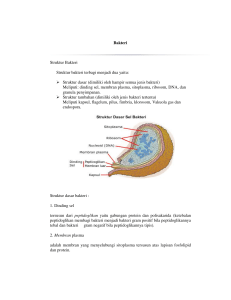
Karakteristik pertumbuhan bakteri asam laktat
advertisement

HASIL DAN PEMBAHASAN Bakteri asam laktat yang digunakan merupakan hasil isolasi dari susu sapi segar dan produk olahannya. Bakteri asam laktat indigenous susu sapi segar dan produk olahannya ini berpotensi untuk menjadi kandidat bakteri probiotik, dengan syarat tahan pada berbagai kondisi dalam saluran pencernaan. Konfirmasi Kultur Starter Indigenous Susu Sapi Segar dan Bakteri Patogen Uji Bakteri asam laktat dan bakteri patogen yang diujikan harus terbebas dari kontaminasi bakteri lain. Uji konfirmasi dilakukan dengan pewarnaan Gram dan uji katalase untuk melihat morfologi BAL dan bakteri patogen uji yang seragam, homogen dan murni. Pewarnaan Gram dilakukan untuk menggolongkan jenis bakteri Gram positif atau negatif. Uji katalase dilakukan untuk mengetahui BAL dan bakteri patogen uji termasuk sifat katalase negatif atau positif. Karakteristik BAL dan bakteri patogen uji melalui uji konfirmasi dapat dilihat pada Tabel 2. Tabel 2. Karakteristik BAL Indigenous Susu Sapi Segar dan Bakteri Patogen Uji Bakteri Pewarnaan Gram Positif Morfologi Bentuk dan Susunan Bulat rantai Sifat Katalase Negatif L. bulgaricus S-01* Positif Batang panjang Negatif B. longum Y-01 * Positif Batang pendek Negatif L. acidophilus Y-01 * Positif Batang Negatif S. aureus ATCC 25923** Positif Bulat Positif E. coli ATCC 25922** Negatif Batang Positif S. Typhimurium ATCC 14028 Negatif Batang Positif S. thermophilus S-01* Keterangan : * sesuai dengan Wardani (2009) ** sesuai dengan Ambarwati dan Gama (2009) Pewarnaan Gram dilakukan untuk mengetahui bakteri tersebut termasuk Gram negatif atau Gram positif. Perbedaan ini dilihat berdasarkan komposisi dinding sel dan sifat pewarnaannya. Selain itu, juga berbeda dalam sifat sensitifitasnya terhadap kerusakan mekanis atau fisis, terhadap enzim, desinfektan dan antibiotik (Hadioetomo, 1993). S. thermophilus S-01, L. bulgaricus S-01, B. longum Y-01, L. acidophilus Y-01 dan S. aureus ATCC 25923 merupakan bakteri Gram 23 positif, sedangkan E. coli ATCC 25922 dan S. Typhimurium ATCC 14028 adalah bakteri Gram negatif. Hasil pewarnaan menunjukkan bakteri-bakteri yang tergolong Gram positif tersebut terlihat berwarna biru (menyerap pewarna kristal violet), sedangkan bakteri-bakteri Gram negatif terlihat berwarna merah, karena lebih menyerap pewarna safranin. Menurut Hadioetomo (1993), bakteri Gram negatif bersifat lebih konsisten terhadap reaksi pewarnaan, tetapi bakteri Gram positif sering berubah sifat pewarnaannya, sehingga bakteri Gram positif dapat menunjukkan reaksi Gram variable. Kultur bakteri yang biasa digunakan dalam pewarnaan Gram adalah kultur berumur 24 jam, jika kultur bakteri terlalu tua, maka bakteri tersebut akan kehilangan kemampuan untuk menyerap kristal violet dan lebih menyerap pewarna safranin. Perubahan ini dapat disebabkan juga oleh perubahan kondisi lingkungan dan modifikasi teknik pewarnaan. Pewarnaan awal bakteri adalah dengan zat warna basa kristal violet, jika kelebihan warna, maka dicuci dengan air, kemudian diberi larutan yodium yang akan membentuk suatu kompleks antara kristal violet dan iodium. Pencucian dengan alkohol dimaksudkan untuk mencuci kompleks tersebut, sehingga kompleks tersebut keluar dari dinding sel bakteri Gram negatif, sehingga saat diberi pewarna safranin, dinding sel Gram negatif akan menyerap safranin dan menjadi berwarna merah. Gram positif tidak ada kompleks yang keluar karena lapisan peptidoglikan pada dinding selnya lebih tebal dan menyebabkan Gram positif tidak menyerap warna safranin dan tetap berwarna biru seperti Kristal violet (Hadioetomo, 1993). Mekanisme pewarnaan Gram memang didasarkan pada struktur dan komposisi dinding sel bakteri. Bakteri Gram negatif mengandung lipid, lemak atau substansi seperti lemak dalam persentase tinggi daripada yang dikandung dari Gram positif. Dinding bakteri Gram negatif juga lebih tipis. Alkohol dalam pewarnaan menyebabkan terekstraksinya lipid sehingga memperbesar daya permeabilitas dinding sel Gram negatif. Pewarna Kristal violet dan iodium yang sudah masuk dalam dinding pun dapat diekstraksi. Gram positif dengan kandungan lemak lebih rendah, jika diberi alkohol justru membuat pori-pori mengecil dan permeabilitas berkurang, sehingga Kristal violet dan iodium tidak dapat terekstraksi (Pelczar dan Chan, 2007). 24 Pewarnaan Gram yang dilakukan terhadap keempat bakteri asam laktat menunjukkan hasil bentuk bakteri B. longum Y-01, L. acidophilus Y-01 dan L. bulgaricus S-01 adalah batang, sedangkan S. thermophilus S-01 berbentuk bulat seperti rantai. Seperti yang dikemukakan Tamime dan Robinson (2008), S.thermophilus memang berbentuk batang yang membentuk susunan seperti rantai dan L. bulgaricus berbentuk batang panjang. Bentuk BAL hasil ewarnaan Gram dapat dilihat pada Gambar 3. a). Streptococcus thermophilus b). Lactobacillus bulgaricus c). Bifidobacterium longum d). Lactobacillus acidophilus Gambar 3. Bentuk Bakteri Asam Laktat Indigenous Susu Sapi Segar Hasil Pewarnaan Gram Pewarnaan Gram bakteri patogen pada Gambar 4, menunjukkan bentuk masing-masing bakteri patogen setelah dilakukan pewarnaan Gram dan sesuai dengan Hadioetomo (1993). E. coli berbentuk batang lurus. S. Typhimurium berbentuk batang, sedangkan S. aureus berbentuk kokus membentuk anggur. Uji katalase digunakan untuk mengetahui aktivitas katalase pada bakteri yang diuji. Kebanyakan bakteri memproduksi enzim katalase yang dapat memecah H2O2 menjadi H2O dan O2. Enzim katalase diduga penting untuk pertumbuhan aerobik 25 karena H2O2 bersifat racun terhadap sel mikroba. Beberapa bakteri yang termasuk katalase negatif adalah Streptococcus, Leuconostoc, Lactobacillus dan Clostridium. Escherichia coli Salmonella Typhimurium Stapylococcus aureus Gambar 4. Bentuk Bakteri Patogen Hasil Pewarnaan Gram Bakteri katalase positif seperti S. aureus bisa menghasilkan gelembunggelembung oksigen karena adanya pemecahan hidrogen peroksida (H2O2) oleh enzim katalase yang dihasilkan oleh bakteri itu sendiri. Komponen H2O2 ini merupakan salah satu hasil respirasi aerobik bakteri, misalnya S. aureus, dimana hasil respirasi tersebut justru dapat menghambat pertumbuhan bakteri karena bersifat toksik bagi bakteri itu sendiri. Oleh karena itu, komponen ini harus dipecah agar tidak bersifat toksik lagi. Bakteri katalase negatif tidak menghasilkan gelembung-gelembung. Hal ini berarti H2O2 yang diberikan tidak dipecah oleh bakteri katalase negatif, sehingga tidak menghasilkan oksigen. Bakteri katalase negatif tidak memiliki enzim katalase yang menguraikan H2O2. Ketahanan Kultur Starter BAL Indigenous Susu Sapi Segar pada pH Berbeda Daya tahan hidup bakteri asam laktat dalam saluran pencernaan merupakan syarat mikroorganisme untuk dapat memberikan manfaat kesehatan setelah dikonsumsi. Stress yang pertama terjadi pada sel bakteri yang ada dalam saluran pencernaan adalah pada kondisi asam lambung. Chou dan Weimer (1999), waktu yang diperlukan mulai dari bakteri masuk sampai keluar lambung sekitar 90 menit. Jadi bakteri asam laktat yang diseleksi untuk digunakan sebagai probiotik harus mampu bertahan dalam keadaan asam lambung (pH 2 dan 2,5) selama sedikitnya 90 menit. Ketahanan BAL pada pH berbeda pada penelitian ini dilakukan selama 180 menit atau 2 kali lebih lama dari waktu normal makanan masuk lambung sampai keluar lambung, untuk memastikan ketahanan BAL pada kondisi pH yang berbeda. 