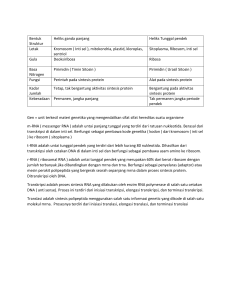
Transkripsi pada makhluk hidup 1. Proses transkripsi pada makhluk hidup prokariot Proses transkripsi dapat diartikan sebagai suatu segemen DNAyang ditranskripsikan untuk menghasilkan satu molekul RNA. Unit dari transkripsi mungkin dapat sama dengan gen suatu individu. Pada bakteri, proses transkripsi digunakan untuk membawa urutan pengkodean beberapa gen. Proses transkripsi dapat dibagi menjadi tiga tahap, yaitu inisiasi rantai RNA baru, perpanjangan rantai atau elongasi dan terminasi . Tahapan dari proses transkripsi dapat dilihat pada gambar 1.1 Gambar 1.1 Tahapan proses Transkripsi pada Prokariotik Sumber : Snustad,dkk (2012) Proses Transkripsi dilakukan katalisasi oleh suatu enzim yang disebut sebagai RNA polimerase. RNA polimerase pada suatu bakteri terdiri atas enam subunit, yaitu dua subunit α, dua subunit β, satu subunit ω, dan satu subunit σ. Suatu RNA polimerase dan subunit σ (holoenzim) akan berjalan sepanjang molekul DNA untuk menemukan lokasi awal dari transkripsi. Subunit σ berfungsi untuk membantu RNA polimerase mengenali suatu urutan tertentu pada molekul DNA yang dapat menandai tempat terjadinya awal transkripsi (awal suatu gen) yang disebut dengan promotor. Pada tahap ke 33 inisiasi, RNA polimerase dan subunit σ mengikat daerah promotor serta memisahkan untai ganda DNA agar inisiasi transkripsi dapat terjadi. Proses selanjutnya yaitu tahap elongasi, rantai RNA disintesis, subunit σ terlepas dari RNA polimerase dan transkripsi berlangsung terus-menerus hingga mencapai suatu daerah di akhir gen yang disebut terminator. Urutan terminator menandai tempat akhir transkripsi (akhir suatu gen). Pada bakteri E. Coli terdapat dua mekanisme terminasi, meliputi adanya protein ρ (rho) yang membantu melepaskan RNA tanpa bantuan protein ρ (rho-independen), pada daerah terminator membentuk seperti loop. Mekanisme protein ρ (rho-independen) dapat terlihat pada gambar 1.2 Gambar 2.2 Mekanisme protein ρ (rho-independen) Sumber : Snustad,dkk (2012) 2. Proses transkripsi pada makhluk hidup eukariot Umumnya organisme prokariotik memiliki struktur yang berbeda dengan organisme eukariotik. Pada gen organisme eukariotik, seluruh nukleotida menspesifikasi asam amino, sedangkan pada gen organisme eukariotik, hanya sebagian urutan nukleotida yang mengkode asam amino. Bagian dari gen yang mengkode asam amino disebut ekson, sedangkan yang tidak mengkode asam amino disebut intron. Ekson dan intron letaknya bergantian. Hasil dari proses transkripsi gen organisme prokariotik dapat ditranslasi menjadi protein, sedangkan pada proses transkripsi gen organisme eukariot, umumnya masih melalui proses tambahan untuk menghilangkan intron. Transkrip pada mRNA yang mengandung intron disebut transkrip primer (pre mRNA). Proses penghilangan intron terjadi di dalam nukleus dan dinamakan splicing. Transkrip bebas intron ini berperan sebagai mRNA yang ditranslasikan menjadi protein. mRNA eukariot mengalami modifikasi pada kedua ujungnya. Pada ujung 5’ ditambahkan beberapa residu guanilat yang termodifikasi menjadi 5’cap (5’-kepala). Sedangkan ujung 3’ dipotong dan ditambahkan 80-250 residu adenilat untuk membentuk suatu ekor poli-A. Hal ini dapat terlihat pada gambar 2.1 Gambar 2.1 Proses Posttranscriptional pada Eukariotik Sumber : Snustad,dkk (2012) Protein yang digunakan untuk proses inisiasi pada transkripsi, namun bukan merupakan bagian RNA polimerase dapat diartikan sebagai faktor transkripsi. Faktor transkripsi adalah aktivator yang berfungsi dalam sebuah promotor serta