hermafroditisme : suatu alternatif pola reproduksi
advertisement

sumber:www.oseanografi.lipi.go.id
ISSN 0216- 1877
Oseana, Volume XXIV, Nomor 3, 1999 : 27 - 33
HERMAFRODITISME : SUATU ALTERNATIF POLA
REPRODUKSI PADA IKAN
oleh
Sasanti R. Suharti 1)
A BS T R A CT
Hermaphoroditism ia a usual phenomenon model in many fishes. Sex reversal
is generally chosen if relationship between reproductive success an size or age are
different between sexes. Individuals should change sex when other sex has a higher
reproductive value. Sex change mechanism is determined by the existing of social
organization and mating system in reef fish. The process seems to be the factor that
regulating the ecosocial structure and position of each individual in the community.
In many cases, sex reversal needs a stimulus from conspecific fish with smaller size
or all fish have an ability to change sex depending on ecological change and changing in
mating system.
1975). Sejak itu hampir semua jenis ikan dalam
suku ini menunjukkan gejala adanya
hermafroditisme. Gejala demikian juga
ditemui di beberapa suku ikan karang lainnya.
Perubahan seks dari betina ke jantan
(protogini) ditemukan pada 14 famili ikan.
Sebelas diantaranya merupakan ikan penghuni
terumbu karang. Sebaliknya, perubahan dari
jantan ke betina (protandri) dijumpai di 8 suku,
3 diantaranya ditemukan hidup di terumbu
karang (WARNER 1984).
Perubahan seks pada ikan kemungkinan
disebabkan adanya perubahan fisiologi yang
bersifat endogenus yang berasosiasi dengan
ukuran tubuh dan umur (WARNER 1975).
Tetapi pengamatan secara langsung
menjelaskan bahwa perubahan yang terjadi di
PENDAHULUAN
Strategi reproduksi pada ikan-ikan Teleost dapat bervariasi. Hampir semua jenis ikan
mempunyai jenis kelamin (seks) yang terpisah
(diocous atau gonokoristik), tetapi banyak jenis
dimana individunya mempunyai kemampuan
untuk berubah jenis kelamin. Fenomena ini
dikenal sebagi hermafroditisme, yakni ikan
betina berubah kelamin menjadi ikan jantan
atau sebaliknya (HOAR 1969).
Perubahan jenis kelamin (sex change)
ditemukan secara luas pada famili ikan laut,
terutama yang mendiami terumbu karang
(WARNER 1984; ROSS 1990). Fenomena ini
pertama kali dijumpai pada beberapa anggota
dari suku Labridae (wrasse) (REINBOTH
1)
Balitbang Biologi Laut-LIPI, Jakarta
27
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
dalam kelompok sosial ikan yang terstruktur
dan mantap hampir selalu memicu terjadinya
perubahan seks melalui proses perilaku
(ROBERTSON 1972; FRICKE & FRICKE
1977; SHAPIRO 1979). Sebagai contoh
perubahan seks terjadi setelah adanya
pemindahan atau pengeluaran ikan jantan yang
dominan dari kelompok sosial yang sudah
tetap/mantap. Tulisan ini bertujuan untuk
mengulas perubahan kelamin pada ikan laut
yang bersifat protogini.
yang bersamaan dan akan melakukan
perilaku perkawinan (mating behaviour)
seperti pada ikan gonokrist, dimana
individu dapat berfungsi baik sebagai
betina ataupun jantan pada waktu
perkawinan.
2. Sekuensial (sequential hermaphroditism) yakni sebagian individu dalam daur
hidupnya berfungsi sebagai individu
dengan seks yang berbeda dan kemudian
berganti seks secara sukses dan berperan
sebagai seks berlawanan dalam sisa
hidupnya.
HERMAFRODITISME
Perubahan seks adalah merupakan
alternatif dari pola reproduksi yang umum
terjadi pada ikan laut maupun ikan air tawar.
