T....... ..T....... ..T....... ..T....... ..T....... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 5555555556 1234567890 TTATTAGCAG C....G.... C....G.... C....G.... C....G.... C....G.... .....G.... .....G.... .....G.... .....G.... .....G.... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 6666666667 1234567890 TACATGGTAC .....A.... .....A.... .....A.... .....A.... .....A.... .....A.... .....A.... .....A.... .....A.... .....A.... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 7777777778 1234567890 ATTTTACTGT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .G.......C .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 8888888889 1234567890 ATACCCGTAC ...TT..... ...TT..... ...TT..... ...TT..... ...TT..... ...T...... ...T...CC. ...T...... ...T...... ...T...... ...T...... ...T...... ...T...... ...T...... ...T...... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555556 ] 9999999990 ] 1234567890 ] ATAGCACATA .......... .......... .......... .......... .......... ..G....... .......... ..G....... ..G....... ..G....... .......... .......... .......... .......... .......... ..G....... ..G....... ..G....... ..G....... ..G....... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 6666666666 0000000001 1234567890 AAGTCAAATC .......... .......... .......... .......... .......... G......... .......... .......... .......... G......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 1111111112 1234567890 TATCCTTGTC C..T...... C..T...... C..T...... C..T...... C..T...... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ...T...... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 2222222223 1234567890 AACATGCGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 3333333334 1234567890 TCCCGTCCAC .......... .......... .......... .......... .......... .........T .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 4444444445 1234567890 TAGATCACGA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ..A....... .......... ..A....... ..A....... ..A.....A. .......... .......... .......... .......... .......... 6666666666 5555555556 1234567890 GCTTGTCGAC .......... .......... .......... .......... .......... ......T... .......... ......T... ......T... ......T... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 6666666667 1234567890 CATGCCGCGT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ......C... .......... .......... .......... .......... .......... 6666666666 7777777778 1234567890 GAAACCAGCA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 8888888889 1234567890 ACCCGCTTGG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......A.. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666667 ] 9999999990 ] 1234567890 ] CAGGGATCCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 109 110 [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 7777777777 0000000001 1234567890 TCTTCTCGCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......C.. .......... .......... .......... .......... .......... 7777777777 1111111112 1234567890 CCGGGCCCAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ........G. .......... .......... .......... .......... .......... 7777777777 2222222223 1234567890 TAACCGTGGG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 3333333334 1234567890 GGTAGCTATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 4444444445 1234567890 TAATGAACTT .......... .......... .......... C......... .......... .......... .......... .......... .......... .......... .......T.. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 5555555556 1234567890 TATCAGACAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 6666666667 1234567890 CTGGTTCTTT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... A.....G... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 7777777778 1234567890 CTTCAGGGCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ........G. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 8888888889 1234567890 ATCTCACCTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777778 ] 9999999990 ] 1234567890 ] AAATCGCCCA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus #Marica-1 #Marica-2 #Marica-3 #Marica-4 #Marica-5 #Kacang-1 #Kacang-2 #Kacang-3 #Kacang-4 #Kacang-5 #Samosir-1 #Samosir-2 #Samosir-3 #Samosir-4 #Samosir-5 #Benggala-1 #Benggala-2 #Benggala-3 #Benggala-4 #Benggala-5 #Jawarandu-1 #Jawarandu-2 #Jawarandu-3 #Jawarandu-4 #Jawarandu-5 #Muara-1 #Muara-2 #Muara-3 #Muara-4 #Muara-5 8888888888 0000000001 1234567890 CTCTTTCCTC .....C.... .....C.... .....C.... .....C.... .....C.... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ..G....... .......... .......... .......... 8888888888 1111111112 1234567890 TTAAATAAGA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888888 2222222223 1234567890 CATCTCGATG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ........C. .