Studi Pendahuluan Resistensi Udang Windu Penaeus monodon
advertisement

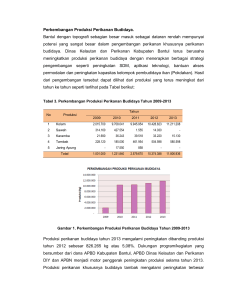
Aquacultura Indonesiana (2004) 5(1): 109–114 ISSN 0216–0749 Studi Pendahuluan Resistensi Udang Windu Penaeus monodon Terhadap Whitespot Baculovirus Hari Suprapto1 dan Muhammad Murjani2 1 PS Perikanan, Fakultas Kedokteran Hewan, Universitas Airlangga Jl. Mulyorejo Kampus C Surabaya 60115, T/F (031)-599-2785/599-3015 2 Balai Budidaya Air Payau Situbondo Jl. Pecaron Situbondo Abstract Hari Suprapto and Muhammad Murjani. 2004. A Prilimanary study of the black tiger shrimp (Penaeus monodon) resistance on whitespot baculovirus (WSBV). Aquacultura Indonesiana, 5(3): 109–114. The WSBV disease caused the decreased shrimp production. The aim of the experiment was to produce the resistant shrimp against WSBV infection with selective breeding. The normal size of nucleocapsid between 78–90 nm (diameter) and 330–350 nm (length) and founded in the hypertrophied of nucleus and cytoplasm. The protein number of virus for the artificial infection is W1 (0.234 mg/mL), W2 (0.117 mg/mL), W3 (0.029 mg/mL) and W4 (0.058 mg/mL). The doses for the artificial infection are 0.029 mg/mL proteins, caused the cumulative mortality 63% and the SR is 33%. The high mortality occurred in male than in female in the molting shrimp. On WSBV infection caused the higher production of proPO compared to control shrimp. Production of proPO similar to reactive oxygen species in which in the fourth weeks relatively higher compared to control shrimp. The higher of proPO production due to WSBV infection in shrimp, as a proved the shrimp still live till to date. The stimulation of shrimp against WSBV can increased the production of reactive oxygen species from hemocyte. The results are constant almost 1 month after infection. Tentatively conclusion is shrimp have raised the resistance against WSBV because the survival rate of infected shrimp reaches 33 percent. Keywords: Penaeus monodon; Resistence; WSBV Abstrak Penyakit WSBV menyebabkan menurunnya produksi udang. Tujuan penelitian adalah untuk menghasilkan resistensi udang terhadap serangan infeksi WSBV melalui seleksi breeding. Ukuran normal virionnya antara 78–90 nm (diameter) dan 330–350 nm ( panjang) dan ditemukan pada pembengkakan inti sel dan sitoplasma. Jumlah protein dari sediaan virus yang digunakan adalah W1 (0,234 mg/mL), W2 (0,117 mg/mL), W3 (0,029 mg/mL) dan W4 (0,058 mg/mL). Dosis untuk infeksi 0,029 mg/mL protein menyebabkan kematian kumulatif 63% dan SR 33%. Kematian banyak terjadi pada jantan dibanding betina pada masa udang molting. Infeksi WSBV menyebabkan produksi proPO lebih tinggi dibandingkan udang kontrol. Produksi proPO mirip dengan reactive oxygen species di mana sampai 4 minggu relatif lebih tinggi dibandingkan udang kontrol. Produksi proPO yang tinggi berkaitan dengan infeksi WSBV pada udang, akan tetapi udang yang terinfeksi WSBV masih hidup hingga saat ini. Stimulasi atau infeksi udang windu dengan WSBV dapat meningkatkan produksi reactive oxygen species dari hemocyte. Hasilnya konstan selama hampir 1 bulan setelah infeksi. Kesimpulan sementara adalah resistensi udang yang terkena serangan