Hal
ini
memungkinkan
individu
memaksimalkan
masa
keberhasilan
reproduksinya dengan berfungsi sebagai jenis
kelamin tertentu ketika kecil dan menjadi jenis
kelamin yangberbeda waktu besar (dewasa)
(GHISELIN 1969). Menurut SHAPIRO
(1988) perubahan seks dapat terjadi kapan saja
pada individu yang sudah dewasa dengan
ukuran tertentu, jika ada kondisi rangsangan
yang sesuai. Lebih jauh ia menjelaskan
individu dapat merubah seksnya setiap saat
dalam kehidupan dewasanya sampai ratio seks
dalam populasi tercapai.
Menurut ROSS (1990), hermafroditisme pada ikan dapat digolongkan menjadi:
1. Sinkronous (synchronous hermaphroditism) dimana individu menghasilkan
baik telur maupun sperma pada waktu
bersamaan.
Hermafrodit sinkronus umumnya terjadi
pada situasi dimana secara ekstrim
kepadatan populasi ikan rendah dan sulit
mendapatkan pasangan (WARNER &
ROBERTSON 1978). Sebagai contoh
anggota dari marga serranus (Serranidae)
dimana individu mengandung baik telur
maupun sperma yang masak pada waktu
Hermafrodit yang sekuensial dapat dibedakan
dalam 2 macam :
1. Protoginus yaitu individu lahir sebagai
betina dan berubah kelamin menjadi jantan
2. Protandrus yaitu individu dilahirkan
sebagai jantan dan berubah menjadi betina
sampai akhir hidupnya.
Dalam ikan-ikan Teleost, hermafrodit
yang protoginus merupakan pola yang paling
umum dan banyak dijumpai (CHOAT &
ROBERTSON 1975; WARNER & ROBERTSON 1978) Ini dapat dijumpai pada
ikan-ikan Labridae, Scaridae, Pomacentridae,
beberapa Serranidae, Sparidae dan sejumlah
ikan-ikan tropis lainnya. Pada jenis yang
protoginus, secara umum betina akan berubah
sel kelaminnya menjadi jantan yang sekunder
setelah masa reproduksi berakhir (KOBAYASHI & SUZUKI 1990).
Perubahan seks pada ikan yang
hermafrodit protoginus terjadi karena beberapa
faktor, antara lain berhubungan dengan sistem
sosial dimana individu yang besar, umumnya
jantan yang dominan, mempunyai derajat
reproduksi yang tinggi (REINBOTH 1973;
ROBERTSON & CHOAT 1974; WARNER et
al. 1975) atau karena faktor sosial dimana
betina berubah seks karena jantan yang
dominan dipindahkan atau mati (ROBERTSON 1972; WARNER et al. 1975).
28
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
ROBERTSON 1972). Sebagai contoh
pengeluaran seekor jantan dominan dari suatu
grup hirarki yang kuat akan menyebabkan
seekor betina yang besar dan dominan untuk
berubah seks.
Dalam hermafrodit yang protoginus,
ikan jantan yang besar dan dominan akan
memonopoli sistem perkawinan dan ikan
jantan kecil lainnya akan menjadi tidak suskes
dalam reproduksinya. Dalam sistem
perkawinan yang demikian jantan yang besar
ukuran tubuhnya akan berhasil bersaing
dengan jantan yang berukuran lebih kecil dan
untuk itu mendapatkan keberhasilan
reproduksi yang lebih besar (ROBERTSON &
CHOAT 1974; ROBERTSON & WARNER
1978; WARNER & HOFFMAN 1980;
WARNER 1984).
Selain itu keberadaan dari sejumlah
ikan jantan yang berukuran kecil dan jantan
konspesifik lainnya yang berukuran relatif
besar dalam suatu daerah jelajah (home arange)
dalam suatu populasi, kemungkinan dapat
menyebabkan terjadinya perubahan seks.
Beberapa nilai ambang dari proporsi ikan ikan
yang berukuran kecil dalam suatu 'home range'
akan merangsang/memicu terjadinya
perubahan seks (ROSS 1990).