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... ..C....... ..C....... ..C....... ..C....... ..C....... 8888888888 3333333334 1234567890 GACTAATGAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .........A .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888888 4444444445 1234567890 TAATCAGCCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888888 5555555556 1234567890 ATGCTCACAC .......... .......... .....A.... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888888 6666666667 1234567890 ATAACTGTGC .......... .......... .......... ....TC.... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 888888888] 777777777] 123456789] TGTCATACA ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... ......... 111 112 Lampiran 6 Pensejajaran berganda nukleotida gen SRY pada kambing lokal [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 1 1234567890 CCAGATGGAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111112 1234567890 GTAGAGACAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222223 1234567890 TGCACCCCTT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333334 1234567890 CACATACAGG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444445 1234567890 GACGATTGTG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555556 1234567890 CCAAGACCAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666667 1234567890 ACACTCACAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777778 1234567890 ATGGAAAGCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 8888888889 1234567890 AATTATGCCG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1 ] 9999999990 ] 1234567890 ] CTCACAGTCC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 1111111111 0000000001 1234567890 CTGATTCTAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111112 1234567890 CCAATTCACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 2222222223 1234567890 TCTGCAAAAG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 3333333334 1234567890 GAGCATCACA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 4444444445 1234567890 GCAGCTGGAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 5555555556 1234567890 AAACCTGGGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 6666666667 1234567890 CACGATAGGG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 7777777778 1234567890 TAACATTGGA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111111 8888888889 1234567890 TACACGGATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 1111111112 ] 9999999990 ] 1234567890 ] TCCGCGGACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 2222222222 0000000001 1234567890 TTCCCTTTTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 1111111112 1234567890 CCAAAGCTTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 2222222223 1234567890 GAGCCTGGGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 3333333334 1234567890 TTTCTTGCGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 4444444445 1234567890 TTATGTTCAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 5555555556 1234567890 TACTGACTTC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 6666666667 1234567890 CTTACTCTCG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 7777777778 1234567890 CTAACAAAGG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222222 8888888889 1234567890 CACGCTTTAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 2222222223 ] 9999999990 ] 1234567890 ] CTCAATTTTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 3333333333 0000000001 1234567890 CTACAATTTC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 1111111112 1234567890 ACCTCCGACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 2222222223 1234567890 TAATTTTAAA .........T .........T .........T .......... .......... .......... .......... .......... .........T .........T .........T .........T .........T .......... 3333333333 3333333334 1234567890 CTAAGGCAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 4444444445 1234567890 TAAGTACATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 5555555556 1234567890 TAACAAGTAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 6666666667 1234567890 AGAATTTG-G ........T. ........T. ........T. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. ........-. 3333333333 7777777778 1234567890 ACTTTCCAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333333 8888888889 1234567890 ATAATTGCTC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 3333333334 ] 9999999990 ] 1234567890 ] CTCTGTTGAT .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......A.. .......... 