WSBV menyebabkan kelulushidupan udang terinfeksi mencapai 33%. Kata kunci: Penaeus monodon; Resistensi; WSBV Pendahuluan White Spot Baculovirus (WSBV) adalah penyakit yang sangat mematikan bagi udang dan kematian mutlak terjadi pada 3–6 hari sesudah infeksi. WSBV menyerang cr ustacea yang mempunyai nilai ekonomis tinggi (Flegel et al., 1992; Lightner, 1994; Takahashi et al., 1994; Chou et al. 1995). Chen (1992), melaporkan udang Penaeus chinensis yang terinfeksi di China dengan mortality rate 5%. Sedangkan Chou et al. (1995) melaporkan White spot disease di Taiwan disebabkan oleh baculovirus. Pada tahun 1993, kematian massal pada kuruma shrimp Penaeus japonicus juga terjadi di Jepang.Virusnya yang menyebabkan penyakit berbentuk bacilliform Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 109 Aquacultura Indonesiana, Vol. 5, No. 3, Desember 2004 : 109–114 (Takahashi et al., 1994), dan dinamai PRDV (Penaeid Rod-shaped DNA Virus). Virion berukuran diameter of 87+7 nm dan 330+20 nm in panjang (Chou et al., 1995). WSBV juga menyebabkan kematian udang windu di Indonesia dan diduga merupakan salah satu penyebab menurunnya budidaya udang. Ukuran virionnya adalah 78–90 nm (diameter) and 330–350 nm (panjang) (Suprapto, 2003). Pada udang yang terinfeksi akan menyisakan sekitar 5–10% yang hidup, dan ini merupakan fenomena yang sangat menarik, sehingga penelitian ini dilakukan berdasarkan fenomena di atas untuk mengetahui mengapa udang bisa resisten terhadap infeksi WSBV. Materi dan Metode Isolasi Virus campuran tersebut ditutup dengan satu tetes mineral oil. Primer yang digunakan yaitu: F002 GATGAGACAGCCCAAG TTGTTAAAC R002 CGAAATTCCATCACTGTAATTGCTTG Sedangkan kontrolnya yaitu: β–Actin– FCCCAGAGCAAGAGAGGTA Actin–R3 GCGTATCCTTCGTAGATGGGC Amplifikasi dilakukan dalam DNA thermal cycles 95ºC selama 5 min dan kemudian 50 cycles (95ºC selama 30 detik, 55ºC selama 30 detik dan 72ºC selama 1 menit) dan akhirnya 5 menit perpanjangan pada suhu 72ºC sesudah 50 cycles. Hasilnya akan dianalisis pada 1,5% agarose gel yang mengandung ethidium bromide 0,5 µg/mL. Infeksi Terhadap Udang dan Pemilihan yang Resisten Udang yang terinfeksi WSBV diambil hepatopancreas (HP) dan saluran pencernaannya (SP) ditimbang dan digerus dengan mortar. Kemudian ditambahkan L–15 yang mengandung Fetal bovine serum (FBS) dengan rasio 9:1, dipusingkan pada 2,000 g selama 10 min dan supernatant difiltrasi dengan 0,20 µm. Sediaan virus disimpan pada suhu –80ºC sampai digunakan untuk penelitian. Sediaan virus ini tahan disimpan sampai beberapa tahun pada suhu –80ºC karena pada suhu penyimpanan tersebut virus masih sangat ganas jika diinfeksikan pada udang. Pengukuran konsentrasi homogenat WSBV dilakukan dengan metode Lowrie (1985). Jumlah virus yang akan disuntikkan ke udang harus di atas LD50 atau dalam lethal doses, sehingga diharapkan hampir semua mati dan kontrol tidak diberi virus. Sedangkan untuk udang yang selamat akan dipelihara dan dibuat induk untuk mendapatkan telur dan ditetaskan lagi. Uji tantang tersebut diulangi lagi terhadap anak udang yang kita sebut dengan Generasi Pertama (F1). Anak udang tersebut dibesarkan kemudian dilakukan lagi uji tantang dan yang selamat