IDENTITAS SEKSUAL
Pada ikan yang protoginus, terutama
suku Scaridae dan Labridae dikenal 4 macam
jenis kelamin yaitu betina, jantan tingkat
pertama (primary male), jantan tingkat kedua
(secondary male) dan individu yang sedang
dalam masa transisi seksual.
Jenis kelamin pada hermafrodit protoginus
dibedakan atas dua macam, yaitu :
1. Jantan tingkat pertama (primary male)
yaitu lahir sebagai jantan dan merupakan
jantan sejati (gonokrist). Tipe ini tidak
mengalami perubahan seks dalam hipnya
(REINBOTH 1977).
2. Jantan tingkat kedua (secondary male)
yaitu berasal dari betina yang berubah seks
(REINBOTH 1977).
Morfologi sel kelamin dari masingmasing tipe jantan ini bebeda secara
mikroskopis
(REINBOTH
dalam
ROBERTSON & CHOAT 1974). Selanjutnya
ia membedakan kondisi antara jantan tingkat
pertama dan tingkat kedua yaitu dengan
memakai istilah diandri dan monandri.
Monandri adalah dalam satu individu jantan
terdapat hanya jantan tingkat kedua, sedangkan
diandri adalah dalam satu individu jantan
terdapat baik primary male maupun secondary male. Diandri ini umum dijumpai dalam
populasi sebagai dikromatisme yang seimbang
secara genetik terhadap hermafroditisme
(ROBERTSON & CHOAT 1974; WARNER
et al. 1975).
SISTEM PERKAWINAN (MATING
SYSTEM) DAN PEMELIHARAAN
OLEH INDUK (PARENTAL CARE)
ROBERTSON & CHOAT (1974);
WARNER & HOFFMAN (1980) WARNER
(1984) menjelaskan bahwa hermafrodit
protoginus berkaitan dengan sistem
perkawinan, dimana jantan dewasa yang
berukuran besar memonopoli reproduksi. Hal
ini sesuai dengan model dari GHISELIN
(1969) mengenai model keuntungan dari
ukuran tubuh (size advantage model). Warner
(1984) menerangkan bahwa ada suatu
keuntungan dengan memiliki ukuran tubuh
yang besar dalam 'mating system'. Dalam
KONTROL SOSIAL DAN
LINGKUNGAN
Kontrol sosial pada saat terjadinya
perubahan seks biasanya digambarkan pada
ikan-ikan yang hidupnya haremik
(ROBERTSON & HOFFMAN 1977) dan
beberapa spesies yang hidup dalam kelompok
yang kurang terorganisir (FISHELSON 1970;
29
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
'mating system' pada jenis yang protoginus,
ikan jantan yang besar bersifat 'promiscuous'
(tidak memilih-milih/ dengan siapa saja) dan
dapat membuahi sejumlah ikan betina. Hal
demikian akan memberikan keuntungan yang
selektif dari jantan yang berukuran besar dan
mendapatkan kesuksesan yang tinggi dalam
perkawinan (WARNER 1984).
Dengan adanya 2 tipe jantan ('primary
dan secondary male') dalam spesies yang
protoginus, erat kaitannya dengan 2 strategi
perkawinan yang berbeda yakni pertama,
perkawinan antar sepasang jantan dengan
warna yang cerah (terminal phase) dengan
seekor betina (pair spawning) dan kedua,
perkawinan grup (group/multiple spawning)
dimana banyak jantan dengan warna yang
kurang cerah mengawini satu ekor beina.
Bagi ikan yang mempunyai sistem
'parental care', hal ini dapat membatasi
kesuksesan reproduksi dari ikan jantan yang
bersifat teritorial, karena hal ini akan
mengurangi kompetisi antara ikan yang
berukuran kecil dan besar sehingga tidak ada
seleksi seksual yang kuat bagi ikan yang
berukuran besar seperti terjadi pada
hermafrodit protoginus (WARNER &
LEJEUNE) 1985. Menurut SCHMALE (1981)
untuk terjadinya perubahan seks paling tidak
pasangan dalam perkawinan harus bebas dari
'parental care'
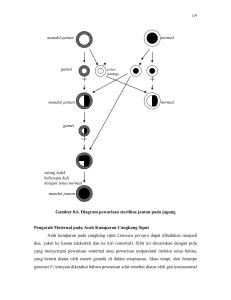
Gambar 1.