113 114 [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 4444444444 0000000001 1234567890 CAGTTCTTTC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 1111111112 1234567890 TGTAAGAGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 2222222223 1234567890 TTTTTTGTAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 3333333334 1234567890 GAAATTATCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 4444444445 1234567890 TAACAGCACC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 5555555556 1234567890 AAAACTGCTT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 6666666667 1234567890 GAGTTAGATC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 7777777778 1234567890 ATCTGTTTTT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444444 8888888889 1234567890 CCTAGTAATG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 4444444445 ] 9999999990 ] 1234567890 ] ACACAATTTT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 5555555555 0000000001 1234567890 TATATTTCTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 1111111112 1234567890 ATTTTAATTG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 2222222223 1234567890 TTCCAGAGAT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 3333333334 1234567890 TGGCCATTAG .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 4444444445 1234567890 TTAGATGGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 5555555556 1234567890 GCATATATTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 6666666667 1234567890 ATAATCTGGA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 7777777778 1234567890 AATAGCCACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555555 8888888889 1234567890 ATAGACAATA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 5555555556 ] 9999999990 ] 1234567890 ] TAACTTTTTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 6666666666 0000000001 1234567890 TTTTAAATGC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 1111111112 1234567890 TGTAACTCCA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 2222222223 1234567890 AACTATAGTA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 3333333334 1234567890 CTTTCAGAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 4444444445 1234567890 CACTCACAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 5555555556 1234567890 TTCATGGTAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 6666666667 1234567890 AGAGGAAAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666666 7777777778 1234567890 ACCTCGTACT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... [ [ [ #C.hircus_EU581862 #Benggala1 #Benggala #Benggala3 #Jwrandu1 #Jwrandu2 #Jwrandu #Kacang1 #Kacang2 #Marica1 #Marica2 #Muara1 #Muara2 #Samosir #Shiba_goat_D82963 7777777777 0000000001 1234567890 TTCTAAAGCA C......... C......... C......... .......... .......... .......... .......... .......... C......... C......... C......... C......... C......... .......... 7777777777 1111111112 1234567890 CTTTCTGATA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......G.. .......G.. .......... .......... 7777777777 2222222223 1234567890 CAAGCTCTGT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 3333333334 1234567890 TCTTTGGTCT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 4444444445 1234567890 TTCTAGCTAC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 7777777777 5555555556 1234567890 TTTCCACCTC ...T...... ...T...... ...T...... .......... .......... .......... .......... .......... .......... .......... ...T...... ...T...... .......... .......... 7777777777 6666666667 1234567890 TTTGTAAATT .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 777] 777] 123] GCA ... ... ... ... ... ... ... ... ... ... ... ... ... ... 6666666666 8888888889 1234567890 TGGATGGAAA .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 6666666667 ] 9999999990 ] 1234567890 ] GCTCCCTACC .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... .......... 115 116 Lampiran 7 Pensejajaran berganda nukleotida gen GDF9 pada kambing lokal [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 1 1234567890 TCGGAGCCTC .......... .......... .......... .......... .......... .......... .......... 1111111112 1234567890 AGGAAGCGGC .......... .......... .......... .......... .......... .......... .......... 2222222223 1234567890 CGAAATCGGA .......... .......... .......... .......... .......... .......... .......... 3333333334 1234567890 CGGACTGAGT .......... .......... .......... .......... .......... .......... .......... 4444444445 1234567890 AGAAGAGCGA .......... .......... .......... .......... .......... .......... .......... 5555555556 1234567890 GGGGCCTGTC .......... .......... .......... .......... .......... .......... .......... 6666666667 1234567890 AACCTCCAGA .......... .......... .......... .......... .......... .......... .......... 7777777778 1234567890 GCTGCCCTAG .......... .......... .......... .......... .......... ..C....... ..C....... 8888888889 1234567890 GGTCGGGTGT .......... .......... .......... .......... .......... .......... .......... 