26 Pengaturan pH media pertumbuhan BAL untuk menyamakan dengan kondisi dalam lambung (2, 2,5 dan 3,2) dan usus halus (7,2) adalah dengan menambahkan HCl dan NaOH. HCl digunakan untuk menurunkan pH media hingga mencapai kondisi pH yang diinginkan. Alasan penggunaan HCl adalah karena HCl mempunyai karakteristik yang hampir sama dengan asam yang dihasilkan di lambung. HCl adalah asam kuat yang mudah terdisosiasi menghasilkan proton, yang menyebabkan penurunan pH medium di luar sel atau pH ekstraseluler. NaOH digunakan untuk menaikkan pH, hingga mencapai kondisi pH 7,2, karena merupakan basa kuat, mengandung Na (garam) yang sama dengan karakteristik garam empedu yang disekresikan di dalam duodenum. Jumlah populasi BAL sebelum dan sesudah inkubasi dapat dilihat pada Tabel 3. Tabel 3. Jumlah Populasi BAL Indigenous Susu Sapi Segar pada pH Berbeda No Lama . inkubasi 1. P0 menit P180 menit (P180- P0) 2. P0 menit P180 menit (P180- P0) 3. P0 menit P180 menit (P180- P0) 4. P0 menit P180 menit (P180- P0) S. thermophilus S-01 Populasi BAL (log10 cfu/ml) L bulgaricus B. longum S-01 Y-01 L. acidophilus Y-01 ---------------------------pH 2,0 ± 0,1------------------------------7,83 ± 0,44 7,15 ± 0,30 7,06 ± 0,12 7,60 ± 0,22 ND 7,31 ± 0,3 7,15 ± 0,13 7,42 ± 0,31 -7,83 ±0,44 0,16 ± 0,11 0,09 ± 0,06 -0,18±0,13 --------------------------pH 2,5 ± 0,1 ----------------------------6,89 ± 0,87 7,69 ± 0,13 7,36 ± 0,09 b 7,25 ± 0,02 a ND 6,50 ± 0,73 7,62 ± 0,06 7,52 ± 0,12 -6,89 ± 0,87 -1,19 ± 0,84 0,26 ± 0,18 0,29 ± 0,21 -----------------------------pH 3,2 ± 0,1 ----------------------------------8,12 a ± 0,71 7,87 ± 0,19 7,50 ± 0,33 7,12 b ± 0,03 6,09 b ± 0,71 7,43 ± 0,52 7,92 ± 0,29 7,43 a ± 0,08 -2,03 ± 1,43 -0,45 ± 0,31 0,42 ± 0,30 0,31 ± 0,22 --------------------------- --pH 7,2 ± 0,1----------------------------------6,90 ± 0,09 8,12 ± 0,14 7,01 b ± 0,17 7,45 b ± 0,08 7,32 ± 0,30 7,28 ± 0,40 7,45 a ±0,15 7,81 a ± 0,14 0,42 ± 0,30 -0,84 ± 0,60 0,44 ± 0,31 0,36 ± 0,25 Keterangan : * apabila hasilnya (-) menunjukan adanya kematian; ** huruf superskript (a dan b) yang berbeda pada kolom yang sama menunjukkan berbeda nyata (P<0,05) ND = not detected (tidak terdeteksi adanya pertumbuhan) Ketahanan bakteri asam laktat indigenous susu sapi segar dan produk olahannya yang akan dijadikan sebagai bakteri probiotik harus tahan pada kondisi 27 asam dalam saluran pencernaan, salah satunya adalah kondisi di lambung yang dalam keadaan kosong mempunyai pH sangat asam, sekitar 1–2,5. Hasil sekresi lambung yang dikenal dengan getah lambung merupakan cairan jernih berwarna kuning pucat yang mengandung HCL 0,2–0,5% dan asam lambung dengan pH 3,2 disekresikan di dalam lambung (Mitsuoka, 1990). Menurut Jacobsen et al. (1999), dalam saluran pencernaan manusia, pH asam lambung yaitu sekitar 2,5, sedangkan pengujian ketahanan BAL pada pH 7,2 diujikan karena garam empedu disekresikan di dalam duodenum yang mempunyai kondisi pH netral sekitar 6,9-7,2. Hasil pada tabel 3, menunjukkan S. thermophilus S-01 pada pH 2 dan 2,5 tidak dapat bertahan, ini dibuktikan dengan tidak adanya pertumbuhan yang terdeteksi setelah akhir perlakuan. S. thermophilus pada pH 3,2 sudah mulai bertahan, walaupun masih mengalami penurunan populasi sekitar 2,03 log10 cfu/ml, sedangkan pada pH 7,2 S. thermophilus dapat bertahan, bahkan mengalami kenaikan populasi sekitar 0,42 log10 cfu/ml. Hasil analisis ragam menunjukkan S.thermophilus S-01 pada kondisi pH 3,2 sebelum dan sesudah inkubasi berpengaruh nyata (P<0,05). Walaupun S. thermophilus S-01 mengalami penurunan, namun populasi akhir setelah diinkubasi 3 jam masih pada populasi 6,09 log10 cfu/ml. Jika dibandingkan dengan kondisi pH 2 dan 2,5, maka S. thermophilus S-01 lebih tahan pada kondisi 3,2 dan 7,2. Pada kondisi pH 3,2, S. thermophilus S-01 mampu mempertahankan pH intraseluler sitoplasma, sehingga pH sitoplasma lebih basa dibanding dengan pH ekstraseluler. Membran sel S. thermophilus S-01 juga lebih tahan terhadap kebocoran pada pH 3,2. Ketiga BAL yang lain mampu bertahan dalam semua kondisi pH, terutama B. longum Y-01 dan L. acidophilus Y-01, bahkan mengalami kenaikan populasi. L. bulgaricus S-01 juga mampu bertahan dalam semua kondisi pH, walaupun mengalami penurunan populasi, mulai dari 0,18–0,8 log10 cfu/ml. Hasil analisis ragam menunjukkan bahwa kondisi pH 7,2 berpengaruh nyata pada populasi awal dan akhir setelah inkubasi 3 jam bakteri asam laktat B. longum Y-01 dan L. acidophilus Y-01. Tiap galur mempunyai ketahanan yang berbeda terhadap asam atau pH rendah, contohnya Lactobacillus lebih toleran terhadap pH rendah dibandingkan Lactococci dan Streptococci (Farida, 2006). Bifidobacteria juga dapat bertahan dalam keadaan pH rendah, yaitu pH 2 (Zinedine dan Faid, 2007). Ketahanan yang 28 tinggi dari Bifidobacteria sangat penting, karena mikroorganisme ini digunakan dalam formulasi atau ditambahkan dalam makanan dan harus tahan terhadap keasaman, terutama selama transit dalam lambung. Toleransi terhadap asam pada bakteri asam laktat tergantung pada pH H+ATPase dan komposisi membran sitoplasma, yang sangat dipengaruhi oleh jenis bakteri, media pertumbuhan dan kondisi inkubasi (Oh et al., 2000). Saat memasuki saluran pencernaan dengan kondisi pH sangat asam, bakteri dapat mengalami stress. Stress yang dialami adalah karena pH intraseluler bakteri berbeda dengan pH ekstraseluler, sehingga bakteri asam laktat harus melakukan metabolisme agar dapat menyesuaikan kondisi intraseluler dengan ekstraselulernya. Menurut Siegumfeldt et al. (2000), pada bakteri asam laktat terjadi perubahan dinamis pH intraseluler seiring dengan terjadinya penurunan pH ekstraseluler sehingga tidak terjadi gradien proton yang besar. Toleransi bakteri asam laktat yang cukup tinggi terhadap asam disebabkan oleh kemampuannya untuk mempertahankan pH sitoplasma lebih basa daripada pH ekstraseluler. Bila sel bakteri terpapar pada kondisi yang sangat asam, maka membran sel dapat mengalami kerusakan dan berakibat hilangnya komponen intraseluler. Biasanya kerusakan ini menyebabkan kematian pada sel. Bakteri yang toleran terhadap asam, membran