Keterangan:
Ketika jantan yang besar memonopoli perkawinan atas jantan yang berukuran lebih kecil, maka
fertilitas jantan yang berukuran besar meningkat secara dramatis pada satu titik pertumbuhan
tertentu. Untuk itu menjadi betina sewaktu kecil dan berubah menjadi jantan setelah besar, dalam
hal ini lebih disukai.
30
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
jantan tertentu ditentukan oleh ukuran dari
jantan lain dalam populasi setempat
(WARNER 1984)
WARNER (1988) menjelaskan, pada
jenis yang protoginus dimana jantan dewasa
yang dominan memonopoli perkawinan dan
terlihat keuntungan yang nyata untuk menjadi
betina diwaktu kecil dan hanya berubah
menjadi jantan ketika ukuran yang cukup
secara kompetitif telah dicapai untuk kawin.
Secara umum model/hipotesa ini menerangkan
bahwa, individu sebaiknya merubah seksnya
ketika jenis kelamin yang lain mempunyai nilai
reproduksi yang lebih besar. Keterangan ini
dapat dilihat pada gambar di bawah ini.
KEUNTUNGAN DARI UKURAN
TUBUH (SIZE ADVANTAGE)
Ukuran tubuh secara nyata dapat
menyebabkan kesuksesan dalam reproduksi,
sehingga 'mating system' memegang peranan
penting (WARNER 1988). Hal ini sesuai
dengan penjelasan SHAPIRO (1988) bahwa
kesuksesan reproduksi meningkat sejalan
dengan bertambah besarnya ukuran ikan.
Hipotesa/model mengenai keuntungan dari
ukuran tubuh menjelaskan bahwa ketika
dominasi tergantung dari ukuran tubuh,
kemungkinan keberhasilan perkawinan dari
Gambar 2.
Keterangan:
Jika kompetisi antar jantan sangat kuat, hanya individu yang terbesar ukuran tubuhnya akan
sukses dalam perkawinan (mating). Walaupun fekunditas betina meningkat dengan bertambahnya
ukuran tubuh, tapi dalam hal mating system pengaruh ukuran tubuh jantan lebih dominan.
31
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
DAFTAR PUSTAKA
(Labridae) Pubbl. Staz. Zool. Napoli 39
supll. : 550-573.
CHOAT, J.H. & D.R. ROBERTSON, 1975.
Protogynus hermaphroditism in fishes
of the family Scaridae. In: Intersexually in the animal kingdom.
(REINBOTH, R. ed). Springer-Verlag,
Berlin, 263-283.
REINBOTH, R. 1977. Thalassoma
bifasciatum (Labridae)-paar-und
gruppenlaichen im korallenriff. Summary. Publ. zu Wissen. Filmen 10(4):
3.
ROBERTSON, D.R. 1972. Social control of
sex reversal in a coral reef fish. Science N.Y., 177 : 1007-1009.
FISHELSON, L. 1970. Protogynus sex-reversal in the fish Anthias squamipinnis
(Teleostei-Anthiidae) regulated by
presence or absence of male fish. Nature 227 : 90.
ROBERTSON, D.R. & H. CHOAT. 1974.
Protogynus hermaphroditism and social
system in labrid fishes. Proc. 2 nd Int.
Symp. Coral Reefs G.B.R. Committee,
Brisbane, 1 : 217-225.
FRICKE, H.W. & S. FRICKE. 1977. Monogamy and sex change by aggressive
dominance in coral reef fish. Nature
266 : 830-832.
ROBERTSON, D.R. & S.G. HOFFMAN.
1977. The role of female mate choice
and predation in the mating system of
some tropical labroid fishes. Z.
Tierpsychol. 45 : 298-320.
GHISELIN, M.T. 1969. The evolution of hermaphroditism among animals. Quart.
