Universitas Gadjah Mada
advertisement

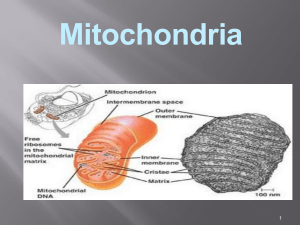
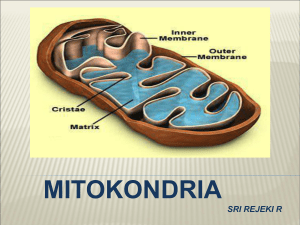
IV. DISTRIBUSI PROTEIN KE MITOKONDRIA DAN KLOROPLAS 1. DISTRIBUSI PROTEIN KE MITOKONDRIA DAN KLOROPLAS Mitokondria dan kloroplas adalah organela yang berisi DNA, ribosom dan semua komponen yang diperlukan untuk sintesis protein. Bagaimanapun juga, protein yang dikode dan dirakit di dalam kedua organela tersebut hanya merupakan sebagaian kecil dan total protein yang terdapat di dalam organela. Sebagian besar protein tersebut dikode dan inti dan disintesis di dalam sitoplasma. Protein yang berasal dari sitoplasma yang terdapat di dalarn organela tersebut, mungkin diarahkan ke satu dan berbagai likasi. Protein yang dikirim ke mitokondria termasuk membran luar, ruang intermembran, membran dalam dan matriks. Kloroplas sedikitnya mempunyai 6 lokasi sebagai tujuan protein dan sitoplasma ialah membran luar, kompartemen intermembran, membran dalam, stroma atau kompartemen dalam yang analog dengan matriks pada mitokondria, membran tilakoid dan kompartemen di dalam yang dibungkus membran tilakoid. Penelitian yang mengarah pada mekanisme impor protein ke mitokondria dilakukan lebih banyak dibanding dengan penelitian yang sama pada kloroplas. Tampaknya mekanisme pengiriman protein dan sitoplasma ke kedua organela tersebut berlangsung serupa, meskipun terdapat perbedaan jumlah membran suborganela dan kloroplas. Penelitian yang dilakukan oleh G. van den Broeck dkk. Mempelajari arahan sinyal subunit kecil dan ribulosa-1,5-bis-fosfat (RuBP) karboksilase dari sitoplasma ke kloroplas. Protein hibrid terbentuk dengan cara memasukkan sinyal RUBP karboksilase ke N-terminal dan enzim marker bakteria. Apabila enzim bakteri yang mendapat tambahan sinyal dan RuBP karboksilase dimasukkan ke dalam sel tanaman tembakau, maka neomycine phophatase akan mengikuti perjalanan yang benar ke arah kloroplas. Percobaan serupa dilakukan dengan cara memindahkan sinyal pengarah mitokondria ke protein sitoplasma bakteri yang akan membawa protein ke dalam mitokondria sel eukariot. Semua protein yang diimpor mitokondria dan kloroplas yang disintesis dalam sitoplasma dapat menyusup ke dalam membran dengan cara pasca translasional. Di samping kemampuan protein menyusup dengan cara tersebut, Universitas Gadjah Mada banyak protein yang dirakit oleh ribosom melintas membran secara kotranslasional. Pada awalnya protein menempel pada membran luar organela akibat interaksi antara sinyal dan membran luar. Proses tersebut diprakarsai oleh faktor-faktor reseptor protein sitoplasmik yang terdapat dalam membran organela. Faktor sitoplasmik yang terlarut mungkin bertindak sebagai chaperones yang menjaga protein organela tidak melipat. Kondisi tersebut diperlukan untuk penetrasi membran. Juga diperoleh kepastian, bahwa energi hasil hidrolisis ATP digunakan untuk menjaga agar molekkul tetap dalam keadaan tidak melipat. Tampaknya protein yang dikode dalam genom mitokondria dan kloroplas terdapat di dalam membran-dalam mitokondria dan membran tilakoid kloroplas. Polipeptida yang disintesis umumnya merupakan subunit dan kompleks protein yang dikode oleh gen inti dan diimpor dan sitoplasma. a. Distribusi protein ke mitokondria Rangakaian sinyal peptith (sinyal N-terminal) protein yang mengarah ke mitokondria, ternyata lebih panjang dari pada sinyal N-terminal yang digunakan untuk pengiriman protein ke RE. Panjang sinyal dalam protein impor tersebut mencapai 70 buah asam amino. Struktur