selnya lebih tahan terhadap kebocoran akibat pH rendah dibandingkan dengan yang tidak tahan. Secara in vitro ketahanan terhadap pH rendah tergantung pada tipe buffer dan tipe makanan yang digunakan. Menurut Hardiningsih et al. (2005), ketahanan fisiologis dari setiap bakteri berbeda-beda. Perbedaan ketahanan membran sel bakteri terhadap kerusakan akibat terjadinya penurunan pH ekstraseluler menyebabkan keragaman ketahanan sel pada pH rendah. Jacobsen et al. (1999) melakukan pengujian ketahanan 47 isolat bakteri asam laktat dari berbagai sumber pada pH 2,5. Sebanyak 47 isolat tersebut hanya 29 isolat yang mampu bertahan pada pH 2,5 dan tidak ada satupun yang mampu tumbuh setelah inkubasi empat jam. Kondisi pH 2,5 bagi beberapa bakteri asam laktat memang sangat asam, sehingga banyak yang tidak dapat mempertahankan kondisi basa intraseluler dan terjadi kerusakan pada membran sel bakteri. Berdasarkan hasil ini, S. thermophilus S-01 dan L. bulgaricus S-01 diduga mengalami kerusakan membran sel. 29 BAL mempunyai dinding sel yang tebal dengan lipid yang tipis sekitar 1-4%. Menurut (Fardiaz, 1989), bakteri Gram positif, 90% dari dinding selnya terdiri dari lapisan peptidoglikan, sedangkan lapisan tipis lainnya adalah asam teikoat. Asam teikoat mengandung unit-unit gliserol atau ribitol yang terikat satu sama lain oleh ester fosfat dan biasanya mengandung gula lain dan D-alanin. Peptidoglikan ini terdiri dari turunan gula yaitu N-asetilglukosamin dan N-asetilmuramat serta beberapa asam amino yaitu L-alanin, D-alanin, asam D-glutamat dan asam diaminopimelat (ADP). Peptidoglikan yang tebal dan ikatan rantai asam teikoat inilah yang dapat mempertahankan bentuk dinding sel dari kondisi asam ekstraseluler. Kondisi ektraseluler yang asam memang dapat menyebabkan dinding sel BAL menjadi lisis. Namun, dinding sel BAL ini dapat mempertahankan bentuknya, sehingga bagian dalam sel masih dapat terlindungi. Membran sel pun dapat mengangkut nutrien dan garam yang dibutuhkan untuk melakukan metabolisme dan tidak permeabel terhadap senyawa yang tidak dibutuhkan. Lapisan lipid yang tipis membuat pori-pori dinding menjadi mengecil, sehingga permeabilitas sel berkurang dan tidak terjadi ekstraksi komponen intraseluler. Asam dapat merusak lapisan lipid yang ada pada membran sel, namun lapisan lipid ini mengandung protein khusus. Beberapa dari protein membran merupakan enzim, sementara yang lainnya dapat mengikat nutrien dan mengangkutnya ke dalam sel. Menurut Chou dan Weimer (1999), strain L. acidophilus pada pH sekitar 3,5 hanya tahan jangka pendek, terlihat dari pertumbuhannya yang sedikit melambat setelah inkubasi 90 menit. Tidak seperti bakteri netrofilik (yang dapat tumbuh pada kondisi pH mendekati netral) yang menjaga pH intraselulernya mendekati netral, pada bakteri asam laktat terjadi perubahan dinamis pH intraseluler seiring dengan perubahan pH lingkungannya (Siegumfeldt et al., 2000). Ini membuktikan, semua bakteri yang diisolasi memang mempunyai karakteristik yang berbeda dan metabolisme yang berbeda, bakteri-bakteri ini bersifat strain dependent. Pertumbuhan optimal L. bulgaricus adalah pada kondisi asam, pH sekitar 5,2–5,8 dan akan terhenti pada pH sekitar 3,8. S. thermophilus S-01 tumbuh optimal pada pH sekitar 6,5 dan akan terhenti pada pH 4,2–4,4. Pada pembuatan yogurt digunakan campuran S. thermophilus dan L. bulgaricus. S. thermophilus akan tumbuh lebih cepat pada awal masa inkubasi. S. thermophilus menghasilkan asam 30 folat, asam laktat piruvat dan asam format untuk menstimulir pertumbuhan L.bulgaricus, sehingga proses fermentasi lebih cepat. Pada akhir proses fermentasi, pertumbuhan S. thermophilus mulai berkurang seiring dengan meningkatnya L.bulgaricus (Surono, 2004; Silva et al., 2005). Oh et al. (2000) menyatakan bahwa dalam proses fermentasi yogurt, terbukti L. bulgaricus lebih optimal tumbuh pada kondisi asam, sedangkan S. thermophilus lebih optimal tumbuh pada pH mendekati netral (6,5). B. longum dan L. acidophilus adalah bakteri yang sering digunakan sebagai bakteri probiotik dalam produk susu, karena pada dasarnya kedua spesies ini dominan dalam saluran pencernaan manusia. Bahkan, kedua spesies ini terdapat secara alami dalam usus manusia. Perubahan populasi BAL indigenous susu sapi segar selama pengamatan 180 menit pada kondisi pH berbeda dapat dilihat pada Gambar 6. S. thermophilus S01 pada kondisi 2, 2,5 dan 3,2 mengalami penurunan populasi setiap pengamatan, terutama setelah 60 menit inkubasi, terjadi penurunan populasi hingga akhir perlakuan. Sedangkan pada kondisi pH 7,2, terlihat grafik yang belum stabil pada 60 menit pertama, namun setelahnya grafik terlihat statis, yang menunjukkan ketahanan bakteri yang baik. L. bulgaricus S-01 pada kondisi pH rendah 2 dan 2,5 menunjukkan pergerakan yang tidak stabil, sedangkan pada kondisi pH 3,2 dan 7,2, grafik terlihat statis, yang menunjukkan L. bulgaricus S-01 lebih mampu bertahan pada pH 3,2 dan 7,2. Perubahan populasi S. thermophilus S-01 pada pH 3,2 lebih tahan dibandingkan pada pH 2, 2,5 dan 7,2. Walaupun mengalami penurunan populasi, namun S. thermophilus S-01 masih dapat bertahan pada kisaran populasi 7-8 log10 cfu/ml. Sedangkan L. bulgaricus S-01 relatif stabil, bahkan mengalami peningkatan populasi pada 60 menit pertama pada semua kondisi pH dan mulai mengalami penurunan populasi setelah 90 menit. BAL yang diseleksi untuk menjadi bakteri probiotik, setidaknya harus mampu bertahan pada kondisi pH rendah sekitar 90 menit. B. longum Y-01 pada kondisi pH 2, menunjukkan grafik yang statis, sedangkan pada pH 2,5, 3,2 dan 7,2 terlihat adanya kenaikan populasi setelah ±2 jam inkubasi. Perubahan populasi BAL selama inkubasi 180 menit dapat dilihat pada Gambar 5. 31 Keterangan : pH 2.0 ( ), pH 2.5 ( ), pH 3.2 ( ), pH 7.2 ( ) Gambar 5. Perubahan Populasi Kultur Starter BAL a). S. thermophilus S-01, b). L.bulgaricus S-01, c). B. longum Y-01, d). L. acidophilus Y-01 pada pH berbeda 32 Grafik L. acidophilus Y-01 pada pH 2, 2,5 dan 3,2 pada 90 menit pertama terlihat masih labil, namun setelah inkubasi 90 menit, grafik menunjukkan kenaikan populasi, sedangkan pada kondisi pH 7,2 L. acidophilus Y-01 lebih mampu bertahan sehingga grafik menunjukkan kenaikan yang konsisten. Waktu normal yang diperlukan makanan mulai dari masuk hingga keluar lambung adalah sekitar 90 menit, sedangkan waktu yang digunakan dalam penelitian ini adalah 180 menit, atau 2 kali dari waktu minimal yang diperlukan untuk menyeleksi BAL menjadi bakteri probiotik. Tujuannya adalah untuk memastikan BAL benar-benar mampu bertahan pada pH 2, 2,5, 3,2 dan 7,2. Ketahanan Kultur Starter BAL Indigenous Susu Sapi Segar pada Garam Empedu Bakteri asam laktat sebagai kultur probiotik harus mampu melewati kondisi saluran pencernaan yang menekan, agar