BAB I PENDAHULUAN Latar Belakang Pengubahan energi sinar
advertisement

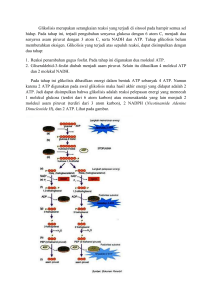
BAB I PENDAHULUAN 1.1 Latar Belakang Pengubahan energi sinar menjadi energi kimia (karbohidrat) atau yang disebut proses fotosintesis, yang mengubah energi kimia menjadi energi kerja pada peristiwa pernafasan dalam tubuh tumbuhan, dan manusia atau yang disebut dengan resprasi yang merupakan suatu rangkaian proses kehidupan di dunia ini. Respirasi atau pernafasan merupakan suatu proses pembongkaran (katabolisme atau desimilasi ) dimana energi yang tersimpan ditimbulkkan kembali untuk menyelenggarakan proses-proses kehidupan. Udara yang masuk dan keluar dari suatu tempat, maka udara yang keluar dari tempat tersebut mempunyai kadar oksigen ( O2 ) yang lebih rendah dan kadar gas CO2 yang tinggi dari pada udara yang masuk. Hal ini berarti bahwa kadar CO2 yang tinggi itu disebabkan oleh terurainya suatu zat organik. Menyusutnya zat organik tersebut diakhibatkan karena penggunaan O2 dan pengeluaran CO2 dan timbulnya panas (energi) itu semua manifesti dari proses respirasi yang terjadi di dalam tiap sel yang hidup. Tidak semua respirasi mengeluarkan CO2 karena respirasi ada 2 macam yaitu respirasi aerob dan anaerob yang terjadi dalam sel-sel tumbuhan. 1.2 Tujuan Untuk mengetahui arti dan peranan respirasi dan fotorespirasi bagi pertumbuhan tanaman. Untuk mengetahui proses atau mekanisme respirasi. Untuk mengetahui proses tau mekanisme respirasi. Untuk mengetahui faktor-faktor yang mempengaruhi proses respirasi. 1 2 BAB II PEMBAHASAN 2.1 Arti dan Peranan Respirasi dan fotorespirasi bagi Pertumbuhan Tanaman 2.1.1 Arti Respirasi Respirasi dalam biologi adalah proses mobilisasi energi yang dilakukan jasad hidup melalui pemecahan senyawa energi bernergi tinggi (SET) untuk digunakan dalam menjalankan fungsi hidup. Dalam pengertian kegiatan sehari-hari, respirasi dapat disamakan dengan pernapasan. Namun demikian, istilah respirasi mencakup proses-proses yang juga tidak tercakup pada istilah pernapasan.Pada dasarnya respirasi adalah proses oksidasi yang dialami SET sebagai unit penyimpanan energi kimia pada organisme hidup. 2.1.2 Manfaat Respirasi Bagi Tumbuhan Respirasi banyak memberikan manfaat bagi tumbuhan. Manfaat tersebut terlihat dalam proses respirasi dimana terjadi proses pemecahan senyawa organik, dari proses pemecahan tersebut maka dihasilkanlah senyawa-senyawa antara yang penting sebagai ”Building Block”. Building Block merupakan senyawa-senyawa yang penting sebagai pembentuk tubuh. Senyawa-senyawa tersebut meliputi asam amino untuk protein; nukleotida untuk asam nukleat; dan prazat karbon untuk pigmen profirin (seperti klorofil dan sitokrom), lemak, sterol, karotenoid, pigmen flavonoid seperti antosianin, dan senyawa aromatik tertentu lainnya, seperti lignin. Telah diketahui bahwa hasil akhir dari respirasi adalah CO2 dan H2O, hal ini terjadi bila substrat secara sempurna dioksidasi, namun bila berbagai senyawa di atas terbentuk, substrat awal respirasi tidak keseluruhannya diubah menjadi CO2 dan H2O. Hanya beberapa substrat respirasi yang dioksidasi seluruhnya menjadi CO2 dan H2O, sedangkan sisanya digunakan dalam proses anabolik, terutama di dalam sel yang sedang tumbuh. Sedangkan energi yang ditangkap dari proses oksidasi sempurna beberapa senyawa dalam proses 3 respirasi dapat digunakan untuk mensintesis molekul lain yang dibutuhkan untuk pertumbuhan. 2.1.3 Arti Fotorespirasi Fotorespirasi adalah sejenis respirasi pada tumbuhan yang dibangkitkan oleh penerimaan cahaya yang diterima oleh daun. Diketahui pula bahwa kebutuhan energi dan ketersediaan oksigen dalam sel juga memengaruhi fotorespirasi. Walaupun menyerupai respirasi (pernafasan) biasa, yaitu proses oksidasi yang melibatkan oksigen, mekanisme respirasi karena rangsangan cahaya ini agak berbeda dan dianggap sebagai proses fisiologi tersendiri. 