akan dipelihara untuk dijadikan induk dan anaknya menjadi generasi kedua (F2) dan proses tersebut dilakukan berulang. Ekstraksi WSBV DNA dari udang Hemolymph untuk mengetahui produksi Reactive oxygen species (ROS) dan aktifitasi prophenoloxidase (proPO) diambil dari segmen ke enam udang dengan 26 gauge needle pada 2 mL syringe yang mengandung 0,2 mL anticoagulant (0,01 M Tris–HCl; 0,25 M Sucrose; 0,1 M Sodium citrate; pH 7,6). Hemolymph yang diisolasi kemudian disaring pada 300 x g selama 10 min pada suhu 4ºC dan supernatant yang dihasilkan dibuang, pellet yang didapatkan diencerkan pada L– 15. Hemocyte lysate supernatant (HLS) dipersiapkan menurut metode Sung et al. (1994), dan hemocyte lysed dilakukan dengan sonifikasi. Usus udang diambil dan dicacah kemudian dihomogenisasi dalam lysis buffer (50 mM Tris– HCl, 10 mM NaCl, 2% SDS, 10 mM EDTA, proteinase K 100 µg/mL) selama 3 at 56ÚC). DNA diekstraksi 1 jam dengan menggunakan phenol seperti yang diuraikan oleh Sambrook et al. (1989). DNA dari udang dipakai sebagai template untuk amplifikasi PCR. Amplifikasi PCR Amplifikasi PCR dilakukan dalam 50µl campuran reaksi yang mengandung 10mM Tris–HCl (pH 8.0), 50 mM KCl, 1.5 mM MgCl2, 0.1% Triton X–100, 0.2 mM tiap dNTP, 100 pmolof, 1 unit Taq DNA polymerase dan 50–100 ng DNA templates, Isolasi Hemolymph Produksi Reactive Oxygen Species Produksi reactive oxygen species diukur dengan metode nitroblue tetrazolium (NBT) (Song dan Hiseh, 1994), dengan cara 100 ¼L dari suspensi Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 110 Studi pendahuluan resistensi udang windu Penaeus monodon (Hari Suprapto dan M. Murjani) Tabel 1. Deteksi WSBV dari udang windu P. monodon dengan PCR dan TEM yang diinfeksi dengan WSBV secara buatan. hemocyte pada L–15 kemudian dipindahkan pada 96 sumuran dan kemudian dilakukan inkubasi selama 30 min pada suhu 37°C. Sesudah inkubasi, NBT dibuang dan 100 ¼L metanol ditambahkan pada setiap sumuran. Metanol kemudian diambil dan sumuran dicuci sebanyak 3 kali dengan 70% metanol, sesudah kering dilapisi dengan larutan 120 ¼L KOH (2M) dan 140 ¼L DMSO untuk melarutkan cytoplasmic formazan. Optical density diukur pada 630 nm menggunakan microplate reader. Jenis infeksi PCR Monodon Monodon Monodon Buatan Alami Kontrol + + - TEM + + - sitoplasma (Tabel 1). Percobaan penularan virus WSBV menyebabkan kematian pada P. vannamei, Penaeus merguiensis dan P. monodon. Partikel virus berbentuk single atau multiple sperical inclusion bodies yang menyebabkan pembengkakan inti sel. Kelulushidupan virus pada air tambak belum diketahui dengan pasti, tetapi SEMBV dapat hidup pada air laut selama 5 hari dengan suhu 28°C, tetapi hasil isolasi virus yang dsimpan pada suhu –80°C tetap virulen seperti sediakala. Untuk mengetahui jumlah dosis virus yang tepat yang digunakan untuk infeksi udang windu, sediaan virus diukur jumlah proteinnya (Lowry et al., 1985). Cara ini digunakan untuk menggantikan TCID 50 yang biasa digunakan untuk menghitung dosis infeksi virus, untuk penghitungan dosis infeksi ini kesulitan karena sampai saat ini tidak ada kultur sel krustasea. Penggunaan dosis semakin besar (W1) mortalitas udang windu semakin tinggi (Gambar 