dapat bertahan dan tumbuh. Salah satunya adalah pada saat bakteri memasuki bagian atau usus di mana empedu disekresikan di dalam usus. Populasi BAL indigenous susu sapi segar pada garam empedu sebelum dan sesudah inkubasi selama 24 jam disajikan pada Tabel 4. Tabel 4. Jumlah Populasi BAL Indigenous Susu Sapi Segar pada Garam Empedu 1. S. thermophilus S-01 Populasi BAL (Log CFU/ml) P0 jam P24 jam (P24 - P0)* 7,91 ± 0,19 ND -7,91 ± 0,19 2. L. bulgaricus S-01 8,08 a ± 0,10 5,84 b ± 0,054 -2,25 ± 1,50 3. B. longum Y-01 8,41 b ± 0,03 8,51 a ± 0,01 0,10 ± 0,07 4. L. acidophilus Y-01 7,54 b ± 0,01 7,78 a ± 0,07 0,24 ± 0,17 No Kultur bakteri Keterangan : * apabila hasilnya (-) menunjukan adanya kematian; ** huruf superskript (a dan b) yang berbeda pada baris yang sama menunjukkan berbeda nyata (P<0,05) ND = not detected (tidak terdeteksi adanya pertumbuhan) Perlakuan secara in vitro, dimana media dikondisikan seperti di dalam saluran usus, yaitu dengan menambahkan oxgall sebanyak 0,3%. Penambahan 0,3% oxgall merupakan konsentrasi kritis dan cukup tinggi untuk menseleksi galur resisten. Peningkatan konsentrasi garam empedu dapat menyebabkan penurunan jumlah koloni bakteri asam laktat yang lebih besar (Farida, 2006). 33 Populasi sebelum perlakuan dan setelah perlakuan pada setiap bakteri asam laktat indigenous susu sapi segar sangat berbeda (P<0,01). Keragaman toleransi bakteri asam laktat terhadap garam empedu tidak berhubungan dengan perbedaan spesies, tetapi tergantung dari masing-masing galur atau bersifat strain dependent. Galur yang berbeda mempunyai mekanisme yang berbeda terhadap garam empedu. Galur bakteri asam laktat yang sama serta diisolasi dari sumber yang sama pun, mempunyai keragaman pada toleransi terhadap garam empedu (Chou dan Weimer, 1999). Menurut Sanders (2000), Lactobacillus dan Bifidobacterium secara umum lebih resisten terhadap garam empedu dibandingkan dengan genus Streptococcus dan genus lainnya, terutama bakteri asam laktat yang digunakan sebagai kultur starter yogurt. Toleransi terhadap garam empedu ini disebabkan oleh peranan polisakarida sebagai salah satu komponen penyusun dinding sel bakteri Gram positif. Toleransi terhadap garam empedu yang rendah pada Streptococcus dibuktikan dengan hasil populasi akhir S. thermophilus S-01 setelah mendapat perlakuan garam empedu, bakteri tidak tahan garam empedu sehingga pada saat akhir perlakuan, bakteri tersebut mati dan tidak terhitung populasinya. Farida (2006) menambahkan, sel yang mati disebabkan karena peningkatan aktivitas enzim β-galactosidase terhadap garam empedu, enzim β-galactosidase pada bakteri asam laktat digunakan untuk menghasilkan asam laktat dari laktosa (Surono, 2004). Peningkatan aktivitas enzim ini adalah upaya bakteri untuk melakukan adaptasi terhadap kondisi ekstraseluler, sehingga kecepatan difusi molekul nutrien pun mengalami peningkatan dan membatasi sel untuk mengontrol metabolismenya Peningkatan permeabilitas sel ini mengakibatkan materi intraseluler, seperti sitoplasma dan ribosom terekstraksi dan sel mengalami lisis. Padahal sitoplasma berfungsi sebagai tempat berlangsungnya hanpir semua reaksi enzimatis dari metabolisme sel. Dalam sitoplasma, sel menggunakan energi kimia untuk membangun dan mempertahankan strukturnya serta melakukan pergerakan sel. Ribosom berfungsi mensintesa protein. Jika materi intraseluler ini, maka bakteri tidak bisa melakukan metabolisme, sehingga tidak dapat tumbuh dan bahkan mati (Lehninger, 2004). Perubahan populasi BAL setiap 60 menit selama pengamatan 24 jam dapat dilihat pada Gambar 6. 34 Keterangan : kontrol ( ), garam empedu ( ) Gambar 6. Perubahan Populasi Kultur Starter a). S. thermophilus S-01, b). L.bulgaricus S-01, c). B. longum Y-01, d). L. acidophilus Y-01 pada Garam Empedu 35 Grafik di atas merupakan populasi BAL yang dihitung berdasarkan persamaan hubungan antara nilai absorbansi dan hasil pemupukan. Nilai optical density yang didapat merupakan populasi bakteri hidup dan mati yang ditangkap oleh spektrum cahaya dalam spektrofotometer, namun dilakukan juga pemupukan, yang hasilnya merupakan populasi bakteri hidup Pertumbuhan S. thermophilus S-01 yang diberi perlakuan garam empedu dan kontrol dapat dilihat pada grafik pertama. Terlihat pada waktu pengamatan awal, populasi S. thermophilus S-01 masih dapat menyamai kontrol dan terlihat statis. Terlihat dari kisaran populasi yang walaupun naik dan turun, namun masih pada kisaran 107-108 cfu/ml. Populasi L. bulgaricus S-01 dengan penambahan garam empedu dapat dikatakan stabil, seiring dengan pertumbuhan L. bulgaricus S-01 kontrol. Populasi L. bulgaricus S-01 ditambah garam empedu mulai mengalami kenaikan setelah 15 jam inkubasi. Dapat dikatakan, L. bulgaricus S-01 sudah mulai beradaptasi dengan kondisi garam empedu. Dari grafik tersebut menunjukkan bahwa L. bulgaricus S-01 dapat bertahan pada kondisi garam empedu, walaupun mengalami penurunan populasi. Pertumbuhan bakteri B. longum Y-01 pada perlakuan garam empedu dan yang tidak ditambahkan garam empedu (kontrol). Populasi yang didapat antara kontrol dan perlakuan menunjukkan hasil yang tidak jauh berbeda. Terlihat pada kurva, bahwa populasi B. longum Y-01 pada garam empedu dapat bertahan, bahkan hampir menyamai populasi kontrol. Ini menunjukkan, bahwa B. longum Y-01 dapat bertahan pada lingkungan garam empedu. Adaptasi Bifidobacteria untuk garam empedu, biasanya dengan konsentrasi dibawah 5 mM, ini merupakan hal yang penting dalam kolonisasi saluran pencernaan (Ruiz et al., 2009). Karena itu, ketahanan B. longum dalam usus akan mempengaruhi populasi B. longum dalam kolon. Pertumbuhan L. acidophilus Y-01 yang diberi perlakuan garam empedu dan kontrol. Perubahan populasi L. acidophilus Y-01 dengan penambahan garam empedu lebih stabil dibandingkan kontrol. L. acidophilus Y-01 bertahan dalam kondisi saluran pencernaan yaitu usus, di mana garam empedu tersebut disekresikan. 