selalu berinteraksi secara langsung dengan RNA polimerase. Faktor transkripsi (TFs) berfungsi untuk mengenali situs pengikatan aktifator dalam daerah promotor dan menstimulasi RNA polimerase yang berikatan dengan promotor. Pengikatan RNA polimerase pada promotor bergantung pada protein yang disebut faktor transkripsi (TFs) seperti TFIID yang mengenali boks TATA. TFIID dapat diartikan sebagai faktor transkripsi pertama yang dapat berikatan dengan promotor pada daerah inisiasi. Promotor dan faktor transkripsi bekerja secara bersama saat menentukan RNA polimerase dan gen yang ditranskripsi. TFIID berikatan dengan boks TATA, Boks TATA adalah urutan nukleotida khas yang terdapat dalam daerah promotor sel eukariot. TFIID berfungsi untuk mengorganisasikan faktor transkripsi lain yang dibutuhkan untuk inisiasi sintesis RNA. Salah satu bagian dari TFIID adalah TFIIA. Faktor TFIIA merupakan suatu protein yang hanya diproduksi dalam sel eukariot yang mempunyai tipe jaringan tertentu. Pada proses transkripsi tingkat dasar membentuk suatu kompleks antara TFIIA dengan TFIIB, TFIID, TFIIE serta RNA polimerase II (Gambar 2.2). Gambar 2.2 Faktor transkripsi pada eukariotik Sumber : Alberts,dkk (2002) 3. Perbedaan transkripsi pada makhluk hidup prokariotik dan eukariotik Proses transkripsi merupakan langkah pertama ekspresi gen. Proses transkripsi pada organisme prokariotik memiliki beberapa perbedaan dengan organisme eukariotik. Perbedaan tersebut, meliputi transkripsi prokariotik terjadi di dalam sitoplasma, sedangkan transkripsi eukariotik terjadi di nukleus. Proses transkripsi pada prokariotik hanya melibatkan satu RNA polymerase, sedangkan pada transkripsi eukariotik melibatkan tiga jenis RNA polimerase. Selain itu, urutan mRNA prokariota yaitu polikistronik, sedangkan pada eukariota, urutan mRNA ialah monokistronik. Modifikasi pasca transkripsi hanya terjadi pada organisme prokariotik. Modifikasi Pasca Transkripsi dan Kode Genetik 1. Proses modifikasi pasca transkripsi pada makhluk hidup eukariotik Intron merupakan urutan yang ditemukan pada daerah yang tidak diterjemahkan dari beberapa gen. Urutan pengkodean dan nonkode yang tetap ada pada molekul mRNA dapat disebut dengan ekson atau urutan yang diekspresikan. Umumnya gen nukleus yang mengkode protein pada eukariotik multiseluler mengandung intron. Beberapa virus prokariota juga mengandung intron. "split" gen atau transkrip primer mengandung seluruh urutan gen serta urutan intron yang dipotong selama pemrosesan RNA. Gen yang berperan sebagai mengkode protein, mekanisme penyambungan harus bergabung dengan urutan ekson atau nukleotida tunggal yang berguna untuk memastikan bahwa kodon dalam ekson distal intron dibaca dengan benar. Namun, dalam transkrip utama gen nukleus, hanya intron berbeda urutan dinukleotida di ujung intron, yaitu untai DNA nontemplate yang setara dengan transkrip RNA. Gambar 1.1 Urutan untai DNA nontemplate Sumber : Snustad,dkk (2012) Pada proses transkripsi RNA, terdapat tiga tipe pemotongan intron, yaitu Intron precursor tRNA dipotong tepat saat pembelahan inti serta reaksi ligasi yang dikatalisis oleh enzim endonuklease. Kemudian, Intron pada Tetrahymena precursor rRNA yang dipindah ke reaksi khusus dan molekul RNA sebagai medianya. Tipe yang ketiga yaitu Intron dari hnRNA digabungkan melalui dua tahap reaksi yang dapat dipengaruhi oleh kompleks partikel ribonukleoprotein yang disebut dengan “spliceosomes”. Proses penyambungan precursor tRNA dapat dilihat hasilnya pada jamur ragi (Saccaromyces sp.). Pada proses penyambungan tRNA