Rev. Biol. 44 : 189-454.
HOAR, W.W. 1969. Reproduction. In: Fish
Physiology. (W.S. HOAR and DJ.
RANDALL, eds.), vol. 3. Acad. Press,
New York.
ROBERTSON, D.R. & R.R. WARNER. 1978.
Sexual patterns in labroid fishes of the
western Carribean, II: The parrotfishes
(Scaridae): Smithsonian Contrib Zool.
225: 1-26.
HOFFMAN, S.G. 1983. Sex-related foraging
behavior in sequentially hermaphroditic hogfish (Bodianus spp.). Ecology
64 : 798-808
ROOS, R.M. 1990. The evolution of sex
change mechanism in fishes. Env. Biol
Fish. 29: 81-93.
KOBAYASHI, K. and K. SUZUKI. 1990.
Gonadogenesis and sex succession in
protogynus wrasse, Cirrhilabrus
temmincki, in Suruga Bay, Central Japan. Japanese J. Ichthyology 37 (3):
256-264.
SCHMALE. M.C. 1981. sexual selection and
reproductive success in males of bicolor
damselfish, Eupomacentrus partitus
(Pisces: Pomacentridae). Anim Behav.,
29: 1172-1184.
SHAPIRO, D.Y 1979. Social behavior, group
structure, and the control of sex reversal in hermaphroditic fish. In: Advances
in the study of behavior. J.S.
RASENBLATT, R.A. HINDE, C.
BEER and M.C. BUSNEL (eds), vol.
10: 43-102. Acad. Press., N. York.
REINBOTH, R. 1973. Dualistic reproductive
behavior in the protogynus wrasse
Thallasoma bifasciatum and some observations on its day-night change over.
Helgol. Wiss. Meers. 24 : 174-191.
REINBOTH, R. 1975. Spontaneous and hormone induce sex-inversion in wrasse
32
Oseana, Volume XXIV no. 3, 1999
sumber:www.oseanografi.lipi.go.id
WARNER, R.R. 1988. Sex change in fishes:
hypotheses, evidence, and objections.
Env. Biol. Fish. 22(2): 81-90.
SHAPIRO, D.Y. 1981. Behavioral charges in
protogynus sex reversal in coral reef
fish in the laboratory. Anim. Behav.
29: 1185-1198.
WARNER, R.R. & D.R. ROBERTSON. 1978.
Sexual pattern in the labroid fishes of
the western Carribean, I: The wrasse
(Labridae). Smitsonian Contrib. Zool.
254: 1-27.
SHAPIRO, D.Y. 1984. Sex reversal and
sociodemographic processes in coral
reef fishes. In: Reproduction: Strategies
and Tactics. G.W. POTTS & R.J.
WOOTON (Eds.) Acad. Press., London.
WARNER, R.R. & S.G. HOFFMAN. 1980.
Local population size as a determinant
of mating system and sexual composition in the two tropical marine fishes
(Thalassoma spp). Evolution 34:
SOS-SIS.
WARNER, R.R. & P. LEJEUNE. 1985. Sex
change limited by paternal care: a test
using four Mediterranean labrid fishes,
genus Symphodus. Mar. Biol 87:
89-99.
WARNER, R.R., D.R. ROBERTSON & E.G.
LEIGH. 1975. Sex change and sexual
selection Science 190: 633-638.
SHAPIRO, D.Y. 1988. Behavioral influences
on gene structure and other new ideas
concerning sex change in fishes. Env.
Biol. Fish. 23 (4): 283-297.
WARNER, R.R. 1975. The adaptive significance of sequential hermaphroditsm in
animals. Am. Nat. 109: 61-82.
WARNER, R.R. 1982. Mating system, sex
change, and sexual demography in the
rainbow wrasse, Thalassoma
lucasanum. Copeis 3: 635-661.
WARNER, R.R. Mating systems and hermaphroditism in coral reef fishes. Amer.
Scient. 72: 129-136.
33
Oseana, Volume XXIV no. 3, 1999