36 Menurut Sanders (2000), Lactobacillus dan Bifidobacterium secara umum lebih resisten terhadap garam empedu dibandingkan dengan genus Streptococcus dan genus lainnya, terutama bakteri asam laktat yang digunakan sebagai kultur starter yogurt. Namun secara keseluruhan, keempat bakteri asam laktat indigenous susu sapi segar ini masih dapat hidup pada kondisi garam empedu. Jacobsen et al. (1990), semua mikroba yang berhasil hidup setelah ditumbuhkan dalam MRSA yang ditambah 0,3% oxgall, dinyatakan bersifat tahan terhadap garam empedu. Ketahanan Kultur Starter BAL Indigenous Susu Sapi Segar pada Antibiotik Berbeda Syarat bakteri asam laktat dikatakan sebagai bakteri probiotik adalah dapat tahan terhadap antibiotik. Antibiotik digunakan dalam dunia kedokteran untuk pengobatan suatu penyakit yang disebabkan bakteri. Terapi obat dengan antibiotik biasa dilakukan pada orang dewasa atau usia lanjut yang mengalami gangguan atau penyakit yang bervariasi. Semakin bertambahnya usia, bakteri baik yang tumbuh secara alami dalam saluran pencernaan pun mengalami penurunan populasi. Karena itu diperlukan pangan yang mengandung bakteri baik agar mikroflora dalam usus antara bakteri menguntungkan dan patogen tetap seimbang. Populasi BAL indigenous susu sapi segar terhadap amoksisilin sebelum dan sesudah inkubasi 24 jam disajikan pada Tabel 5. Tabel 5. Jumlah Populasi BAL Indigenous Susu Sapi Segar pada Amoksisilin 1. S. thermophilus S-01 Populasi BAL (Log10 cfu/ml) P0 jam P24 jam (P24 - P0)* 7,78 ± 0,92 5,49 ± 0,35 -2,32 ± 1,64 2. L. bulgaricus S-01 7,72 ± 0,05 6,23 ± 0,66 -1,49 ± 1,05 3. B. longum Y-01 7,65 ± 0,13 7,82 ± 0,10 0,17 ± 0,12 4. L. acidophilus Y-01 7,82 ± 0,79 8,20 ± 0,37 0,38 ± 0,27 No Kultur bakteri Keterangan : * apabila hasilnya (-) menunjukan adanya kematian; ** huruf superskript yang berbeda pada baris yang sama menunjukkan berbeda nyata (P<0,05) Antibiotik yang sering digunakan dalam dunia kedokteran untuk mengobati berbagai macam penyakit adalah amoksisilin dan kloramfenikol. Penggunaan antibiotik ini adalah karena keduanya mempunyai spektrum luas terhadap aktivitas 37 bakteri Gram positif dan Gram negatif. Namun, cara kerja amoksisilin dan kloramfenikol terhadap bakteri-bakteri tersebut berbeda. Tabel 5 menunjukkan hasil bahwa keempat bakteri tersebut dapat tahan terhadap amoksisilin. Hasil pada Tabel 5 menunjukkan B. longum Y-01 dan L.acidophilus Y-01 mengalami kenaikan populasi, sedangkan S. thermophilus S-01 dan L. bulgaricus S-01 mengalami penurunan populasi setelah diinkubasi selama 24 jam. S. thermophilus S-01 mengalami penurunan populasi sekitar 2,3 log10 cfu/ml, sedangkan L. bulgaricus S-01 mengalami penurunan populasi sekitar 1,4 log10 cfu/ml. Hasil ini menunjukkan ketahanan B. longum Y-01 dan L. acidophilus Y-01 lebih baik dibandingkan dengan S. thermophilus S-01 dan L. bulgaricus S-01. Formulasi amoksisilin adalah mengandung antibiotik semisintetik, analog ampisilin, dengan spektrum yang luas dari aktivitas bakterisidal terhadap mikroorganisme Gram negatif dan Gram positif. Alcamo (1984) juga menyatakan bahwa amoksisilin bekerja pada spektrum luas, terutama dalam menghambat bakteri Gram positif. Kline (2009), mengatakan untuk mengurangi perkembangan bakteri resisten terhadap obat dan menjaga efektivitas amoksisilin dan obat antibakterial lain, amoksisilin harus digunakan hanya untuk mengobati atau mencegah infeksi yang terbukti atau diduga disebabkan oleh bakteri. Penurunan populasi ini disebabkan karena amoksisilin dapat menghambat biosintesis mukopeptida dinding sel dan menghambat komponen peptidoglikan pada dinding sel. Lapisan peptidoglikan terdiri dari unit-unit glikan tetrapeptida yang membentuk suatu polimer, yaitu mukopeptida. Peptidoglikan ini terdiri dari turunan gula yaitu N-asetilglukosamin dan N-asetilmuramat serta beberapa asam amino yaitu L-alanin, D-alanin, asam D-glutamat dan asam diaminopimelat (ADP). Peptidoglikan yang tebal dan ikatan rantai asam teikoat inilah yang dapat mempertahankan bentuk dinding sel dari kondisi asam ekstraseluler (Fardiaz, 1989). Lapisan peptidoglikan juga berperan penting dalam menstabilkan sel bakteri. Jika lapisan peptidoglikan rusak, maka pori-pori pada dinding sel membesar, bentuk sel tidak kaku, tidak tahan terhadap kondisi ekstraseluler yang ekstrim, sehingga terjadi permeabilitas sel dan materi intraseluler keluar. Sitoplasma dan ribosom yang keluar mengakibatkan sel tidak dapat melakukan metabolisme dan sintesis protein, dinding sel menjadi rusak dan tidak dapat melindungi materi dalam sel. Amoksisilin aktif 38 pada beberapa strain mikroorganisme tertentu seperti mikroorganisme Gram positif dan negatif yang bersifat aerobik (Kline, 2009). Cara kerja amoksisilin mirip dengan ampisilin dalam menghambat aksi bakterisidal terhadap organisme yang rentan selama pembelahan masih aktif. Amoksisilin bersifat bakterisidal, yaitu membunuh bakteri secara langsung, dengan menghambat pembentukan lapisan khusus di seluruh bagian tubuh bakteri. Lapisan ini berperan penting dalam melindungi bakteri dari kondisi lingkungan dan agar tubuh bakteri tidak tercerai –berai (Schunack et al., 1990). Populasi BAL indigenous susu sapi segar terhadap kloramfenikol sebelum dan sesudah inkubasi 24 jam dapat dilihat pada Tabel 6. Tabel 6. Jumlah Populasi BAL Indigenous Susu Sapi Segar pada Kloramfenikol 1. S. thermophilus S-01 Populasi BAL (Log10 cfu/ml) P0 jam P24 jam (P24 - P0)* 7,70 ± 0,03 8,27 ± 0,42 0,58 ± 0,41 2. L. bulgaricus S-01 7,29A ± 0,02 8,04B ±0,08 0,76 ± 0,54 3. B. longum Y-01 7,92A ± 0,03 8,12B ±0,03 0,20 ± 0,14 4. L. acidophilus Y-01 7,73a ± 0,05 8,23b ± 0,11 0,50 ± 0,35 No Kultur bakteri Keterangan : * apabila hasilnya (-) menunjukan adanya kematian; ** huruf superskript (a dan b) yang berbeda pada baris yang sama menunjukkan berbeda nyata (P<0,05) *** huruf superskript (A dan B) yang berbeda pada baris yang sama menunjukkan berbeda sangat nyata (P<0,01) Selain diujikan pada amoksisilin, bakteri asam laktat indigenous susu sapi segar juga diujikan pada antibiotik kloramfenikol. Kloramfenikol memiliki spektrum yang luas terhadap aktivitas bakteri Gram positif dan negatif. Namun, di Amerika serikat, kloramfenikol dilarang penggunaannya pada hewan yang akan dikonsumsi (USDA, 2003). Penggunaan berlebihan kloramfenikol dapat menyebabkan karsinogen pada manusia dan bersifat bakterisidal pada bakteri. Namun, kloramfenikol memang sangat efektif menghambat Gram positif, Gram negatif, riketsia dan fungi. Bahkan kloramfenikol tetap dipertahankan karena penting dalam pengobatan berbagai penyakit. Semua bakteri asam laktat mampu bertahan dalam media yang ditambahkan kloramfenikol (Tabel 6), bahkan mengalami kenaikan populasi dari awal pengujian 39 sampai masa inkubasi 24 jam. Antibiotik ini juga berpengaruh secara nyata terhadap populasi awal dan akhir BAL L. acidophilus Y-01 (P>0,05) dan berpengaruh sangat nyata (P<0,01) pada BAL L. bulgaricus S-01 dan B. longum Y-01. Kenaikan populasi semua BAL mulai dari 0,2–0,7 log10 cfu/ml. Cara kerja kloramfenikol bersifat bakteriostatik, yaitu menghambat pertumbuhan bakteri. Kerja kloramfenikol dengan menghambat sintesis protein pada bakteri dengan mengikat subunit pada ribosom dan menahan beberapa enzim yang dihasilkan dalam sintesis protein. Ribosom berfungsi untuk sintesis protein. Penyusun utama bakteri adalah protein, yaitu sekitar 15% berat sel. Jika sintesis protein terhambat, maka protein tidak dihasilkan untuk menyusun lapisan tubuh dan membentuk ikatan peptida pada bakteri. Asam amino dari protein dibutuhkan untuk membentuk lapisan peptidoglikan yang kaku untuk mempertahankan bentuk sel. Ribosom pada sel