1), begitu berurutan sampai dengan jumlah yang terkecil (W3). Infeksi yang dilakukan pada udang putih P. vannamei dengan mortalitas untuk kedua dosis W1 dan W2 mencapai 100%. Dari hasil tersebut dosis infeksi yang digunakan adalah W3 dengan jumlah 0,029 mg/mL protein, dengan dosis Aktifitas Prophenoloxidase Aktifitas prophenoloxidase diukur dengan menggunakan metode Sung et al. (1994). Sebanyak 50 ¼L HLS dipindah ke dalam 96 sumuran microtitre plate dan ditambah 100 ¼L L – DOPA (3 – 4 Dihydroxyphenil L–alanine, 1,6 mg/ml pada L–15). Sesudah 10 min OD diukur pada panjang gelombang 490 nm menggunakan microplate reader. Kandungan protein kemudian diukur dengan metode reaksi Folin (Lowry et al., 1985). Satu unit aktifitas enzim adalah penambahan absorbance sebanyak 0,001/min/ng protein. Hasil dan Pembahasan Udang windu yang diinfeksi secara buatan tidak menunjukkan adanya bintik putih seperti pada udang yang terinfeksi secara alami. Pada infeksi buatan dengan dosis infeksi virus yang tinggi deposit kalsium tidak sempat terbentuk pada cuticular epidermis karena udang terlalu cepat mati. Virion diketemukan pada pembengkakan inti sel dan di Kandungan Protein (mg/mL) Jenis sampel 0,25 0,2 0,15 0,1 0,05 0 W1 W2 W3 W4 Dosi s W S BV Gambar 1. Jumlah protein dari sediaan virus yang digunakan untuk infeksi WSBV pada udang windu. Jumlah dosis tersebut adalah W1 (0,234), W2 (0,117), W3 (0,029) dan W4 (0,058). Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 111 Aquacultura Indonesiana, Vol. 5, No. 3, Desember 2004 : 109–114 tersebut sampai bulan ke lima jumlah kematian kumulatif adalah 67% dan SR adalah 33% (Tabel 2). Jumlah jantan yang mati jauh lebih banyak dibandingkan dengan betina mencapai 35 ekor, pada umumnya kematian terjadi pada saat molting. Tabel 2. Udang windu P. monodon yang hidup (SR) sesudah diinfeksi dengan WSBV, keseluruhan udang yang diifeksi adalah 50 pasang udang windu Dosis yang digunakan 0,029 ng/mL Kontrol Kelulushidupan (%) 37 100 Lama kematian (hari) 150 0 Udang yang telah diinfeksi dengan WSBV terdeteksi virusnya dengan terekspresinya pita pada hasil PCR,Infeksi dilakukan dengan cara penyuntikan intramuscular dengan dosis 0,029 mg per ekor udang (Gambar 2). Proses metabolisme kalsium tersebut jika terganggu akan menyebabkan timbulnya bintik putih pada karapaks udang bagian dalam. Sedangkan kontrol dengan actin–R3 sebagai kontrol juga muncul dengan band tersendiri. Aktin sangat berperan pada proses metabolisme kalsium untuk pembentukan karapaks udang sehingga udang yang terinfeksi aktin tersebut tidak terekspresikan pada band kontrol. Udang yang hidup sekarang ini (F1) sudah terkontaminasi dengan WSBV, dan kualitas air serta faktor pertumbuhan dijaga seoptimal mungkin agar udang bisa hidup terus. Udang yang hidup sesudah terinfeksi oleh WSBV kemudian diisolasi hemocyte untuk dilakukan beberapa uji resistensi terhadap virus. Semestinya jika resisten respiratory burst activity akan meningkat karena jumlah atau produksi beberapa killing subctance akan meningkat. WSBV Gambar 2. Deteksi WSBV dengan PCR dari P. monodon Adanya reactive oxygen species dapat diuji dengan uji NBT telah lama dipakai oleh beberapa peneliti untuk