1 ] 9999999990 ] 1234567890 ] CCTGGCGCGG .......... .......... .......... .......... .......... .......... .......... [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 1111111111 0000000001 1234567890 TGGAAAGCGT .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111112 1234567890 CTTTGAGGCC .......... .......... .......... .......... .......... .......... .......... 1111111111 2222222223 1234567890 GCCGCCTGGC .......... .......... .......... .......... .......... .......... .......... 1111111111 3333333334 1234567890 AGTCAACGCG .......... .......... .......... .......... .......... .......... .......... 1111111111 4444444445 1234567890 AGGTTTTGTA .......... .......... .......... .......... .......... .......... .......... 1111111111 5555555556 1234567890 CTGTCATGTC .......... .......... .......... .......... .......... .......... .......... 1111111111 6666666667 1234567890 CAGTAAGCTG .......... .......... .......... .......... .......... .......... .......... 1111111111 7777777778 1234567890 GCAAACCGGA .......... C......... C......... C......... C......... C......... C......... 1111111111 8888888889 1234567890 ATCGGCTATG .......... .......... .......... .......... .......... .......... .......... 1111111112 ] 9999999990 ] 1234567890 ] GTCAGGCCCT .......... .......... .......... .......... .......... .......... .......... [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 2222222222 0000000001 1234567890 CAGCGACCGC .......... .......... .......... .......... .......... .......... .......... 2222222222 1111111112 1234567890 AAGACCGGAA .......... .......... .......... .......... .......... .......... .......... 2222222222 2222222223 1234567890 TCAGGGACAC .......... .......... .......... .......... .......... .......... .......... 2222222222 3333333334 1234567890 GGATCTGAGA .......... .......... .......... .......... .......... .......... .......... 2222222222 4444444445 1234567890 CTTCTCTTGT .......... .......... .......... .......... .......... .......... .......... 2222222222 5555555556 1234567890 CAGCCACCAG .......... .......... .......... .......... .......... .......... .......... 2222222222 6666666667 1234567890 ATTAGAATCC .......... .......... .......... .......... .......... .......... .......... 2222222222 7777777778 1234567890 ACTGGCAGGG .......... .......... .......... .......... .......... .......... .......... 2222222222 8888888889 1234567890 TGTAGATTTT .......... .......... .......... .......... .......... .......... .......... 2222222223 ] 9999999990 ] 1234567890 ] CCCAAGAGAT .......... .......... .......... .......... .......... .......... .......... [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 3333333333 0000000001 1234567890 AATAAAAATG .......... .......... .......... .......... .......... .......... .......... 3333333333 1111111112 1234567890 TTTCCATTTA .......... .......... .......... .......... .......... .......... .......... 3333333333 2222222223 1234567890 GCCACAAGAG .......... .......... .......... .......... .......... .......... .......... 3333333333 3333333334 1234567890 TCCTTAGGTA .......... .......... .......... .......... .......... .......... .......... 3333333333 4444444445 1234567890 ATAGTACAGC .......... .......... .......... .......... .......... .......... .......... 3333333333 5555555556 1234567890 AAGTGATGAG .......... .......... .......... .......... .......... .......... .......... 3333333333 6666666667 1234567890 CAGCTGTTAG .......... .......... .......... .......... .......... .........T .........T 3333333333 7777777778 1234567890 TGATTGTAGG .......... .......... .......... .......... .......... CCT..T..-CCT..T..-- 3333333333 8888888889 1234567890 AGGGAAGAGG .......... .......... .......... .......... .......... .A.C.TATT. .A.C.TATT. 3333333334 ] 9999999990 ] 1234567890 ] CAACCTCCGT .......... .......... .......... .......... .......... ..GTA.T.CA ..GTA.T.CA [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 4444444444 0000000001 1234567890 CGCAGCCGGC .......... .......... .......... .......... .......... A.T.-----. A.T.-----. 4444444444 1111111112 1234567890 TTCTCCCTCA .......... .......... .......... .......... .......... A.A...T.A. A.A...T.A. 4444444444 2222222223 1234567890 CAGAGTTCT.............................................A..GC.GAGA A..GC.GAGA 4444444444 3333333334 1234567890 GCCTGGGGCC .......... .......... .......... .......... .......... .AACTA...A .AACTA...A 4444444444 4444444445 1234567890 GAATGTCATG .......... .......... .......... .......... .......... A...AG..-. A...AG..-. 