2.1.4 Manfaat Fotorespirasi Peran fotorespirasi diperdebatkan namun semua kalangan sepakat bahwa fotorespirasi merupakan penyia-nyiaan energi. Dari sisi evolusi, proses ini dianggap sebagai sisa-sisa ciri masa lampau (relik). Atmosfer pada masa lampau mengandung oksigen pada kadar yang rendah, sehingga fotorespirasi tidak terjadi seintensif seperti masa kini. Fotorespirasi dianggap bermanfaat karena menyediakan CO2 dan NH3 bebas untuk diasimilasi ulang, sehingga dianggap sebagai mekanisme daur ulang (efisiensi). Pendapat lain menyatakan bahwa fotorespirasi tidak memiliki fungsi fisiologis apa pun, baik sebagai penyedia asam amino tertentu (serin dan glisin) maupun sebagai pelindung klorofil dari perombakan karena fotooksidasi. Karena tidak efisien, sejumlah tumbuhan mengembangkan mekanisme untuk mencegah fotorespirasi. Untuk menekan fotorespirasi, tumbuhan C4 mengembangkan strategi ruang dengan memisahkan jaringan yang melakukan reaksi terang (sel mesofil) dan reaksi gelap (sel selubung pembuluh, atau bundle sheath). Sel - sel mesofil tumbuhan C4 tidak memiliki Rubisco. Strategi yang diambil tumbuhan CAM bersifat waktu (temporal), yaitu memisahkan waktu untuk 4 reaksi terang (pada saat penyinaran penuh) dan reaksi gelap (di malam hari). 2.3 Tahapan Respirasi 2.3.1 Respirasi Aerob Respirasi aerob yaitu respirasi yang menggunakan oksigen oksigen bebas untuk mendapatkan energi. Ada beberapa tumbuhan yang kegiatan respirasinya menurun bila konsentrasi oksigen di udara dibawah normal, misalnya bayam, wortel dan beberapa tumbuhan lainnya. Persamaan reaksi proses respirasi aerob secara sederhana dapat dituliskan: C6H12O6 + 6H2O >> 6H2O + 6CO2 + 675 kal Dalam kenyataan reaksi yang terjadi tidak sesederhana itu. Banyak tahapan yang terjadi dari awal hingga terbentuknya energi. Reaksi-reaksi itu dapat dibedakan menjadi 3 tahapan yaitu glikolosis, siklus krebs dan transport elektron (Syamsuri, 1980). Pembongkaran sempurna terjadi pada oksidasi asam piruvat dalam respirasi aerob. Dari proses ini dihasilkan CO2 dan H2O serta energy yang lebih banyak , yaitu 38 ATP (Loveless, A.R, 1991). 5 Proses aerob dibagi menjadi sebagai berikut : a. Glikolisis Pada tahap awal dari glikolisis (dari kata latin, glykos = gula; lysis = pemecahan), karbohidrat diubah menjadi heksosa fosfat, yang kemudian dipecah menjadi dua molekul triosa fosfat. Selanjutnya kedua molekul triosa fosfat tersebut dioksidasi menjadi dua molekul piruvat. Disamping menyiapkan substrat untuk oksidasi dalam siklus asam sitrat, glikolisis juga menghasilkan sejumlah kecil ATP dan NADH. Glikolisis pada tumbuhan memiliki mekanisme pengaturan yang khas, berlangsung secara paralel antara glikolisis sitotol dan plastida, serta jalur-jalur alternatif dari reaksi-reaksi glikolisis di dalam sitosol. Pada hewan, substrat utama respirasi adalah glukosa dan produk akhirnya adalah piruvat. Pada tumbuhan, sukrosa merupakan bentuk gula utama yang ditranslolasikan dan oleh karenanya merupakan jenis gula utama yang diimpor oleh jaringan/organ non fotositesis, maka sukrosa diyakini merupakan substrat utama respirasi tumbuhan dan 6 hasil akhirnya tidak hanya piruvat, tetapi juga dapat berupa asam organik lainnya yaitu malat. Pada tahap awal glikolisis, sukrosa dipecah menjadi dua monosakarida, glukosa dan fruktosa, yang dapat segera masuk ke lintasan glikolisis. Terdapat dua lintasan yang memecah sukrosa di dalam tumbuhan. Pertama, pada sebagian besar jaringan tumbuhan, enzim