mengukur immunostimulasi pada udang. Sung et al. (1996) mencatat bahwa produksi reactive oxygen (O2 ) yang mendapat perlakuan dengan pencelupan Vibrio bacterin dan glucan menjadi meningkat. Tetapi turun ke keadaan semula setelah 12 jam. Perbedaan konsentrasi immunostimulan akan mempengaruhi respon imun dari udang dan yang tertinggi mencapai 72 jam sesudah perlakuan. Kesimpulan sederhana adalah stimulasi atau infeksi udang windu dengan WSBV dapat meningkatkan produksi reactive oxygen species dari hemocyte (Tabel 3). Hasil diatas terlihat konstan selama hampir 1 bulan, untuk sementara hanya akan diperiksa sampai 1 bulan, karena keterbatasan jumlah sampel udang. Tabel 3. Produksi reactive oxygen species dari hemocyte udang sesudah infeksi dengan WSBV dan kontrol (tanpa perlakuan) Perlakuan OD630 sesudah perlakuan 7d Infeksi WSBV 2.1 Kontrol 0.1 14 d 21 d 28 d 2.3 2.2 2.1 0.1 0.2 0.1 Pro–phenol oxidase (proPO) merupakan sistem yang penting di dalam sistem pertahanan diri krustasea (Soderhall and Cerenius, 1992). Aktifasi proPO system (diukur dengan aktifitas prophenol oksidase) telah banyak dipakai oleh beberapa peneliti untuk mengukur sistem imun dari udang (Sung et al., 1994; 1996). Infeksi dengan WSBV akan membuat produksi proPO menjadi lebih tinggi jika dibandingkan dengan kontrol yang tanpa stimulasi apapun. Pola produksi proPO mirip dengan reactive oxygen species di mana sampai minggu keempat masih agak tinggi dibandingkan dengan kontrol udang. Tingginya proPO kemungkinan karena adanya infeksi WSBV dalam tubuh udang windu, tetapi alasan ini masih harus dibuktikan lebih lanjut, tetapi yang terpenting adalah bahwa udang yang diinfeksi dengan WSBV masih bisa hidup sampai saat ini. Pada Tabel 4 menunjukkan produksi proPO dari udang windu yang terinfeksi oleh WSBV. Infeksi dengan WSBV membuat produksi proPO tinggi selama hampir 1 bulan sesudah infeksi pertama. Apakah ini disebabkan oleh adanya WSBV pada tubuh udang Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 112 Studi pendahuluan resistensi udang windu Penaeus monodon (Hari Suprapto dan M. Murjani) windu, tentunya masih harus diketahui dengan pasti, misalnya –glucan mempunyai peran penting didalam pengaktifan proPO system pada arthropoda (Soderhall, 1982; 1992; 1990; 1998) –glucan binding protein (BGBP) melakukan penguatan untuk aktifasi. Tabel 4. Produksi proPO dari hemocyte udang windu sesudah infeksi dengan WSBV dan kontrol (tanpa perlakuan) Perlakuan Phenol oxidase activity (/min/ng protein) 7d Infeksi WSBV Kontrol 31.10 3.00 14 d 21 d 28 d 33.05 32.22 32.20 4.10 3.20 4.10 Pada udang yang sama seperti pada krustase lain, melanisasi disebabkan oleh reaksi prophenoloxidase (ProPO) yang menyebabkan hydroksilasi phenol dan oksidasi dari o–phenol mejadi quinons, sebagai respon dari invasi material asing ke dalam tubuh udang selama proses penyembuhan (Soderhall et al., 1994). Quinons ini kemudian ditranformasikan menjadi melanin oleh reaksi non enzymatic. Walaupun aktifitas mikroba terhadap melanin secara langsung telah banyak dibahas, tetapi produksi recative oxygen species misalnya superoxide anginons dan gugus hydroxyl radicals selama proses quinons generasi juga amat penting