mitokondria mamalia juga berpengaruh terhadap jaringan perkembangbiakan, misalnya sistem hematopoietik sebagai toksisitas kloramfenikol (USDA, 2003). Sebagian besar bakteri asam laktat lebih sensitif terhadap antibiotik jenis kloramfenikol dibandingkan dengan amoksisilin, terlihat dari jumlah kematian dan penurunan populasi (Firmansyah, 2009). Tetapi bakteri asam laktat S.thermophilus S-01, L. bulgaricus S-01, B. longum Y-01 dan L. acidophilus Y-01 menunjukkan lebih resisten pada antibiotik kloramfenikol. Perbedaan yang terjadi pada masingmasing antibiotik disebabkan antibiotik mempunyai cara kerja yang berbeda-beda. Galur yang berbeda pada tiap bakteri asam laktat mempunyai mekanisme yang berbeda terhadap berbagai perlakuan yang diberikan. Galur bakteri asam laktat yang sama serta diisolasi dari sumber yang sama pun, mempunyai keragaman toleransi terhadap kondisi lingkungan yang dapat menyebabkan stress (Chou dan Weimer, 1999). Perubahan populasi antara kontrol, amoksisilin dan kloramfenikol setiap satu jam selama pengamatan 24 jam dapat dilihat pada Gambar 7. Keempat grafik menunjukkan ketahanan bakteri terhadap kloramfenikol lebih baik dibandingkan pada amoksisilin. Beberapa jam pada awal perlakuan populasi amoksisilin hampir sama dengan kloramfenikol, bahkan pada L. bulgaricus S-01 nilai absorbansi amoksisilin lebih tinggi dibandingkan dengan kloramfenikol. 40 Keterangan : kontrol ( ), amoksisilin ( ), kloramfenikol ( ) Gambar 7. Grafik Kurva Pertumbuhan Kultur Starter a). S. thermophilus S-01, b). L. bulgaricus S-01, c). B. longum Y-01, d). L. acidophilus Y-01 pada Amoksisilin dan Kloramfenikol 41 Cara kerja kloramfenikol dan amoksisilin terhadap masing-masing bakteri asam laktat berbeda-beda. Firmansyah (2009), melakukan penelitian bakteri asam laktat yang diujikan dalam antibiotik kloramfenikol dan amoksisilin, dan hasilnya adalah bakteri asam laktat lebih tahan tumbuh pada amoksisilin dibandingkan pada kloramfenikol. Ini menunjukkan mekanisme bakteri asam laktat terhadap perlakuan yang diberikan berbeda, setiap galur mempunyai mekanisme berbeda dalam memberikan reaksi terhadap penghambatan yang dilakukan oleh antibiotik (Chou dan Weimer, 1999). Kontrol (tidak ditambahkan antibiotik) pada setiap jamnya terjadi kenaikan populasi. Hasil ini membuktikan bahwa antibiotik dapat menghambat pertumbuhan bakteri asam laktat. Antibiotik merupakan senyawa antibakteri alami yang dihasilkan oleh beberapa mikroorganisme untuk menghambat mikroorganisme lain. Antibiotik sintesis juga sudah banyak digunakan dalam dunia kedokteran, terutama dalam pengobatan penyakit karena bakteri. Antibiotik memang digunakan untuk membidik bakteri Gram negatif atau Gram positif serta melakukan penghambatan terhadap proses biokimia bakteri, terutama dalam proses infeksi oleh bakteri (Pelczar dan Chan, 2008). Aktivitas Antimikroba Bakteri Asam Laktat Indigenous Susu Sapi Segar terhadap Bakteri Patogen Salah satu karakteristik suatu bakteri asam laktat dapat dikategorikan sebagai probiotik adalah memiliki aktivitas antimikroba terhadap bakteri patogen. Sifat antimikroba adalah suatu kemampuan antagonistik suatu senyawa kimia untuk menghambat pertumbuhan mikroba yang tidak diinginkan. Kemampuan penghambatan ini disebabkan oleh produksi senyawa antimikroba berupa asam laktat dan metabolit lain seperti bakteriosin, hidrogen peroksida dan asam lemak rantai pendek (Drago et al., 1997). Mekanisme penghambatan pertumbuhan mikroba oleh senyawa antimikroba bakteri asam laktat antara lain (1) perusakan dinding sel sehingga mengakibatkan lisis atau menghambat pembentukan dinding sel yang sedang tumbuh, (2) mengubah permeabilitas membran sitoplasma sehingga terjadi kebocoran nutrien dalam sel dan (3) merusak sistem metabolisme dengan cara menghambat kerja enzim intraseluler (Widiasih, 2008). 42 Penggunaan S. Typhimurium, E. coli, dan S. aureus ini adalah karena patogen tersebut sering menyerang manusia dan menjadi penyebab penyakit. S.Typhimurium merupakan patogen penyebab tipus, gangguan perut dan infeksi pada usus. E. coli merupakan bakteri alami dalam saluran pencernaan manusia, dengan jumlah terbatas. Jika jumlahnya melebihi batas dan lebih besar dibandingkan dengan bakteri probiotik, maka E. coli dapat menghasilkan keton yang merupakan racun bagi manusia. Keton kemudian mengikis permukaan usus dan menyebabkan diare, bahkan diare berdarah. S. aureus terdapat secara alami pada sel pernafasan dan kulit. S. aureus merupakan penyebab keracunan makanan, yang mengakibatkan penderita mengalami diare, muntah dan kejang (Todar, 2008; Drago et al., 1997) Aktivitas antagonistik BAL terhadap S. Typhimurium ATCC 14028 dapat dilihat pada Tabel 7. Bakteri asam laktat yang berbeda dari indigenous susu sapi segar tidak mempengaruhi besarnya diameter zona hambat yang dihasilkan masingmasing bakteri asam laktat S.Typhimurium (P>0,05). yang dikonfrontasikan pada bakteri patogen Tiap bakteri asam laktat indigenous susu sapi segar menghasilkan zona bening di sekitar lubang dan menunjukkan adanya aktivitas antimikroba pada bakteri asam laktat yang diujikan terhadap S. Typhimurium. Tabel 7. Aktivitas Antagonistik BAL Indigenous Susu Sapi Segar terhadap S. Typhimurium ATCC 14028 No 1. 2. 3. 4. Kultur Bakteri Diameter penghambatan (mm) S. thermophilus S-01 15,0 ± 0,4 L. bulgaricus S-01 11,0 ± 0,7 B. longum Y-01 13,1 ± 0,2 L. acidophilus Y-01 15,4 ± 0,3 Keterangan : besarnya diameter lubang 7 mm Besarnya zona hambat S. thermophilus S-01, L. bulgaricus S-01, B. longum Y-01 dan L. acidophilus Y-01 berbeda-beda. L. bulgaricus S-01 menghasilkan zona hambat yang paling kecil dibanding dengan S. thermophilus S-01, B. longum Y-01 dan L. acidophilus Y-01. Ini menunjukkan S. Typhimurium lebih sensitif terhadap produksi antimikroba S. thermophilus S-01, B. longum Y-01 dan L. acidophilus Y01. 