4444444444 5555555556 1234567890 TTGCCCACTG .......... .......... .......... .......... .......... ..CT.TG..C ..CT.TG..C 4444444444 6666666667 1234567890 TTCACTGCCC .......... .......... .......... .......... .......... .CTGGAAT.T .CTGGAAT.T 4444444444 7777777778 1234567890 AGGAGGAAGG .......... .......... .......... .......... .......... CAATTTC..T CAATTTC..T 4444444444 8888888889 1234567890 CAGGAACGTC .......... .......... .......... .......... .......... TGT...TT.. TGT...TT.. 4444444445 ] 9999999990 ] 1234567890 ] GCGTGTGAGG .......... .......... .......... .......... .......... T.TCACCCTC T.TCACCCTC [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 5555555555 0000000001 1234567890 CGATTCCATT .......... .......... .......... .......... .......... .A.A..A.CA .A.A..A.CA 5555555555 1111111112 1234567890 TTCGGAAAGT .......... .......... .......... .......... .......... .G.A.T..A. .G.A.T..A. 5555555555 2222222223 1234567890 GCTTTCTACT .......... .......... .......... .......... .......... AAA....... AAA....... 5555555555 3333333334 1234567890 TAAGGATACT .......... .......... .......... .......... .......... .......... .......... 5555555555 4444444445 1234567890 GCTGTCCCGT .......... .......... .......... .......... .......... .......... .......... 5555555555 5555555556 1234567890 ACAGCTGACT .......... .......... .......... .......... .......... .......... .......... 5555555555 6666666667 1234567890 GACTTTTCCA .......... .......... .......... .......... .......... .......... .......... 5555555555 7777777778 1234567890 CTTGATGCAA .......... .......... .......... .......... .......... .......... .......... 5555555555 8888888889 1234567890 CGTGTCTGTA .......... .......... .......... .......... .......... .......... .......... 5555555556 ] 9999999990 ] 1234567890 ] AATGTTTTTT .......... .......... .......... .......... .......... .......... .......... 117 118 [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 6666666666 0000000001 1234567890 TCATGGAATT .......... .......... .......... .......... .......... .......... .......... 6666666666 1111111112 1234567890 GCTGGCCATG .......... .......... .......... .......... .......... .......... .......... 6666666666 2222222223 1234567890 TGTAATCTAG .......... .......... .......... .......... .......... .......... .......... 6666666666 3333333334 1234567890 GACTAAGTTC .......... .......... .......... .......... .......... .......... .......... 6666666666 4444444445 1234567890 TCATTCAGCA .......... .......... .......... .......... .......... .......... .......... 6666666666 5555555556 1234567890 GTTTGGGTGT .......... .......... .......... .......... .......... .......... .......... 6666666666 6666666667 1234567890 GTTTTATAAC .......... .......... .......... .......... .......... .......... .......... 6666666666 7777777778 1234567890 CCTCCTTGGC .......... .......... .......... .......... .......... .......... .......... 6666666666 8888888889 1234567890 AGATCCGAGC ........A. ........A. ........A. ........A. ........A. ........A. ........A. 6666666667 ] 9999999990 ] 1234567890 ] CAGGGAGGTT .......... .......... .......... .......... .......... .......... .......... [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 7777777777 0000000001 1234567890 TGGCAGACAA .......... .......... .......... .......... .......... .......... .......... 7777777777 1111111112 1234567890 TGAACTTAAA .......... .......... .......... .......... .......... .......... .......... 7777777777 2222222223 1234567890 ATTTACCAGT .......... .......... .......... .......... .......... .......... .......... 7777777777 3333333334 1234567890 ATACCTCGGT .......... .......... .......... .......... .......... .......... .......... 7777777777 4444444445 1234567890 TTTATCAGCC .......... .......... .......... .......... .......... .......... .......... 7777777777 5555555556 1234567890 TGGGTACTGC .......... .......... .......... .......... .......... .......... .......... 7777777777 6666666667 1234567890 ACAGACTGGT .......... .......... .......... .......... .......... .......... .......... 7777777777 7777777778 1234567890 TCTGTATCTT .......... .......... .......... .......... .......... .......... .......... 7777777777 8888888889 1234567890 ACACGGTAAG .......... .......... .......... .......... .......... .......... .......... 7777777778 ] 9999999990 ] 1234567890 ] TATATTCAGC .......... .......... .......... .......... .......... .......... .......... [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 8888888888 0000000001 1234567890 TCTTCTTAGT .......... .......... .......... .......... .......... .......... .......... 8888888888 1111111112 1234567890 GTACCCATGT .......... .......... .......... .......... .......... .......... .......... 8888888888 2222222223 1234567890 CTCATTTATC .......... .......... .......... .......... .......... .......... .......... 8888888888 3333333334 1234567890 TGAAACATAA .......... .......... .......... .......... .......... .......... .......... 