sukrosa sintase yang ada di dalam sitosol, digunakan untuk memecah sukrosa dengan cara menggabungkan sukrosa dengan UDP menjadi fruktosa dan UDP-glukosa. Kemudian enzim UDP-glukosapirofosforilase mengubah UDP-glukosa dan pirofosfat (PPi) menjadi UTP dan glukosa-6-fosfat. Kedua, pada beberapa jaringan tumbuhan, enzim invertase yang terdapat pada dinding sel, vakuola atau sitosol menghidrolisis sukrosa menjadi dua heksosa, yaitu fruktosa dan glukosa, yang kemudian difosforilasi dalam suatu reaksi yang menggunakan ATP. Plastida, seperti kloroplas dan amiloplas, dapat juga menyediakan substrat untuk glikolisis. Pati disintesis dan dipecah hanya di dalam plastid. Plastid mengubah pati menjadi triosa fosfat menggunakan enzim-enzim glikolisis yang mengubah heksosa fosfat menjadi triosa fosfat. Senyawa karbon yang diperoleh dari pemecahan pati masuk ke dalam lintasan glikolisis di dalam sitosol. Senyawa karbon tersebut terutama dalam bentuk heksosa fosfat yang ditranslokasikan dari amiloplas atau triosa fosfat yang ditranslokasikan dari kloroplas. Hasil fotosintesis dapat juga langsung masuk lintasan glikolisis dalam bentuk triosa fosfat. Pada tahap awal glikolisis, tiap unit heksosa difosforilasi dua kali dan dipecah menjadi dua molekul triosa fosfat. Bergantung apakah pemecahan tersebut menggunakan enzim sukrosa sintase atau invertas, serangkaian reaksi pemecahan tersebut menggunakan dua sampai empat molekul ATP untuk tiap unit sukrosa. Reaksi-reaksi tersebut juga melibatkan dua dari tiga reaksi tidak dapat balik penting dari lintasan glikolisis yang dikatalisis oleh enzim heksokinase dan 7 fosfofruktokinase . Reaksi fosfofruktokinase adalah satu dari titik kendali glikolisis baik pada tumbuhan maupun hewan. Ketika molekul gliseraldehid-3-fosfat terbentuk, lintasan glikolisis memulai memanen energi pada tahap pemanenan energi dari glikolisis. Enzim gliseraldehid-3-fosfat dehidrogenase mengkatalisis oksidasi aldehid untuk membentuk asam karboksilat dan mereduksi NAD+ menjadi NADH. Reaksi tersebut melepas energi bebas dalam jumlah cukup untuk melakukan fosforilasi gliseraldehid-3-fosfat menggunakan fosfat inorganik dan membentuk 1,3-bisfosfogliserat. Molekul ini merupakan suatu pemberi gugus fosfat yang cukup kuat karena memiliki standar energi bebas untuk hidrolisis yan gcukup tinggi (-49.3 kJ per mol atau –11.8 kkal per mol). Pada tahap berikutnya gugus fosfat dari ato carbon ke 1 dari 1,3bisfosfogliserat dipindhakan ke molekul ADP untuk membentuk ATP dan molekul 3-fosfogliserat. Untuk tiap sukrosa yang masuk glikolisis akan dihasilkan empat ATP dari reaksi ini. Sintesis ATP dengan cara tersebut disebut fosforilasi tingkat substrat, yang melibatkan pemindahan langsung sebuah gugus fosfat dari molekul substrat ke ADP untuk membentuk ATP. Sintesis ATP melalui proses ini berbeda mekanismenya dengan sintesis ATP melalui fosforilasi oksidatif di mitokondria atau sintesis ATP pada proses fotosintesis. Pada dua reaksi berikutnya, fosfat dari molekul 3-fosfogliserat dipindahkan ke ataom karbon ke dua dan kemudian sebuah molekul air dilepas menghasilkan senyawa yang disebut fosfoenol piruvat (PEP). Gugus fosfat dari PEP memiliki standar energi bebas yang tinggi (61.9 kJ per mol atau –14.8 kkal per mol), sehingga PEP merupakan suatu donor fosfat yang baik untuk membentuk ATP. Dengan menggunakan PEP sebagai substrat, enzim piruvat kinase memindahkan gugus fosfat dari PEP ke ADP untuk membentuk ATP dan piruvat. Tahap terahir ini, yang merupakan tahap tidak dapat balik ketiga yang penting dalam glikolisis, menghasilkan empat molekul 8 ATP untuk tiap molekul sukrosa yang masuk ke dalam lintasan glikolisis. Di dalam tumbuhan, selain