43 Antimikroba bakteri asam laktat yang diisolasi dari produk susu seperti yogurt dan yogurt probiotik mempunyai aktivitas antimikroba. Chuayana et al. (2003), menyatakan bakteri yang diisolasi dari produk yogurt mengandung bakteri asam laktat Streptococcus spp., Lactobacillus spp. dan Bifidobacterium spp. dan menghasilkan zona hambat terhadap semua patogen, termasuk S. Typhimurium. S. Typhimurium mampu dihambat oleh asam propionat, asam asetat dan asam laktat. Senyawa metabolit yang paling penting dalam menginaktivasi Salmonella adalah asam laktat (Ray, 2004). Ini dikarenakan S. Typhimurium tumbuh pada pH netral 6,6–8,2. Kondisi pH >9,2 dan <4,0 akan menyebabkan kematian bakteri ini. Keempat bakteri ini adalah bakteri asam laktat, dan menghasilkan molekul organik sebagai adanya aktivitas antimikroba. S. thermophilus memproduksi asam laktat dalam bentuk L(+), asetildehida dan diasetil dari laktosa, sedangkan L. bulgaricus menghasilkan asam laktat D(-), asetaldehida dan hidrogen peroksida. L. acidophilus dan L. bulgaricus memproduksi bakteriosin dengan tingkat kesensitifan yang bervariasi (Nowroozi et al., 2004). Menurut Surono (2004), dibandingkan dengan asam laktat D(-), asam laktat L(+) memberikan penghambatan yang lebih kuat terhadap bakteri yang tidak diinginkan. Ini dapat dibuktikan dengan diameter yang dihasilkan S. thermophilus S-01 lebih besar dibandingkan dengan L. bulgaricus S-01. Surono (2004) menambahkan, efek antimikroba merupakan akibat dari penurunan pH dan bentuk asam organik yang tidak terdisosiasi. Asam tidak terdisosiasi ini dapat menembus dinding sel bakteri Gram negatif yang memiliki kandungan lipid tinggi, yaitu 11-22%. Kondisi asam ekstraseluler mengakibatkan pori-pori dinding sel bakteri Gram negatif membesar dan terjadi permeabilitas sel, sehingga materi intraseluler banyak keluar. Kondisi pH ekstraseluler yang rendah mengakibatkan asidifikasi sel sitoplasma dan asam yang tidak terdisosiasi menjadi lipofilik sehingga dapat berdifusi ke dalam membran. Keadaan ini yang akan mengganggu sistem transport substrat dalam sel, sehingga tidak ada pasokan nutrien untuk proses metabolisme sel. Senyawa antimikroba yang dihasilkan BAL selain asam organik adalah bakteriosin dan hidrogen peroksida (H2O2). Target utama bakteriosin yang diproduksi adalah membran sitoplasma, karena bakteriosin memulai reaksi-reaksi yang mengubah permeabilitas membran sehingga mengganggu transpor membran 44 atau menghilangkan tenaga gerak proton yang menghasilkan terhambatnya produksi energi dan biosintesis protein atau asam nukleat (Nissen-Meyer et al., 1992). Hidrogen peroksida merupakan hasil aerobik bakteri, bekerja menghambat pertumbuhan bakteri karena bersifat toksik bagi bakteri yang tidak tahan. Uji katalase menunjukkan S. Typhimurium ATCC 14028 termasuk dalam katalase positif, yaitu bakteri ini mempunyai enzim katalase untuk memecah H2O2 menjadi H2 dan O2, sehingga bakteri ini mampu bertahan terhadap hidrogen peroksida yang dihasilkan BAL (Hadioetomo, 1993). Diameter zona bening yang terbentuk sebagai aktivitas antagonistik BAL terhadap E.coli ATCC 25922 dapat dilihat pada Tabel 8. Tabel 8. Aktivitas Antagonistik BAL Indigenous Susu Sapi Segar terhadap E. coli ATCC 25922 No Kultur Bakteri Diameter penghambatan (mm) 1. S. thermophilus S-01 15,6 ± 0,7 2. L. bulgaricus S-01 11,0 ± 0,1 3. B. longum Y-01 4. L. acidophilus Y-01 14,7 ± 0,4 15,1 ± 0,4 Keterangan : Besarnya diameter lubang 7 mm Besarnya diameter zona hambat terhadap bakteri patogen E. coli tidak dipengaruhi oleh bakteri asam laktat indigenous susu sapi segar yang berbeda (P>0,05). Diameter zona bening terlihat di sekitar sumur, dan menunjukkan adanya aktivitas antimikroba yang dihasilkan setiap bakteri asam laktat. Berdasarkan Tabel 8 zona hambat L. bulgaricus S-01 lebih kecil dibandingkan S. thermophilus S-01, B. longum Y-01 dan L. acidophilus Y-01. Ini menunjukkan E. coli kurang sensitif terhadap aktivitas antimikroba yang dihasilkan L. bulgaricus S-01. Namun, menurut Nuryani (2006), E. coli merupakan bakteri Gram negatif yang paling sensitif terhadap aktivitas antimikroba bakteri asam laktat yogurt, dengan rataan diameter zona hambat 8,05±0,18 (diameter sumur 5 mm), jika dibandingkan dengan S.Typhimurium dan Pseudomonas sp. Dibandingkan dengan penelitian yang dilakukan Nuryani (2006), diameter penghambatan zona bening BAL hasil isolasi susu sapi segar ini lebih besar, ini dikarenakan filtrat bebas sel yang dikonsentrasikan 45 hingga 5 kali dari volume awal, sehingga senyawa antimikroba yang dihasilkan lebih maksimal. E. coli tumbuh optimum pada pH 7,0-7,5, dengan pH minimum 4 dan pH maksimum 8,5. Asam laktat yang tak terdisosiasi dapat menghambat pertumbuhan E. coli (Cotter and Hill, 2003). Asam-asam lipofilik seperti asam laktat dan asetat dalam bentuk tidak terdisosiasi dapat menembus sel antimikroba dan pada pH intraseluler yang lebih tinggi, berdisosiasi menghasilkan ion-ion hidrogen dan mengganggu fungsi metabolik essensial seperti translokasi substrat dan fosforilaso oksidatif, dengan demikian mereduksi pH intraseluler (Naidu dan Clemens, 2000). Bakteri asam laktat yang dikonfrontasikan menghasilkan asam, sehingga bakteri patogen disekitar sumur menjadi terhambat pertumbuhannya. S. thermophilus, L. bulgaricus, B. longum dan L. acidophilus digunakan dalam memfermentasi yogurt probiotik, yang menghasilkan metabolit antimikroba yaitu asam asetat dan asam laktat. antimikroba tersebut. E. coli sangat sensitif terhadap metabolit Keempat bakteri ini adalah bakteri asam laktat dan menghasilkan molekul organik sebagai adanya aktivitas antimikroba. S.thermophilus memproduksi asam laktat dalam bentuk L(+), asetildehida dan diasetil dari laktosa, sedangkan L. bulgaricus menghasilkan asam laktat D(-), asetaldehida dan hidrogen peroksida (Nowroozi et al., 2004) Bakteri Gram negatif lebih sensitif terhadap metabolit antimikroba dibandingkan dengan bakteri Gram positif. Asam laktat yang dihasilkan dapat melemahkan permeabilitas bakteri Gram negatif dengan merusak membran luar bakteri Gram negatif. Bakteri Gram negatif tidak mempunyai mekanisme pompa proton yang mampu menyeimbangkan pH di dalam sel dan substrat antimikroba lain tidak dapat berpenetrasi ke dalam membran sitoplasma (Cotter dan Hill, 2003). Kerusakan yang terjadi pada membran luar bakteri, terjadi karena bakteri Gram negatif seperti E. coli ini mempunyai lapisan dinding sel yang tipis dan lipid yang tebal mencapai 11-22%. Keadaan asam ekstraseluler membuat pori-pori sel membesar, terjadi peningkatan permeabilitas sel, sehingga materi yang tidak dibutuhkan untuk metabolisme masuk ke dalam sel, sedangkan materi intraseluler sel banyak yang keluar. Materi intraseluler seperti sitoplasma dan ribosom tidak dapat melakukan fungsinya untuk mengangkut nutrien dan mensintesis protein. 