N-terminal pengarah protein ke mitokondria disajikan pada Gambar 8.23. Semua sinyal mitokondria berisi daerah panjang dengan residu yang bermuatan positif dekat dengan N-terminal. Residu yang bermuatan positif tersebut termasuk Lys dan Arg yang letaknya terpisah oleh rangkaian asam amino netral sebanyak 4-8 buah. Pada daerah sentral yang letaknya di belakang segmen awal yang bermuatan, banyak sinyal mitokondria yang berisi rangkaian residu asam amino polar sekitar 20-25 buah yang cukup Universitas Gadjah Mada panjang untuk melintas rentang membran rangkap dan fosfolipida. Rangkaian asam amino membentuk segmen yang berongga, sehingga sisi yang satu polar dan yang lain non-polar apabila molekul tersebut membetuk struktur α atau β heliks. Heliks tersebut terbentuk apabila protein membran berupa multisegmen transmembran. Pengaturan segmen protein ini akan membentuk kanal dan protein yang menembus membran, sehingga memungkinkan transpor protein dengan rantai samping yang bermuatan dan polar melintas membran. Protein yang diimpor ke dalam matriks mitokondria biasanya berlangsung satu atau dua menit sesudah lepas dari poliribosom. Apabila Protein yang diimpor ke dalam matriks mitokondria biasanya berlangsung satu atau dua menit sesudah lepas dari poliribosom. Apabila dalam protein tersebut hanya terdapat satu sinyal N-terminal, maka sesudah protein yang diimpor masuk ke dalam matriks mitokondria, sinyal terminal (sinyal peptida) akan dipotong oleh protease spesifik yang dikenal dengan nama snyal peptidase di dalam matriks. Kemungkinan segmen tersebut akan mengalami degradasi menjadi asam amino dalam kompartemen tersebut. Apabila di samping sinyal peptidase pertama terdapat sinyal kedua yang merupakan untai asam amino hidrofobik, pemotongan sinyal pertama akan menjadikan sinyal kedua pada ujung N-terminal dan protein. Sinyal kedua Universitas Gadjah Mada akan mengarahkan molekul protein menyusup melintas membran dalam. Tergantung pada rrangkaian informasi lainnya, protein akan tinggal dalam membran dalam atau melintas membran dalam dan menuju ke ruang intermembran. Secara skematis proses tersebut dilukiskan pada Gambar 8.25. Percobaan dengan menggunakan isolat mitokondria yang diinkubasikan pada suhu 5 °C, protein hanya sebagian yang masuk ke organel tersebut dan hampir semua rantai polipeptida berada di luar. Apabila suhu dinaikkan menjadi 37°C, maka translokasi protein menjadi sempurna. Tampak molekul protein akan Iangsung menembus kedua membran mitokondria lewat satu titik kontak. Peneliti N. Pfanner, W. Neupert, G. Schatz dkk. menemukan banyak protein yang dapat bertindak sebagai chaperones, juga reseptor dan lain-lain elemen mitokondria yang diperlukan dalam mekanisme impor yang perannya dapat dilihat pada Gambar 8.26. Protein dalam sitoplasma dengan sinyal N- Universitas Gadjah Mada terminal mengarah ke mitokondria bersama dengan chaperones yang menjaga polipeptida agar tidak melipat. Sinyal N-terminal yang tidak melipat dengan konfigurasi yang memungkinkan protein menyusup ke membran mitokondria. Ikatan antara protein dengan chaperone memerlukan energi dari ATP. Protein dan reseptor yang merupakan protein transparan mitokondria yang disebut MOM (mitochondrian outer membrane) akan berikatan dengan reseptor peptida. Protein akan bergerak menembus membran mitokondria secara simultan yang prosesnya dibantu oleh protein membran kedua ialah General Insertion Protein (GIP). Pada saat protein berada dalam matriks, sinyal N-terminal dilepas oleh sinyal