PEP diubah ke bentuk piruvat, PEP juga dapat mengalami karboksilasi untuk membentuk asam organik oksaloasetat (OAA) dengan bantuan enzim PEP karboksilase. OAA kemudian direduksi menjadi malat dengan bantuan enzim malat dehidrogenase dan menggunakan NADH. Malat yang dihasilkan dapat disimpan di dalam vakuola atau ditransport ke mitokondria dan masuk ke siklus asam sitrat. Berikut prosesnya: 1. Fosforilasi glukosa oleh ATP. Penambahan satu fosfat oleh ATP terhadap glukosa menghasilkan glukosa 6-fosfat dan ATP berubah menjadi ADP. 2. Penyusunan kembali diikuti dengan fosforilasi kedua. Hasil akhir berupa fruktosa 1,6- bifosfat. 3. Perubahan 1,6 fruktosa-bifosfat yang memiliki 6 atom C menjadi gliseraldehida 3-fosfat (3 atom C). 4. Selanjutnya fosfogliseraldehida bersenyawa dengan suatu asam fosfat (H3PO4) dan berubah menjadi 1,3 –disfosfogliseraldehid kemudian menjadi asam 1,3 –difosfogliserat dengan bantuan enzimdehidrogenase. Peristiwa ini terjadi karena adanya penambahan H2. 5. Dengan bantuan enzim transfosforilase fosfogliserat serta ion – ion Mg++, asam 1,3- difosfogliserat kehilangan satu fosfat sehingga berubah menjadi asam – 3 – fosfogliserat. 6. Selanjutnya asam – 3 – fosfogliserat menjadi asam – 2 – fosfogliserat karena pengaruh enzim fosfogliseromutase. 7. Dengan pertolongan enzim enolase dan ion – ion Mg++, maka asam- 2-fosfofogliserat melepaskan H2O dan menjadi asam -2fosfoenolpiruvat. 9 8. Perubahan terakhir dalam glikolisisadalah pelepasan satu fosfat dari asam-2-fosfoenolpiruvat menjadi asam piruvat. Enzim transfosforilase fosfopiruvat dan ion – ion Mg++ membantu proses ini sedang ADP meningkat menjadi ATP. (Pratiwi dkk, 2003) b. Dekarboksilasi Oksidatif Sebelum masuk ke tahap selanjutnya dalam mitokondria, asam piruvat terlebih dahulu akan diubah menjadi senyawa Asetil Co-A dan berlangsung dalam membrane mitokondria. Tahapannya sebagai berikut: i) Senyawa asam piruvat yang mengandung 3 atom karbon, dioksidasi dengan bantuan enzim piruvat dehidrogenase untuk melepas 1 atom karbonnya dan mengubahnya menjadi CO2. Bersamaan dengan terbentuknya CO2, NAD+ akan direduksi dan membentuk NADH. ii) Selanjutnya, terbentuklah senyawa dengan 2 atom karbon yang disebut acetyl group, yang kemudian akan ditambahkan dengan koenzim A membentuk Acetyl Koenzim-A 10 c. Siklus Krebs Siklus kreb berlangsung di matriks mitokondria. Asetil KoA bergabung dengan asam oksaloasetat membentuk asam sitrat. KoA dilepaskan sehingga memungkinkan untuk mengambil fragmen 2C lain dari asam piruvat. Pembentukan asam sitrat terjadi diawal siklus krebs , sementara itu sisa dua karbon dari glukosa dilepaskan sebagai CO2. Selama terjadi pembentukan – pembentukan , energy yang dibutuhkan dilepaskan untuk menggabungkan fosfat denga ADP membentuk molekul ATP. Pada siklus krebs , pemecahan rantai karbon pada glukosa selesai, Jadi, sebagai hasil dari glikoslisis , reaksi antara dan siklus krebs adalah pemecahan satu molekul glukosa 6 karbon menjadi 6 molekul 1 karbon, selain itu juga dihasilkan 2 molekul ATP dari glikolisis dan 2 ATP lagi dari siklus krebs. Perlu diingat bahwa tiap – tiap proses melepaskan atom hydrogen yang ditranspor ke sistem transport electron oleh molekul pembawa (Pratiwi, dkk. 2003). Siklus asam sitrat disebut juga siklus asam trikarboksilat untuk menunjukkan pentingnya dua molekul trikarboksilas, citrat dan isositrat, sebagai dua intermediet pertama dari siklus ini. Untuk berlangsungnya siklus asam sitrat, piruvat yang dihasilkan dari glikolisis harus ditrasport ke dalam matriks mitrokondria melalui protein transport khusus yang terdapat pada membran dalam mitrokondria. Setelah piruvat ada di dalam matriks, piruvat