jamur ragi menggunakan sistem penyambungan secara invitro maupun penyambungan secara mutan. Proses pemotongan precursor tRNA terjadi dalam dua tahap. Tahap pertama yaitu ikatan membran nuclear menggabungkan endonuklease dan membuat pemotongan tersebut terjadi pada ujung intron. Kemudian, menggunakan suatu reaksi kompleks, ligase digabungkan dengan tujuan untuk menggabungkan 2 bagian tRNA, sehingga dihasilkan suatu molekul tRNA yang utuh. Pemotongan precursor menghasilkan ujung 5’-OH dan kelompok 2’-3’ phospat yang siklik pada ujung 3’. Tahap kedua pada proses ligasi melibatkan 4 reaksi yang terpisah, diantaranya yaitu penambahan kelompok phospat pada ujung 5’-OH. Reaksi ini membutuhkan aktifitas enzim kinase dan donor phospat. Kemudian, kelompok 5’ phospat diaktifkan dengan memindahkan AMP ke ujung molekul. Lalu ikatan siklik 2’-3’ phospat terbuka dikarenakan aktivitas enzim cyclic phosphodiesterase yang dapat menghasilkan 2’ phospat dan gugus 3’ hidroksil. Reaksi ligasi yang terakhir yaitu proses pemecahan gugus 3’-OH dengan cara melepaskan AMP. Mekanisme pemotongan intron pada sel mamalia memiliki edikit perbedaan dengan sel yang lain. Tipe selanjutnya yaitu penyambungan Autokatalitik dari Prekursor Rrna Tetrahymena. Intron pada precursor tRNA dari Tetrahymena dipotong tanpa menggunakan protein dan beberapa proses autokatalisis terjadi pada precursor rRNA, beberapa eukariot, precursor rRNA, tRNA, dan mRNA mitokondria. Pemotongan secara autokatalisis pada intron dalam precursor rRNA Tetrahymena membutuhkan transfer phospphodiester untuk memotong intron. Dua bagian intron yang telah dipotong akan dipindah ke ikatan phosphodiester yang lainnya. Aktivitas dari autokatalisis tergantung pada struktur intron atau struktur sekunder dari precursor tRNA. Gambar 1.2 Proses penyambungan Autokatalitik dari Prekursor Rrna Tetrahymena Sumber : Snustad,dkk (2012) Tipe pada penyambungan pre-mRNA, meliputi snRNAs, snRNPs, dan Spliceosome. Intron precursor pada inti sel dipotong melalui dua tahap seperti yang terjadi pada jamur ragi. Akan tetapi pada precursor inti intronnya tidak dipotong oleh enzim nuklease atau ligase. Intron tersebut dipotong oleh struktur protein yang disebut Spliceosome. Spliceosome mengandung suatu molekul RNA yang disebut snRNA. Tahap awal pemotongan terjadi pada ujung 5’ intron dan 2’-5’ phosphodiester dibentuk diantara posisi 5’-G yang ditempatkan dekat ujung3’ intron. Pada tahap kedua dimulai dengan kedua gen digabungkan oleh ikatan 3’-5’ phosphodiester dan intron yang telah dibentuk akan dilepaskan. Beberapa tahapan ini terjadi pada Spliceosome dan membutuhkan hidrolisis ATP. Molekul lain yang terkandung pada spliceosome yaitu molekul RNA yang disebut snRNP. Molekul snRNP akan ditambahkan pada proses pemotongan agar prosesnya berlangsung secara sempurna. Molekul snRNP U2 diikat pada suatu jaringan yang khusus dan membentuk suatu percabangan. Kemudian, snRNP U5 dan U4 atau U6 ditambahkan untuk menghasilkan spliceosome. Pada pembelahan ujung 5’ intron, snRNA U4 dilepaskan dari spliceosome. Setelah itu intron dipotong, dua bagian dari exon digabungkan dengan menyambungan 5’-3’ phosphodiester, sehingga mRNA yang sudah dipotong siap dipindah ke sitoplasma dan melanjutkan proses transkripsi. Proses tersebut terlihat pada gambar 1.2 Gambar 1.3 Peran snRNP yang mengandung snRNA pada penyambungan pra-mRNA Sumber : Snustad,dkk (2012) 2. Macam-macam kode genetik pada makhluk hidup Gen berperan untuk mengendalikan struktur polipeptida. Perantara mRNA yang berupa urutan empat basa dalam