peptidase matriks, Mitochondrial Processing Peptidase (MPP) yang diaktifkan oleh potein kedua ialah Processing Enhancing Protein (PEP). Apabila protein sinyal kedua berada dekat protein sinyal pertama, maka sinyal kedua akan berikatan dengan chaperone lain dalam matriks. Chaperon hsp60 menjaga protein dalam konformasi tertentu agar sinyal kedua dapat menyusup ke arah luar membran-dalam mitokondria. Diperkirakan dalam membran-dalam terdapat Universitas Gadjah Mada protein reseptor sinyal kedua. Setelah protein masuk ke dalam ruang intetrmembran, sinyal kedua dilepas. Tidak semua protein dalam membran, matriks dan ruang intermembran sebagai hasil mekanisme sinyal. Sitokrom c adalah protein perifer yang menempel pada permukaan luar membran-dalam mitokondria yang masuk ke dalam organela tersebut tanpa rangkaian sinyal yang dapat dilacak dan tidak diketahui interaksi dengan reseptor atau elemen membran lainnya. Protein tersebut mungkin secara spontan melipat dan otomatis menuju ke lokasi terakhirnya. Tampaknya banyak atau semua rangkaian dan konformasi lipatan bertindak sebagai rangkaian sinyal. b. Distribusi Protein ke Kloroplas Transpor protein ke kloroplas dalam beberapa hal serupa dengan transpor protein ke mitokondria. Persamaannya adalah proses berlangsung secara pasca translasional, memerlukan energi dan keduanya memerlukan sinyal peptida amino-terminal yang hidrofilik yang dilepas sesudah digunakan. Dalam sel tanaman ditemukan mitokondria dan kloroplas bersama-sama dan meskipun kedua proses translokasi serupa, tetapi protein dapat memilih arah yang tepat. Kloroplas mempunyai kompartemen lain yang disebut tilakoid. Banyak protein kloroplas termasuk protein subunit dalam sistem fotosintetik dan ATP sintetase diimpor tilakoid dan sitosol. Protein diangkut ke tujuan akhir dengan dua langkah ialah melewati membran rangkap kloroplas dan masuk ke dalam stroma. Kedua protein mengalami translokasi ke dalam membran tilakoid atau masuk ke dalam ruang tilakoid. Kebanyakan sinyal pada kloroplas tidak bersifat amfipatik. Ujung belakang sinyal merupakan tempat pemotongan oleh sinyal peptidase. Proses translokasi protein ke dalam tilakoid disajikan pada Gambar 8.27. Universitas Gadjah Mada Sinyal N-terminal yang mengarahkan protein ke kloroplas panjangnya berkisar antara 30 sampai mendekati 100 asam amino dan kebanyakan berisi sikitar 50 residu. Penelitian K. Keegstra, S. Smeekens dan P. Weisbeck menyatakan bahwa protein yang diarahkan ke kedua membran dan stroma berisi sinyal N-terminal yang banyak mengandung Ser dan Thr, tetapi sedikit atau tidak terdapat residu asam amino asam. Sinyal kedua dari beberapa protein kloroplas mempunyai sifat yang serupa dengan sinyal prokariot yang disebut sinyal pengarah-stroma. Sinyal kedua dimulai dengan bebrapa residu bermuatan yang diikuti oleh rangkaian teras yang hidrofobik dan diakhiri dengan sinyal pemotongan yang serupa dengan sinyal prokariot. Tergantung pada informasi sinyal berikutnya, protein mungkin tinggal dan tertanam dalam membran tilakoid atau masuk ke dalam ruang tilakoid. Sinyal peptidase kedua yang terikat pada membran tilakoid, melepas sinyal kedua yang sama dengan sinyal kedua dan mitokondria yang mengarahkan protein membran-dalam atau ruang intermembran. Kloroplas juga berisi protein yang dikode dan disintesis di dalam stroma dan dialamatkan ke membran atau kompartemen tilakoid. II. DISTRIBUSI PROTEIN KE MEMBRAN INTl SEL Inti sel dibungkus oleh 2 macam membran ialah membran-dalam inti sel yang berisi protein spesifik yang merupakan tapak pengikat untuk lamina inti dan berhubungan dengan kromosom dan RNA inti. Membran-luar inti yang serupa dengan membran retikulum endoplasmik adalah pengembangan dari organela Universitas Gadjah Mada tersebut. Protein yang disintesis dalam ribosom diangkut ke dalam ruang antara membran-dalam dan membran-luar inti sel (ruang perinuklear). Proses tersebut disajikan pada Gambar 8. 