kemudian didekarboksilasi dalam suatu reaksi opksidasi oleh ensim piruvat dehidrogenase. Hasil dari reaksi ini adalah NADH, CO2 dan asam asetat dalam bentuk asetil koA. Piruvat dehidrogenase merupakan kompleks enzim yang mengkatalisis keseluruhan tiga proses ini, yaitu dekarboksilasi, oksidasi dan konjugasi asam asetat dengan KoA. Pada reaksi berikutnya enzim sitrat sintase menggabungkan grup asetil dari asetil koA dengan OAA, suatu asam dikarboksilat beratom karbon empat, untuk menghasilkan sitrat, yang kemudian diisomerasi oleh enzim aconitase untuk menghasilkan isositrat. Dua tahap reaksi berikutnya 11 adalah reaksi dekarbokislasi oksidatif berurutan yang masing-masing menghasilkan satu meolekul NADH, melepas satu moelkul CO2 dan membentuk suksinil koA. Sampai tahap ini sudah tiga molekul CO2 dilepas untuk tiap piruvat yang masuk ke mitokondria, atau 12 molekul CO2 dari tiap molekul sukrosa yang dioksidasi. Tahap berikutnya adalah oksidasi suksinil koA menjadi OAA, sehingga memungkinkan siklus asam sitrat terus berputar. Mula-mula energi bebas yang tersedia dari ikatan tioester pada seuksinil koA disimpan di dalam ATP dari ADP dan Pi melalui proses fosforilasi tingkat substrat yang dikatalisis oleh enzim suksinil koA sitetase. Suksinat yang dihasilkan dioksidasi menjadi fumarat oleh enzim suksinat edhidrogenase, yang berada pada membran dalam mitokondria dan merupakan enzim yang juga berperan di dalam rantai transport elektron. Elektron dan proton dilepas dari suksinat dari suatu reaksi redoks yang melibatkan FAD (flavin adenin dinukleotida). FAD secara kovalen terikat pada sisi aktif dari suksinat ehidrogenase dan melangsungkan reduksi FAD menjadi FADH2. Dua tahap terakhir dari siklus asam sitrat adalah hidrasi fumarat menjadi malat, yang selanjutnya dioksidasi oleh malat dehidrogenase menjadi OAA dan menghasilkan satu molekul NADH. Sebagai kesimpulan, setiap piruvat yang masuk mitokondria akan menghasilkan tiga molekul CO2, empat molekul NADH, satu molekul FADH2 dan satu molekul ATP. Siklus Asam Sitrat Tumbuhan Memiliki Keunikan Siklus asam sitrat tumbuhan tidak identik dengan siklus asam sitrat yang terjadi pada mitokondria hewan. Sebagai contoh apda tahapan yang dikatalisis oleh enzim suksinil koA sintetase, di tumbuhan menghasilkan ATP, sedangkan pada hewan menghasilkan GTP. Perbedaan yang kedua adalah adanya aktifitas enzim NAD+-malat yang tidak dijumpai pada organisme lain. Enzim ini mengkatalisis reaksi dekarboksilasi oksidatif dari malat menjadi piruvat: 12 Malat + NAD+ → piruvat + CO2 + NADH Keberadaan enzim NAD+-malat memungkinkan mitokondria tumbuhan mampu melakukan lintasan alternatif dari metabolisme PEP yang dihasilkan melalui glikolisis. Seperti diketahui, malat dapat disintesis dari PEP di sitosol dengan bantuan enzim PEP karboksilase dan malat dehidrogenase. Malat kemudian ditransport ke dalam matriks mitokondria, dimana enzim NAD+-malat dapat mengoksidasinya menjadi piruvat. Reaksi ini memungkinkan oksidasi sempurna dari itermediat-intermediat siklus asam sitrat, seperti malat atau sitrat. Alternatif lain dari jalur metabolisme PEP adalah digunakannya malat sebagai pengganti intermediat siklus asam sitrat. Malat yang masuk ke mitokondria hasil PEP karboksilase dapat mengisi kekurnag malat akibat keluarnya intermediat respirasi dari siklus. Misal 2 oksoglutarat dapat keluar siklus dan digunakan dalam asimilasi nitrogen. Hal ini dapat menyebabkan kekurang malat untuk kelangsung siklus. Adanya malat yang masuk ke matriks dari sitosol hasil metabolisme PEP dapat memenuhi kekurangan malat tersebut, sehingga siklus tetap berjalan. Memasuki siklus krebs, asetil KoA direaksikan dengan asam