molekul mRNA dapat berperan untuk menentukan urutan asam amino dari polipeptida. Sifat kode genetik terdiri beberapa macam, yang meliputi kembar tiga nukleotida., kode genetik tidak tumpang tindih, kode genetik bebas koma, kode genetik yang mengalami degenerasi, kode genetik dipesan, kode genetik berisi kode start dan stop serta kode genetik hampir universal. Selama proses translasi terdapat dua puluh asam amino yang berbeda yang dimasukkan kedalam polipeptida. Pada tahun 1961, Francis Crick memiliki bukti yang mendukung kode triplet (tiga nukleotida per kodon).Crick melakukan analisis genetik mutasi yang diinduksi revertant di rII lokus bakteriofage T4 oleh proflavin kimia dan kemudian diisolasi. Revertant ini terbukti sebagai hasil dari terjadinya mutasi tambahan di lokasi terdekat dari pembalikan mutasi yang asli. Jika suatu kembar tiga nukleotida berurutan dalam mRNA berperan menentukan asam amino, maka setiap urutan nukleotida dapat dikenali atau dibaca selama penerjemahan dengan tiga cara berbeda. Misalnya, urutan AAAGGGCCCTTT dapat dibaca AAA, GGG, CCC, TTT atau A, AAG, GGC, CCT, TT dan AA, AGG, GCC, CTT, T. Crick melakukan percobaan kedua yang dilakukan dengan cara menambahkan dua macam basa nukleotida. Hasilnya yaitu asam amino yang ditranslasikan tidak sama dengan asam amino awal. Sedangkan pada percobaan Crick yang terakhir dilakukan dengan cara menambahkan tiga jenis basa nukleotida dan hasilnya yaitu asam amino hasil translasi yang berada disebelah tempat terjadinya mutasi sama dengan asam amino awal sebelum dilakukan mutasi. Setiap deretan tiga basa pada mRNA, mempunyai fungsi untuk mengkode satu asam amino spesifik disebut kodon. Terdapat empat basa pada nukleotida (A, G, C dan T) sehingga terdapat 43 = 64 kodon. Beberapa kodon memiliki fungsi khusus. AUG merupakan kodon inisiasi yang merupakan awal suatu polipeptida. Kodon UAA, UAG dan UGA tidak mengkode asam amino dan merupakan kodon terminasi atau stop kodon. Translasi terjadi dengan cara, setiap kodon dibaca secara berurutan dan tidak tumpang tindih. Kodon pertama pada gen menyediakan reading frame (pola pembacaan). Contoh urutan kode genetic dapat terlihat pada gambar 2.1 Gambar 2.1 Kode Genetik Sumber : Smith,dkk (2005) Pertanyaan dan Jawaban (Murniati Agustin/180341617524) 1. Sebutkan dan uraikan tiga macam RNA Polimerase pada proses transkripsi di eukariotik ? Jawab : - Terdapat tiga macam kompleks RNA polimerase, yang masing-masing diperlukan untuk proses transkripsi. Perbedaan ketiga macam RNA polimerase tersebut dapat diketahui melalui pemurnian menggunakan teknik kromatografi dan elusi pada suatu konsentrasi garam yang berbeda. Masing-masing RNA polimerase mempunyai sensitivitas yang berbeda terhadap toksin jamur α-amanitin, dan hal ini dapat dilakukan untuk membedakan aktivitasnya satu sama lain. Macam-macam kompleks RNA polymerase, meliputi : - RNA polimerase I (RNA Pol I) : Mentranskripsi sebagian besar gen rRNA. Enzim ini berada di dalam nukleoli dan tidak sensitif terhadap α-amanitin. - RNA polimerase II (RNA Pol II) : Mentranskripsi semua gen penyandi protein dan beberapa gen RNA nuklear kecil (snRNA). Enzim ini terdapat di dalam nukleoplasma dan sangat sensitif terhadap α-amanitin. - RNA polimerase III (RNA Pol III) : Mentranskripsi gen-gen tRNA, 5S rRNA, U6 snRNA dan beberapa RNA. Enzim ini terdapat di dalam nukleoplasma dan sedikit sensitif terhadap α-amanitin. 