8888888888 4444444445 1234567890 TCTTTAACCT .......... .......... .......... .......... .......... .......... .......... 8888888888 5555555556 1234567890 GATCTTAGCT .......... .......... .......... .......... .......... .......... .......... 8888888888 6666666667 1234567890 CAGAGGCAAG ..A....... .......... ..A....... .......... .......... .......... .......... 8888888888 7777777778 1234567890 AACATAAATA .......... .......... .......... .......... .......... .......... .......... 8888888888 8888888889 1234567890 CTTATACAGG .......... .......... .......... .......... .......... .......... .......... 8888888889 ] 9999999990 ] 1234567890 ] TAAGAACAAA .......... .......... .......... .......... .......... .......... .......... [ [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 9999999999 0000000001 1234567890 AATACCTAGC .......... .......... .......... .......... .......... .......... .......... 9999999999 1111111112 1234567890 AAATTCTCCC .......... .......... .......... .......... .......... .......... .......... 9999999999 2222222223 1234567890 ACCATTTGAG .......... .......... .......... .......... .......... .......... .......... 9999999999 3333333334 1234567890 TGATTAATAA .......... .......... .......... .......... .......... .......... .......... 9999999999 4444444445 1234567890 AAAGCAAACC .......... .......... .......... .......... .......... .......... .......... 9999999999 5555555556 1234567890 TTTCATTTAT .......... .......... .......... .......... .......... .......... .......... 9999999999 6666666667 1234567890 TTTCTAGCAC .......... .......... .......... .......... .......... .......... .......... 9999999999 7777777778 1234567890 CTGAAAACGT .......... .......... .......... .......... .......... .......... .......... 9999999999 8888888889 1234567890 CTTTAAACCT .......... .......... .......... .......... .......... .......... .......... 1 9999999990 9999999990 1234567890 TTCAATTAAA .......... .......... .......... .......... .......... .......... .......... ] ] ] ] [ [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 1111111111 0000000000 0000000001 1234567890 GTATGCATAG .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000000 1111111112 1234567890 ATGAATGAGC ........C. .......... .......... .......... .......... .......... .......... 1111111111 0000000000 2222222223 1234567890 AGACCCAAAT .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000000 3333333334 1234567890 AAAGGCCCCA .......... ......A... ......A... ......A... ......A... ......A... ......A... 1111111111 0000000000 4444444445 1234567890 AGAAATTATA .......... ...T...... ...T...... ...T...... ...T...... ...T...... ...T...... 1111111111 0000000000 5555555556 1234567890 CTTTAAAGTG .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000000 6666666667 1234567890 AGTCAAGATT .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000000 7777777778 1234567890 AAATGTGATG .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000000 8888888889 1234567890 TCCTCCTTTT .......... .......... .......... .......... .......... .......... .......... 1111111111 0000000001 9999999990 1234567890 TAAAGCATAT .......... .......... .......... .......... .......... .......... .......... ] ] ] ] [ [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 1111111111 1111111111 0000000001 1234567890 TGCAGTATTC .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 1111111112 1234567890 CAAGTACATA .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 2222222223 1234567890 TCCTTAAAAG .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 3333333334 1234567890 GCTGCGAGAA .......... ....A..... ....A..... ....A..... ....A..... ....A..... ....A..... 1111111111 1111111111 4444444445 1234567890 CTAGGCAAAA .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 5555555556 1234567890 TAGCAGTTCT .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 6666666667 1234567890 CTGCTCTCTG .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 7777777778 1234567890 GAATCTCAAT .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111111 8888888889 1234567890 TTCAGTTGTG .......... .......... .......... .......... .......... .......... .......... 1111111111 1111111112 9999999990 1234567890 AATTTCTCTC .......... .......... .......... .......... .......... .......... .......... ] ] ] ] 119 120 [ [ [ [ #KACANG-tunggal(08-K1) #KACANG-kembar(07-K2) #PE-tunggal(04-PE1) #PE-kembar(15-PE3) #MUARA-tunggal(06-R1) #MUARA-kembar(05-R2) #SAMOSIR-tunggal(14-S1) #SAMOSIR-kembar(12-S2) 1111111111 2222222222 0000000001 1234567890 ACCCTCCAAA .......... .......... .......... .......... .......... .......... .......... 1111111111 2222222222 1111111112 1234567890 TCAACATGCA .......... .......... .......... .......... .......... .......... .......... 111111] 222222] 222222] 123456] GTAAAT ...... ...... ...... ...... ...... ...... ......