28. Protein dalam inti seperti histon, DNA dan RNA polimerase dan protein gen regulator diimpor dari sitosol. Molekul tersebut harus melewati membran-luar dan membran-dalam agar dapat masuk ke dalam lumen inti. Sinyal pengarah protein ke dalam intl sel berlainan dengan sinyal pangarah-RE atau pengarah organela. Sinyal pengarah-inti tidak hanya terdapat pada ujung N-terminal, tetapi juga pada berbagai tempat dalam molekul protein. Sinyal inti harus menonjol ke permukaan dan umumnya tetap tinggal dalam protein setelah mencapai sasaran. Sinyal tersebut biasanya pendek dan terdiri atas asam amino basa seperti Mg dan Lys yang menyusun Muster. Susunan molekul tersebut diselingi oleh asam amino netral atau hidrofobik. Prolin atau asam amino asam dengan rantai samping yang besar terdapat dekat dengan sinyal. Universitas Gadjah Mada Protein jnti dirakit oleh ribosom bebas di da1m sitosol dan dikirim ke inti secara pasca translasional lewat kompleks porus ini. Dua tahap reaksi yang terlibat dalam transpor protein ke inti sel ialah: 1. Penempelan protein ke porus kompleks dan mengikat sinyal inti. 2. Penetrasi protein lewat porus inti. Tahap kedua memerlukan ATP. III. TRANSPOR PROTEIN KE DALAM PEROKSISOM Peroksisom yang juga disebut miicrobodies berbeda dengan mitokondria dan kloroplas, karena peroksisom hanya terdiri atas satu lapis membran dan tidak berisi DNA atau ribosom. Semua protein di dalam peroksisom berasal dari sitoplasma dengan menggunakan ribosom bebas (tidak terikat pada RE). Peroksisom sel hati mempunyai garis tengah sekitar 0,5 µm yang berisi paling sedikit 3 macam enzim ialah D-asam amino oksidase, urat oksidase dan katalase. Peroksisom juga banyak menggunakan oksigen untuk melepas hidrogen dan substrat organik spesifik (disim disebut R) dalam suatu reaksi oksidatif yang menghasilkan hidnogen peroksida. Reaksi oksidatif antara substrat dengan oksigen sebagai berikut: RH2+O2 R+H2O2 Semua protein baik di dalam lumen maupun membran peroksisom berasal dan sitoplasma. Katalase yang banyak dipelajari merupakan tetramer protein yang berisi haem yang diimpor dan sitosol dalam bentuk monomer. Molekul enzim tersebut tidak berisi sinyal pengarah yang dilepas sesudah digunakan, tetapi mempunyai sinyal yang mengarahkan protein ke peroksisom. Kemungkinan sinyal terdiri atas 3 buah asam amino yang terletak dekat karboksil terminal yang ditemukan dalam banyak molekuk protein peroksisom. KEPUSTAKAAN Albert, B., D. Bray, J. Lewis, M. Raff, K. Robefts, and J.D. Watson. 1991. A Molecular Biology of the Cell. Gerland Publihing, Inc. New York and London. Berg, J.M., J.L. Timoczko, L. Stryer. 2002. Biochemistry. Fifth Edition. W.H. Freeman and Company. New York. Horton, H.R., L.A. Moran., R.S. Ochs, J.D. Rawn and KG. Scrimgeour. 1996. Principles of Biochemistry. Second Edition. Prentice-Hall International., Inc. New Jersey. Universitas Gadjah Mada Stryer, L. 1988. Biochemistry. Third Edition. W.H. Freeman Company. New York. Wolfe, S.L. 1993. Molecular and Cellular Biology. Wardsworth Publishing Company. Belmont, California. Universitas Gadjah Mada