1. Proses floem loading
advertisement

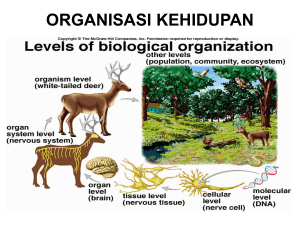
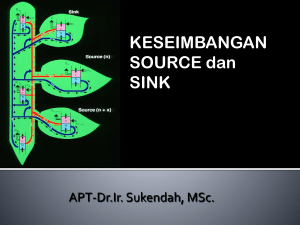
Pertemuan : Minggu ke 6 Estimasi : 150 menit Pokok Bahasan : Translokasi asimilat Sub pokok bahasan : 1. Proses floem loading – unloading 2. Konsep source dan sink 1. Mahasiswa Tujuan khusus : mengetahui dan dapat menjelaskan pengangkutan asimilat ke dalam floem 2. Mahasiswa mengetahui dan dapat menjelaskan proses pembongkaran asimilat dari floem 3. Mahasiswa mengetahui dan dapat menjelaskan konsep source dan sink dalam pengangkutan asimilat. Metode : Materi perkuliahan : Kuliah dan diskusi Reduksi karbon merupakan reaksi yang akan menghasilkan rangka karbon untuk pembentukan senyawa-senyawa organik lain, sehingga produk dari reaksi ini diperlukan oleh semua sel penyusun individu tumbuhan. Dengan demikian hasil fotosintesis akan segera ditranslokasi ke sel-sel yang memerlukan dengan melalui floem. Sukrose adalah bentuk utama karbohidrat yang ditranslokasikan keseluruh tubuh tumbuhan, dan amilum adalah bentuk karbohidrat yang tidak larut dan stabil ditempat penyimpanan makanan cadangan. Sintesis sucrose berlangsung di dalam sitosol, dan sintesis amilum dalam daun adalah didalam kloroplas (Gambar Prekursor untuk 22). sintesis kedua senyawa ini adalah triose-P, yang dihasilkan dan daur Calvin. Dalam sintesis sukrose, triose-P berdifusi menuju ke sitosol dan akan diubah menjadi glukose-P melalui fruktose 1,6 di-P. Selanjutnya menggunakan Fosfat (UTP) dengan Uridin Tri molekul glukose-P diaktifkan menjadi UDP-glukose untuk dapat bereaksi dengan fruktose-P menjadi sukrose. Enzim yang mengkataliser pembentukan sukrose ini memerlukan Mg2+ sebagai kofaktor sehingga ketersediaan unsur ini di dalam nutrisi tumbuhan sangatlah penting. Sintesis amilum di dalam kloroplas terjadi bila laju fotosintesis melebihi laju respirasi dan laju translokasi dari sel mesofil serta terjadi pada siang hari. Amilum terbentuk dari penambahan secara terus menerus molekul adenosin difosfoglukose (ADPG), yaitu senyawa hasil reaksi antara ATP dengan glukose-1-P. Reaksi pembentukan amilum ini dikataliser oleh enzim amilum sinthetase yang diaktifkan oleh ion K+. Inilah sebabnya tanaman yang kekurangan K hanya dapat mengakumulasi gula, bukan amilum. Pada malam hari amilum yang disimpan di dalam kloroplas ini akan keluar dari kloroplas setelah terlebih dahulu diubah menjadi sukrose. Gula yang ditranslokasikan adalah gula yang tidak dalam bentuk terreduksi, dan sukrose adalah bentuk yang paling banyak dijumpai. Selain sukrose, juga senyawa gula lainnya yang mengikat sukrose, seperti: rafinose, sukrose yang mengikat 1 mol. galaktose, stachiose, sukrose dengan 2 mol. galaktose, verbaskose, dengan 3 mol. galaktose, dan juga gula alkohol seperti mannitol dan sorbitol. Senyawa-senyawa tersebut akan diangkut ke sel-sel floem dan tulang daun yang terkecil. Pengangkutan ini kebanyakan dilakukan secara simplas, walaupun pada beberapa spesies pengangkutan juga dilakukan secara apoplas, dan merupakan pengangkutan jarak pendek. Dari penelitian