oksaloasetat (4C) menjadi asam piruvat (6C). selanjutnya asam oksaloasetat memasuki daur menjadi berbagai macam zat yang akhirnya menjadi asam oksalosuksinat. Dalam perjalanannya, 1C (CO2) dilepaskan. Pada tiap tahapan, dilepaskan energi dalam bentuk ATP dan hidrogen. ATP yang dihasilkan langsung dapat digunakan. Sebaliknya, hidrogen berenergi digabungkan dengan penerima hidrogen yaitu NAD dan FAD, untuk dibawa ke sistem transport elektron. Dalam tahap ini dilepaskan energi, dan hidrogen direasikan dengan oksigen membentuk air. Seluruh reaksi siklus krebs berlangsung dengan memerlukan oksigen bebas (aerob). Siklus krebs berlangsung didalam mitokondria (Syamsuri, 1980). 13 d. Transpor Elektron Energi yang terbentuk dari peristiwa glikolisis dan siklus krebs ada dua macam. Pertama dalam bentuk ikatan fosfat berenergi tinggi, yaitu ATP atau GTP (Guanin Tripospat). Energi ini merupakan energi siap pakai yang langsung dapat digunakan. Kedua dalam bentuk transport elektron, yaitu NADH (Nikotin Adenin Dinokleutida) dan FAD (Flafin adenine dinukleotida) dalam bentuk FADH2. Kedua macam sumber elektron ini dibawa kesistem transfer elektron. Proses transfer elektron ini sangat komplek, pada dasarnya, elektron dan H+ dan NADH dan FADH2 dibawa dari satu substrak ke substrak yang lain secara berantai. Setiap kali dipindahkan, energi yang terlepas digunakan untuk mengikatkan fosfat anorganik (P) kemolekul ADP sehingga terbentuk ATP. Pada bagian akhir terdapat oksigen sebagai penerima, sehingga terbentuklah H2O. katabolisme 1 glukosa melalui respirasi aerobik menghasilkan 3 ATP. Setiap reaksi pada glikolisis, siklus krebs dan transport 14 elektron dihasilkan senyawa – senyawa antara. Senyawa itu digunakan bahan dasar anabolisme (Syamsuri, 1980). a) Sebanyak 10 molekul NADH2 dan 2 molekul FADH2 dihasilkan selama tahap glikolisis dan siklus Krebs. Seluruhnya akan memasuki reaksi redoks pada sistem transpor elektron. b) Tahapan : i) Mula-mula molekul NADH2 memasuki reaksi dan dihidrolisis oleh enzim dehidrogenase kembali menjadi ion NAD+ diikuti pelepasan 3 ATP, kemudian diikuti molekul FADH2 yang dihidrolisis oleh enzim flavoprotein kembali menjadi ion FAD+ dan menghasilkan 2 molekul ATP, keduanya juga melepaskan ion Hidrogen diikuti elektron, peristiwa ini disebut reaksi oksidasi. ii) Selanjutnya elektron ini akan ditangkap oleh Fe+++ sebagai akseptor elektron dan dikatalis oleh enzim sitokrom b, c, dan a. Peristiwa ini disebut reaksi reduksi. Reaksi reduksi dan oksidasi ini berjalan terus sampai elektron ini ditangkap oleh Oksigen (O2) sehingga berikatan dengan ion Hidrogen (H+) menghasilkan H2O (air). Hasil akhir dari sistem transpor elektron ini adalah 34 molekul ATP, 6 molekul H2O (air). Secara keseluruhan reaksi respirasi sel aerob menghasilkan 38 molekul ATP, 6 molekul H2O, dan 6 molekul CO2. 15 2.3.2 Respirasi Anaerob Respirasi anaerobik adalah reaksi pemecahan karbohidrat untuk mendapatkan energi tanpa menggunakan oksigen. Respirasi anaerob disebut fermentasi atau respirasi intramolekul. Respirasi anaerob dibedakan menjadi obligatif dan fakultatif, respirasi anaerob obligatif mutlak memerlukan oksigen sedangkan anaerob fakultatif dapat berlangsung tanpa atau dengan oksigen. Tujuan fermentasi sama dengan respirasi aerob, yaitu mendapatkan energy. Hanya saja energi yang dihasilkan jauh lebih sedikit dari respirasi aerob. Respirasi anaerob dapat berlangsung didalam udara bebas, tetapi proses ini tidak menggunakan O2 yang disediakan di udara. Fermentasi sering pula disebut sebagai peragian alcohol atau alkoholisasi. Pada respirasi aerob maupun anaerob, asam piruvat hasil proses glikolisis merupakan substrat ( Loveless, A.R, 