2. Mengapa RNA Polimerase tidak memiliki aktifitas Proofreading (Pembacaan kembali), seperti yang terdapat di DNA Polimerase ? Jawab : - Proofreading dapat diartikan sebagai sistem koreksi jika terjadi kesalahan dalam penggabungan aminoasil-tRNA pada ribosom. RNA polimerase tidak memiliki aktifitas proofreading (pembacaan kembali) eksonuklease 3’→5’, seperti yang dimiliki oleh DNA polimerase, sehingga terdapat sekitar satu kesalahan yang terjadi disetiap 104 -105 ribonukleotida yang dimasukkan selama transkripsi RNA. Di dalam sel diproduksi banyak salinan RNA dari satu gen, hal ini menyebabkan semua RNA segera di degradasi dan diganti, maka kesalahan pada molekul RNA tidak terlalu berpengaruh terhadap sel dibandingkan dengan kesalahan pada informasi yang tersimpan dalam DNA. 3. Mengapa proses transkripsi dan translasi pada prokariotik dapat terjadi secara bersamaan ? Jawab : - Proses transkripsi dan translasi pada prokariotik dapat berlangsung secara bersamaan yang diakibatkan pada prokariot tidak ada hambatan struktural sel, karena semua komponen transkripsi dan translasi terletak pada ruangan sitoplasma yang sama. Sebaliknya pada eukariotik, transkripsi berlangsung di dalam nucleus, sedangkan translasi berlangsung di dalam sitoplasma. Hal ini dapat diartikan bahwa translasi baru dapat dijalankan jika proses transkripsi sudah selesai dilakukan. Jeda waktu tersebut disebut fase pasca transkripsi. 4. Bagaimana proses terbentuknya DNA Supercoiling pada proses elongasi saat terjadi transkripsi ? Jawab : - Pada proses elongasi terdapat suatu hambatan yang dinamakan DNA Supercoiling. Hal ini dapat terjadi ketika gaya torsi terbentuk saat proses transkripsi DNA. Proses terbentuknya DNA Supercoiling diawali dengan DNA gyrase menggunakan ATP secara aktif untuk menciptakan supercoiling di polimerase. Supercoiling yang diciptakan berkebalikan belakang (negative RNA supercoiling) dengan supercoiling yang disebabkan oleh RNA polymerase, sehingga efeknya saling menghilangkan. Negative supercoiling dapat memfasilitasi proses pembukaan heliks dari DNA, sehingga dapat membantu proses transkripsi DNA pada bakteri. Perlu diingat bahwa DNA gyrase tidak ada pada organisme eukariotik. Untuk mengurangi atau mengatasi supercoiling, terdapat mekanisme berupa enzim topoisomerase pada eukariota dan bakteri. 5. Apa fungsi penguraian suatu kode genetik ? Jawab : - Fungsi dari penguraian suatu kode genetik ialah dapat menentukan kodon yang menentukan asam amino. Jumlah berapa banyak dari 64 kodon yang digunakan. Digunakan untuk mengetahui bagaimana kode diselingi serta penguraian kode genetik untuk mengetahui spesies berbeda menggunakan kodon sama atau berbeda. 6. Sebutkan dan uraikan tipe-tipe degenerasi kodon spesifik sebuah asam amino ! Jawab : - Proses degenerasi pada kodon dapat dibagi menjadi dua tipe, yang meliputi degenerasi parsial terjadi ketika basa ketiga, mungkin salah satu dari dua pirimidin (U atau C) maupun dua purin (A atau G). Selanjutnya yaitu degenerasi komplit, satu dari empat kodon mungkin menunjukkan posisi kodon ketiga, dan kodon akan tetap ditetapkan oleh asam amino yang sama. Misalnya, valin yang ditetapkan oleh GUU, GUC, GUA, dan GUG Daftar Pustaka : Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K, and Watter, P. 2002, Molecular Biology of The Cell, 4rd ed., New York : Garland Science. Gardner, 1991. Principles Of Genetics Eighth Edition. New York: John Wiley & Sons. Snustad, D. Peter dan Michael J. Simmons. 2012. Principles of genetics, sixth edition. United States of America: John Wiley and Sons, Inc. Smith, C., Marks, A.D., and Lieberman, M. 2005. Basic Medical Biochemistry, 2nd ed, Philadelphia : Lippincott William & Wilkins.