yang pernah dilakukan ternyata bahwa pengangkutan sukrose secara apoplas menupakan pengangkutan aktif yang memenlukan ATP, atau dengan proton simport. Dalam pengangkutan sukrose secara simplastik, sel-sel mesofil daun yang merupakan sumber dari larutan yang akan ditranslokasikan (source) mempunyai konsentrasi yang lebih tinggi daripada komplek Gambar 2.3. Model penangkap polymer dalam pengangkutan fotosintat ke floem (dan buku Plant Physiology. Taiz & Zieger. 1998) sel floem dan sel pendamping (sel intermedier) karena sukrose yang terakumulasi didalam sel intermedier akan bereaksi dengan galaktose menjadi raffinose dan stachiose (Gambar 23). Kedua molekul ini akan diangkut ke sel-sel floem, sehingga walaupun konsentrasi gula didalam sel-sel floem dan sel pendamping meningkat tetapi karena ukuran molekulnya besar maka gula tersebut tidak dapat diangkut kembali ke sel-sel mesofil (menurut “polymer trapping model” oleh Robert Turgeon). Pengangkutan di dalam floem merupakan pengangkutan jarak jauh, dan tidak dapat dikatakan sebagai pengangkutan keatas atau kebawah, karena pengangkutan ini dan sel-sel mesofil daun sebagai sumber atau source menuju ke tempat metabolisme atau penyimpanan (sink). Karena pengangkutan mi melalui pembuluh floem, maka “source” akan lebih dulu memberikan fotosintat kepada “sink” yang mempunyai hubungan langsung dengan pembuluh floem. Penelitian dengan menggunakan 14 CO2 yang diberikan pada daun tanaman beet gula (Beta vulgaris) no. 14 (Gambar 24), yang menjadi “source” akan memberikan fotosintatnya kepada daun pucuk (no. 1) dan daun no. 6, yang menjadi “sink” serta terdapat pada sam garis duduk daun (orthostich) dengan “source”. Ketika daun-daun (B) yang berlawanan arah dengan daun “source” (daun no. 10) dihilangkan 24 jam sebelum pemberian karbon dioksida yang radioaktif maka daun-daun yang terdapat pada dua sisi orthostich dan menjadi “sink”, akan mendapatkan fotosintat secara merata. Gambar 24. Diagram distribusi radioaktivitas dan satu daun yang menjadi sumber pada daun-daun yang menjadi “sink” (dan Plant Physiology. Taiz & Zieger. 1998). Di tempat penimbunan (sink), gula yang terdapat di dalam sel-sel floem akan dipindahkan ke sel-sel penerima yang strukturnya sangat bervariasi, mulai dan sel-sel meristem apikal, sel-sel penyimpan makanan cadangan di akar, batang, biji, dan buah. Dalam proses pemindahan dan komplek sel floem dan sel pendamping ke sel-sel penerima dapat dilakukan secara simplas atau secara apoplas. Komplek sel floem dan sel pendamping dalam hal mi menjadi sumber dan mempunyai konsentrasi gula yang tinggi, sedang sel-sel penerima mempunyai konsentrasi gula yang rendah. Bila sel-sel penerima akan menggunakan fotosintat untuk metabolisme, maka sukrose segera diubah menjadi glukose dan fruktose yang dapat langsung digunakan sebagai substrat respirasi. Bila sel penerima adalah sel penimbun, maka akan terjadi akumulasi sukrose, sehingga transfer secara simplas dapat dibantu dengan transfer secara apoplas untuk mencegah arus balik sukrose dan sel-sel penerima ke sel-sel floem.