1991). Respirasi anaerobik menggunakan senyawa tertentu misalnya asam fosfoenol piruvat atau asetal dehida, sehingga pengikat hidrogen dan membentuk asam laktat atau alcohol. Respirasi anaerobik terjadi pada jaringan yang kekurangan oksigen, akan tumbuhan yang terendam air, biji–biji yang kulit tebal yang sulit ditembus oksigen, sel–sel ragi dan bakteri anaerobik. Bahan baku respirasi anaerobik pada peragian adalah glukosa. Selain glukosa, bahan baku seperti fruktosa, galaktosa dan malosa juga dapat diubah menjadi alkohol. Hasil akhirnya adalah alcohol, karbon dioksida dan energi. Glukosa tidak terurai lengkap menjadi air dan karbondioksida. Reaksinya: C6H12O6 Ragi >> 2C2H5OH + 2CO2 + 21Kal Dari persamaan reaksi tersebut terlihat bahwa oksigen tidak diperlukan. Bahkan bakteri anaerobik seperti klostidrium tetani (penyebab tetanus) tidak dapat hidup jika berhubungan dengan udara bebas. Infeksi tetanus dapat terjadi jika luka tertutup sehingga memberi kemungkinan bakteri tambah subur (Syamsuri, 1980). Beberapa proses respirasi anaerob : 16 Fermentasi alkohol : Proses ini terjadi pada beberapa mikroorganisme seperti jamur ( ragi ), dimana tahapan glikolisis sama dengan yang terjadi pada respirasi aerob. Setelah terbentuk asam piruvat ( hasil akhir glikolisis ), asam piruvat mengalami dekarboksilasi (: sebuah molekul CO2 dikeluarkan ) dan dikatalisis oleh enzim alkohol dehidrogenase menjadi etanol atau alkohol dan terjadi degradasi molekul NADH menjadi NAD+ serta membebaskan energi/kalor. Proses ini dikatakan sebagai "pemborosan" karena sebagian besar energi yang terkandung dalam molekul glukosa masih tersimpan di dalam alkohol. Itulah sebabnya, alkohol/etanol dapat digunakan sebagai bahan bakar. Fermentasi alkohol pada mikroorganisme merupakan proses yang berbahaya bila konsentrasi etanolnya tinggi. Secara sederhana, reaksi fermentasi alkohol ditulis : 2CH3COCOOH asam piruvat ----------> 2CH3CH2OH + 2CO2 + 28 kkal etanol/alkohol Fermentasi asam laktat : Pada sel hewan ( juga manusia ) terutama pada sel-sel otot yang bekerja keras , energi yang tersedia tidaklah seimbang dengan kecepatan pemanfaatan energi karena kadar O2 yang tersedia tidak mencukupi untuk kegiatan respirasi aerob ( reaksi yang membutuhkan oksigen ). Proses fermentasi asam laktat dimulai dari lintasan glikolisis yang menghasilkan asam piruvat. Karena tidak tersedianya oksigen maka asam piruvat akan mengalami degradasi molekul ( secara anaerob ) dan dikatalisis oleh enzim asam laktat dehidrogenase dan direduksi oleh NADH untuk menghasilkan energi dan asam laktat. Secara sederhana reaksi fermentasi asam laktat ditulis sebagai berikut. 2CH3COCOOH ----------> 2CH3CHOHCOOH + 47 kkal Asampiruvat asamlaktat 17 2.4.Proses Fotorespirasi Proses yang disebut juga "asimilasi cahaya oksidatif" ini terjadi pada sel-sel mesofil daun dan diketahui merupakan gejala umum pada tumbuhan C3, seperti kedelai dan padi. Lebih jauh, proses ini hanya terjadi pada stroma dari kloroplas, dan didukung oleh peroksisom dan mitokondria. Secara biokimia, proses fotorespirasi merupakan cabang dari jalur glikolat. Enzim utama yang terlibat adalah enzim yang sama dalam proses reaksi gelap fotosintesis, Rubisco (ribulosa-bifosfat karboksilase- oksigenase). Rubisco memiliki dua sisi aktif: sisi karboksilase yang aktif pada fotosintesis dan sisi oksigenase yang aktif pada fotorespirasi. Kedua proses yang terjadi pada stroma ini juga memerlukan substrat yang sama, ribulosa bifosfat (RuBP), dan juga dipengaruhi secara positif oleh konsentrasi ion Magnesium dan derajat keasaman (pH) sel. Dengan demikian fotorespirasi menjadi pesaing bagi fotosintesis, suatu kondisi yang tidak