sebagai antimikrobial (Song and Hsieh, 1994). ProPO pada krustase terletak pada granule hemocytic sebagai pro–enzyme inaktif yang biasa disebut dengan proPO yang diketahui ikut di dalam serangkaian reaksi yang dikenal dengan proPO activating system. Sistem ini secara khusus dapat diaktifkan dengan –1, 3–glucan (Vargas Albores, 1995), sehingga proPO krustase diakui sebagai sistem pertahanan diri. Pada udang ada 2 macam protein yang terlibat untuk mengenali produk mikroba dan seluler aktifasinya telah dijelaskan (Vargas Albores, 1995). Grup yang pertama adalah multivalanet sugar binding agglutinins juga dinamakan agglutininins atau lektin. Grup kedua terdiri dari melekul monovalen dan tidak mengakibatkan agglutinasi walaupun mampu mengikat residu gula. Aktifitas agglutinasi telah dideteksi pada plasma P. monodon (Ratapao and Chulavatnatol, 1990). P. monodon plasma adalah sebuah molekul glycoprotein dengan berat 420 kDa dan lektin ini dinamakan monodin dapat beraksi dengan NANA (N – acethyl neuraminic acid) dan N–acethyl amino sugars (GalNAc, GlcNAc dan ManNAc) yang diketahui dengan studi inhibisi. Monodin diketahui dapat menyebabkan aglutinasi pada Vibrio vulnificus, bakteri yang sangat merugikan budidaya udang. Walaupun secara khusus belum diketahui secara pasti, tetapi agglutinins ini dapat bereaksi dengan LPS dari bakteri dan dapat menyebabkan agglutinasi dari beberapa Vibrio, termasuk V. parahaemolyticus. Selanjutnya reaksi tersebut juga bisa dihambat dengan LPS (Vargas Albores, 1993). LPS ” binding agglutinins (LPS – BA) kelihatannya adalah melekul dengan berat 180–170 kDa yang tersusun dari 41 kDa sebanyak 4 unit. Protein yang digunakan untuk mengenali produk mikroba terdapat pada plasma udang dan mempunyai kemampuan beraksi dengan beta glucan sehingga dinamakan beta glucan binding protein (BGBP). BGBP dari krustase yang pertama kali dilaporkan dari crayfish Pasifastacus leniusculus (Duvic dan Soderhall, 1990). Antibodi terhadap crayfish BGBP telah dipakai untuk mendeteksi protein ini pada crustacea lain termasuk diantaranya adalah udang (Duvic dan Soderhall, 1993). BGBP telah dimurnikan dari Penaeus californiensis, Penaeus vannamei dan Penaeus stylirostris plasma dengan berat molekul 100–kDa protein monomeric. Daftar Pustaka Chen, D. 1992. An overview of the disease situation, diagnostic techniques, treatments and preventives used on shrimp farms in China. Diseases of Cultured Penaeid Shrimp in Asia and United States, edited by The Oceanic Institute, Hawaii, pp. 47–55. Chou, H.Y., C.Y. Huang, C.H. Wang, H.C. Chiang and C.F. Lou. 1995. Pathogenicity of baculovirus infection causing white spot syndrome in the cultured penaeid shrimp in Taiwan. Diseases of Aquatic and Organisms, 23: 165–173. Duvic, B. and Soderhall. 1990. Purification and characterization of a –1, 3–glucan –binding protein in crustacean blood. Journal of Biological and Chemical, 269: 29462–29467. Duvic, B. and Soderhall. 1993. –1, 3–glucan–binding protein from plasma of the freshwater crayfish Astacus astacus and Procambarus clarkii. Journal Crustasean Biology, 13: 403–408. Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 113 Studi pendahuluan resistensi udang windu Penaeus monodon (Hari Suprapto dan M. Murjani) Flegel, T., D.F. Fegan, S. Kongsom, S. Vutikomudomkit, S. Sriurairatana, S. Boonyaratpalin, C. Chantanacookhin, J.E. Vickers and O.D. McDonalds. 1992. Occurrence, Diagnosis and Treatment of Shrimp Disease in Thailand. The Oceanic Institut, Honolulu Hawaii, pp. 57–112. Lightner, D.V. 1994. A handbook of shrimp pathology and diagnostic procedure for disease of cultured penaeid shrimp. Journal of The World Aquaculture Society, pp. 12–16 Lowry, D.B., P.S. Jackett and P.W. Andrew. 1985. Activation of macrophage for antimicrobial activity. Immunology Letter, 11: 195–203. Ratapao, S. and M. Chulavatnatol. 1990 Monodin, a new sialic acid specific lectin from black tiger prawn Penaeus monodon. Comparative Biochemistry Physiology, 97B: 515–520. Sambrook, J., E.F. Fritsch and T. Maniatis. 1989 Molecular cloning, a laboratory manual 2nd. Cold Spring Harbour Laboratory Press., Harbour, New York, 66 pp. Song, Y.L. and Y.T. Hsieh. 1994. Immunostimulan of tiger shrimp Penaeus monodon hemocyte for generation of microbial substances. Analysis of Reactive Oxygen Species Develop. and Comparative Immunology, 18: 201–209. Sung, H.H., G.H. Kou and Y.L. Song. 1994. Vibriosis resistance induce by glucan treatment in tiger shrimp P. monodon. Fish Pathology, 29: 11–17. Sung, H.H., Y.L. Yans and Y.L.J. Song. 1996. Enhancement of microbial activity in the tiger shrimp P. monodon via immunostimulation. Journal Crustacean Biology, 16: 278–284. Soderhall, K. and L. Cerenius. 1992. Crustacean Immunity. Ann. Rev. Fish. Dis., 2: 3–23. Soderhall, K. 1982. Prophenoloxidase activating system and melanization–a recognition system of anthropods. A review. Development Comparative Immunology, 6: 601–611. Soderhall, K. 1992. Biochemical and molecular aspects of cellular communication in arthropods. Biology Zoology, 59: 141–151. Soderhall, K., W. Rogener, I. Soderhall, R.P. Newton and N.A. Ratcliffe. 1998. The properties and purification of a Blaberus craniifer plasma protein which enhances the activation of hemocyte prophenoloxidase by –1–3–glucan. Insect. Biochem., 18: 323–330. Soderhall, K., L. Cerrenius and M.W. Johansen. 1994. The prophenoloxidase activating system and its role in intervertebrate defense. Ann. NY Acad. Sci., 712: 155–161. Suprapto, H. 2003. Electron microscope evidence of bacilliform virus the causative agent of White spot disease in tiger shrimp Penaeus monodon in natural infection. Jurnal Kelautan Undip. 3(2) : 6–8. Takahashi, Y., T. Itami, M. Kondo, M. Maeda, R. Fujii, S. Tomonaga, K. Supamattaya and S. Bonnyaratpalin. 1994. Electrone microscopic evidence of bacilliform virus infection in Kuruma shrimp Penaeus japonicus. Fish Pathology, 29: 121–125. Vargas Albores, F. 1995. The defense system of brown shrimp Penaeus californiensis: Humoral recognition cellular responses. Journal marine Biotechnology, 3: 153–156. Vargas Albores, F., A. Guzman Murillo and J.L. Ochoa. 1993. A lipopolysaccharide bidning protein agglutinin isolated from brown shrimp Penaeus californiensis HOLMES ha emolymph. Comparative Biochemystry Physiology, 194A: 407–413. Hak cipta oleh Masyarakat Akuakultur Indonesia 2005 114