46 Selain sensitif terhadap asam organik, bakteri patogen juga dapat dihambat dengan bakteriosin dan hidrogen peroksida. Uji katalase yang dilakukan menunjukkan bahwa E. coli ATCC 25922 ini termasuk dalam katalase positif. Bakteri E. coli mempunyai enzim katalase untuk memecah H2O2 menjadi H2 dan O2, karena H2O2 ini dapat menjadi toksik bagi bakteri E. coli itu (Hadioetomo, 1993). Diameter penghambatan yang merupakan aktivitas antagonistik BAL terhadap S.aureus ATCC 25923disajikan pada Tabel 9. Tabel 9. Aktivitas Antagonistik BAL Indigenous Susu Sapi Segar terhadap S. aureus ATCC 25923 No Kultur Bakteri Diameter penghambatan (mm) 1. S. thermophilus S-01 14,6 ± 0,4 2. L. bulgaricus S-01 12,8 ± 0,3 3. B. longum Y-01 13,8 ± 0,5 4. L. acidophilus Y-01 11,8 ± 0,1 Keterangan : Besarnya diameter lubang 7 mm Perbedaan jenis bakteri asam laktat tidak mempengaruhi besarnya diameter zona hambat terhadap bakteri patogen S. aureus. Zona hambat yang terbentuk menunjukkan adanya aktivitas antimikroba dalam menghambat bakteri S. aureus. Bakteri probiotik mampu menghambat bakteri Gram positif dan negatif, termasuk S. aureus (Nowroozi et al., 2004). S. aureus tumbuh pada pH 4,0–8,0, produksi asam laktat akan menurunkan pH disekitar S. aureus, dan pada kondisi pH rendah, pertumbuhan bakteri ini terhambat. Selain itu, S. aureus sensitif terhadap metabolit antimikroba yang dihasilkan bakteri asam laktat, antara lain asam asetat dan asam laktat. Asam-asam organik yang dihasilkan oleh bakteri asam laktat yang mengakibatkan akumulasi produk akhir asam dan turunnya pH yang menyebabkan penghambatan yang luas terhadap bakteri Gram positif dan negatif. L. plantarum, L. bulgaricus, L. acidophilus dan L. casei yang dapat memproduksi senyawa antimikroba dibandingkan 28 jenis Lactobacilli lainnya yang dikonfrontasikan terhadap bakteri patogen. Sensitifitas masing-masing bakteri juga sangat bervariasi (Nowroozi et al., 2004). Senyawa antimikroba yang dihasilkan L.bulgaricus adalah bakteriosin, yang mempunyai aktivitas antimikroba terhadap 47 beberapa penyebab pembusukan makanan dan bakteri patogen antara lain L. monocytogenes, Y. enterocolitica, S. aureus, B. subtilis dan S. Typhimurium. Semakin luas zona bening yang terlihat, maka makin sensitif bakteri patogen yang diujikan. S. aureus adalah bakteri Gram positif yang paling sensitif terhadap antimikroba bakteri asam laktat yogurt, jika dibandingkan dengan Bacillus cereus. Rataan diameter zona hambat Gram positif adalah 6,81±0,11 (diameter sumur 5 mm) (Nuryani, 2006). Struktur dinding sel bakteri Gram positif memiliki satu lapisan tebal peptidoglikan, sedangkan Gram negatif relatif lebih kompleks dengan tiga lapisan berupa lipoprotein (lapisan luar), lipolisakarida (lapisan tengah) dan peptidoglikan (lapisan dalam). Kandungan lipid pada dinding sel Gram positif hanya sekitar 1-4%, lebih tipis dibandingkan dengan kandungan lipid Gram negatif. Perbedaan lapisan ini mempengaruhi penghambatan terhadap bakteri asam laktat yang diujikan. Menurut Nowroozi et al. (2004), L. acidophilus dan L. bulgaricus memproduksi bakteriosin dan mempunyai tingkat kesensitifan yang sangat bervariasi. Maksimum aktivitas antimikroba adalah pada akhir fase pertumbuhan log dan awal fase statis (Nowroozi et al., 2004; Oh et al., 2000). Zona hambat yang dihasilkan BAL terhadap ketiga bakteri secara keseluruhan menunjukkan bahwa senyawa antimikroba yang dihasilkan memang dapat menghambat pertumbuhan bakteri patogen. Beberapa senyawa antimikroba memang dihasilkan oleh BAL, namun dalam penelitian ini diduga asam organiklah yang berperan besar dalam menghambat pertumbuhan patogen. Ini dikarenakan filtrat bebas sel terkonsentrasi yang digunakan mempunyai pH sekitar 4,0-4,2. Tetapi, tidak menutup kemungkinan bahwa zona hambat yang terbentuk juga disebabkan oleh hidrogen peroksida, bakteriosin atau senyawa antimikroba lainnya. Diameter penghambatan yang terbentuk pada bakteri asam laktat yang biasa digunakan untuk pembuatan yogurt, yaitu S. thermophilus dan L. bulgaricus, mempunyai besar penghambatan yang berbeda. Diameter penghambatan S.thermophilus terhadap ketiga bakteri patogen, lebih besar daripada L. bulgaricus. Ini terkait dengan jenis asam laktat yang dihasilkan kedua bakteri ini. S.thermophilus menghasilkan asam laktat L(+), sedangkan L. bulgaricus menghasilkan asam laktat D(-). Menurut Surono (2004), asam laktat L(+) mempunyai kemampuan menghambat yang lebih kuat terhadap bakteri-bakteri yang tidak diinginkan, 48 dibanding dengan asam laktat D(-). Asam laktat (+) maksimal dihasilkan pada awal fase pertumbuhan, sedangkan asam laktat D(-) dihasilkan pada akhir fase pertumbuhan. Dalam pangan, asam laktat L(+) lebih cepat dimetabolit dan lebih mudah diserap tubuh. Hasil rekapitulasi karakteristik BAL untuk menjadi kandidat bakteri probiotik melalui pengujian pada kondisi saluran pencernaan, khususnya pada kondisi lambung dan usus secara in vitro, disajikan dalam Tabel 10. Tabel 10. Rekapitulasi Karakteristik BAL Indigenous Susu Sapi Segar sebagai Kandidat Bakteri Probiotik Kultur starter S. thermopilus pH 2 2,5 - - S-01 L. bulgaricus S-01 B. longum Y-01 Garam 7,2 empedu + + - (75%) (106%) 3,2 Y-01 Amoksisilin Antimikroba Kloramfenikol + + (71%) (107%) + + + + + + + (97%) (84%) (94%) (89%) (72%) (81%) (110%) + + + + + + + (106%) (101%) (102%) (103%) + + + + (105%) (103%) (105%) (106%) (102%) (103%) (106%) L. acidophilus Antibiotik + + + (101%) (103%) (104%) + + + + Keterangan : (-) : BAL tidak tahan dan tidak terdapat pertumbuhan (+) : BAL mampu bertahan ditunjukkan oleh (%) bakteri hidup Hasil rekapitulasi menunjukkan S. thermophilus tidak tahan pada kondisi pH 2 dan 2,5 serta garam empedu. Ini berarti S. thermophilus tidak tahan di dalam lambung dan usus halus (duodenum). Padahal, salah satu syarat BAL menjadi kandidat bakteri probiotik adalah tahan pada kondisi asam dalam lambung dan tahan garam empedu. probiotik. Karena itu, S. thermophilus bukan sebagai kandidat bakteri Ketiga BAL lain, yaitu L. bulgaricus S-01, B. longum Y-01 dan L.acidophilus Y-01 dapat dikatakan sebagai kandidat bakteri probiotik karena kemampuannya bertahan pada semua kondisi pH, garam empedu, antibiotik dan mampu menghasilkan senyawa antimikroba dalam menghambat bakteri patogen. Kemampuan BAL bertahan atau tidak dalam kondisi pH yang berbeda, garam empedu dan antibiotik, dipengaruhi oleh lapisan dinding sel masing-masing bakteri. Lapisan dinding sel bakteri Gram positif lebih tebal dibanding Gram negatif, namun 49 mempunyai lapisan lipid yang lebih tipis sekitar 1-4%. Dinding sel Gram positif terdiri dari 90% peptidoglikan dan 10% asam teikoat. Lapisan lipid yang tipis menyebabkan pori-pori sel menjadi mengecil saat asam, garam dan antibiotik melakukan penghambatan terhadap BAL. Hal ini menghambat terjadinya permeabilitas sel yang berlebihan dan tidak terjadi ekstraksi materi intraseluler sel. Lapisan dinding sel inilah yang menyebabkan BAL mampu bertahan dalam kondisi ektraseluler yang berbeda dengan intraseluler. Selain itu, dibandingkan dengan Gram negatif, bakteri Gram positif juga mempunyai kemampuan mempertahankan pH sitoplasma lebih basa daripada pH ekstraseluler. Setiap galur bakteri mempunyai ketahanan yang berbeda terhadap asam atau pH rendah, contohnya Lactobacillus lebih toleran terhadap pH rendah dibandingkan Lactococci dan Streptococci. Dibandingkan dengan Streptococci, galur Lacto- bacillus dan Bifidobacteria memang lebih toleran pada kondisi asam dan garam empedu (Farida, 2006; Zinedine dan Faid, 2007). L. bulgaricus S-01 lebih optimal tumbuh pada kondisi asam, sedangkan S. thermophilus S-01 lebih optimal tumbuh pada kondisi pH mendekati normal (Oh et al., 2000). 50