disukai kalangan pertanian, karena mengurangi akumulasi energi. Jika kadar CO2 dalam sel rendah (misalnya karena meningkatnya penyinaran dan suhu sehingga laju produksi oksigen sangat tinggi dan stomata menutup), RuBP akan dipecah oleh Rubisco menjadi P-glikolat dan P-gliserat (dengan melibatkan satu molekul air menjadi glikolat dan POH). P-gliserat (P dibaca "fosfo") akan didefosforilasi oleh ADP sehingga membentuk ATP. P-glikolat memasuki proses agak rumit menuju peroksisoma, lalu mitokondria, lalu kembali ke peroksisoma untuk diubah menjadi serin, lalu gliserat. Gliserat masuk kembali ke kloroplas untuk diproses secara normal oleh siklus Calvin menjadi gliseraldehid-3-fosfat (G3P). 18 Gambar proses fotorespirasi 2.5 Faktor Faktor Yang Mempengaruhi Laju Respirasi Laju respirasi dapat dipengaruhi beberapa faktor antara lain : 1. Ketersediaan substrat Karbohidrat merupakan substrat respirasi utama yang terdapat dalam sel tumbuhan tinggi. Tumbuhan dengan kandungan substrat yang rendah akan melakukan respirasi dengan laju yang rendah pula. Demikian sebliknya bila substrat yang tersedia cukup banyak maka laju respirasi akan meningkat. 2. Ketersediaan oksigen Ketersediaan oksigen akan mempengaruhi laju respirasi, namun besarnya pengaruh tersebut berbeda bagi masing-masing spesies dan bahkan berbeda antara organ pada tumbuhan yang sama. 3. Suhu 19 Semakin tinggi suhu, semakin tinggi laju respirasi. Laju reaksi respirasi akan meningkat untuk setiap kenaikan suhu sebesar 10oC, namun hal ini tergantung pada masing-masing spesies. 4. Tipe dan umur tumbuhan Masing-masing spesies tumbuhan memiliki perbedaan metabolisme, dengan demikian kebutuhan tumbuhan untuk berespirasi akan berbeda pada masing-masing spesies. Tumbuhan muda menunjukkan laju respirasi yang lebih tinggi dibanding tumbuhan yang tua. Demikian pula pada organ tumbuhan yang sedang dalam masa pertumbuhan. 20 BAB III KESIMPULAN Respirasi atau pernafasan merupakan suatu proses pembongkaran (katabolisme atau desimilasi ) dimana energi yang tersimpan ditimbulkkan kembali untuk menyelenggarakan proses-proses kehidupan. Energi yang ditangkap dari proses oksidasi sempurna beberapa senyawa dalam proses respirasi dapat digunakan untuk mensintesis molekul lain yang dibutuhkan untuk pertumbuhan. Tahapan Respirasi antara lain respirasi aerob dan respirasi anaerob. Respirasi aerob yaitu respirasi yang menggunakan oksigen oksigen bebas untuk mendapatkan energi. Sedangkan respirasi anaerobik adalah reaksi pemecahan karbohidrat untuk mendapatkan energi tanpa menggunakan oksigen. Respirasi anaerob disebut fermentasi atau respirasi intramolekul. Laju respirasi dapat dipengaruhi beberapa faktor antara lain : Ketersediaan substrat, Ketersediaan oksigen, Suhu, Tipe dan umur tumbuhan. 21 DAFTAR PUSTAKA Champbell, N.A, dkk. 2002. “Biologi”- Edisi lima Jilid satu. Erlangga: Jakarta. Dwidjoseputro. 1986. Biologi. Erlangga. Jakarta. Jasin,Maskoeri.1989. “Biologi Umum Untuk Perguruan Tinggi”. Bina Pustakatama:Surabaya Kimball, J.W. 2002. Fisiologi Tumbuhan. Erlangga. Jakarta. Lakitan, Benyamin, 2007. Dasar-dasar Fisiologi Tumbuhan, PT. Raja Grafindo Persada, Jakarta Loveless, A.R. Gramedia 1991. Biologi Tumbuhan untuk Daerah Tropik, PT. Pustaka Utama, Jakarta Mertens, Thomas R, dkk.1966. “Laboratory Exercises in the principles of biology”. Burgess. Pratiwi, dkk.2003. Biologi SMA Jilid II. Erlangga. Jakarta. Salisbury, Frank and Ross, Cleon. 1995. Fisiologi Tumbuhan Jilid 2. Penerbit ITB: Bandung. Simbolon, Hubu dkk. 1989. Biologi Jilid 3. Erlangga. Jakarta. Yasa, I Beberapa Komang Jaya Santika. 2009. Respirasi Dipengaruhi oleh Faktor. www.idonbiu.com diambil tanggal 6 November 2009. 22