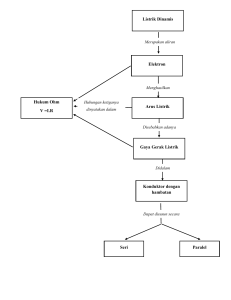
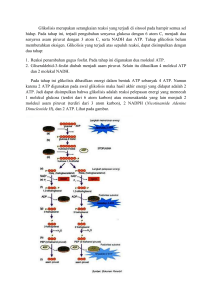
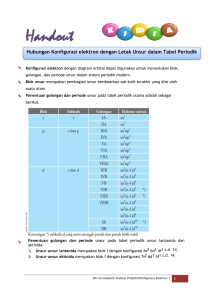
MAKALAH BIOKIMIA (METABOLISME KARBOHIDRAT) OLEH: KELOMPOK :IV (EMPAT) NAMA :AFDIN (181431312) :MAURA KHUMAIRA (181421341) :RAHMAWATI (181431356) :HALMA (181421334) :WA ITI LA ANDA (181421375) PROGRAM STUDI PENDIDIKAN BIOLOGI FAKULTS KEGURUAN DAN ILMU PENDIDIKAN UNIVERSITAS SEMBILANBELAS NOVEMBER KOLAKA 2019 KATA PENGANTAR Assalamu’alaikum Wr.Wb Puji syukur kita marilah kita panjatkan kehadirat Allah SWT yang telah banyak memberikan beribu-ribu nikmat kepada kita umatnya. Rahmat beserta salam semoga tetap tercurahkan kepada junjungan kita, pemimpin akhir zaman yang sangat dipanuti oleh pengikutnya yakni Nabiullah Muhammad SAW. “ Metabolisme Karbohidrat” ini sengaja dibahas karena sangat penting untuk kita khususnya sebagai mahasiswa yang ingin lebih mengenal tentang biokimia. Selanjutnya, penyusun mengucapkan terimakasih kepada dosen pembimbing mata kuliah ibu Tri Marniata Sari,S.Pd., M.Pd yang telah memberikan pengarahan-pengarahan sehingga kami dapat menyelesaikan makalah ini dengan tepat waktu. Kepada dosen dan teman-teman kami membutuhkan kritik dan saran agar penyusunan makalah ini kedepannya lebih baik lagi. Demikian, semoga makalah ini dapat bermanfaat khususnya bagi penyusun dan umumnya bagi semua yang membaca makalah ini. DAFTAR ISI KATA PENGANTAR ........................................................................................... ii DAFTAR ISI ........................................................................................................ iii 1. Rumusan Masalah ......................................................................................... v 2. Tujuan Penulisan ........................................................................................... v 3. Manfaat Penulisan ......................................................................................... v BAB II ................................................................................................................... vi PEMBAHASAN ................................................................................................... vi A. Pengertian Metabolisme Karbohidrat .......................................................... vi a. Glikolisis ....................................................................................................... vii b. Jalur Asam Sitrat (Siklus Krebs) ............................................................... xi c. Jalur Pentosa Fosfat .................................................................................. xvii d. Jalur Tranfer Elektron .............................................................................. xxi e. Jalur Fosforilasi Oksidatif ....................................................................... xxiv BAB III ............................................................................................................ xxviii PENUTUP ....................................................................................................... xxviii A. KESIMPULAN .......................................................................................... xxviii B. SARAN ....................................................................................................... xxviii DAFTAR PUSTAKA BAB I PENDAHULUAN A. Latar Belakang Kata karbohidrat berasal dari kata karbon dan air. Secara sederhana karbohidrat didefnisikan sebagai polimer gula. Karbohidrat yang paling sederhana adlah aldehid (disebut polihidroksialdehid atau aldosa) atau berupa keton (disebut polihidroksiketon atau ketosa). Karbohidrat terdiri atas atom C,H, dan O. Karbohidrat merupakan senyawa biomolekul yang paling banyak jumlahnya di permukaan bumi. Polimer karbohidrat yang tidak larut merupakan pelindung dan pembentuk energi sel bakteri; pada tumbuhan senyawa ini berfungsi sebagai penopang dan pada hewan berfungsi sebagai jaringan ikat dan “cel coat”. Fungsi utama dari metabolisme karbohidrat adalah untuk menghasilkan energi dalam ntuk senyawa yang mengandung ikatan fosfat bertenga tinggi. Metabolisme karbohidrat yaitu metabolisme mencakup sintesis (anabolisme) dan penguraian (katabolisme) molekul organuk kompleks. Metabilisme biasanya terdiri atas tahapan-tahapan yang melibatkan enzim, yang dikenal pula sebagai jalur metabolisme. Metabolisme total merupakan semua proses biokimia didalam organisme.Metabolisme sel mencakup semua proses kimia didalam sel. Metabolisme karbohidrat dimulali dengan pencernaan amilum dalam usus halus. Hasli pencernaan berupa monosakarida diserap oleh usus halus. Glukosa merupakan senyawa utama yang paling bnayak dibahas dalam metabolisme karbohidrat. Rangkaian reaksi yang membentuk beberapa jalur, seperti glikolisis, siklus asam sitrat, jalur pentose,transfer elektron, hingga fosforilasi oksidatif. 1. Rumusan Masalah Dalam makalah ini, masalah yang akan dibahas adalah: 1. Bagaimana menjelaskan jalur metabolisme karbohidrat ? a. Glikolisis b. Siklus asam sitrat c. Jalur pentose fosfat d. Transfer Elektron e. Fosforilasi Oksidatif 2. Apa yang dimaksud dengan metabolisme ? 2. Tujuan Penulisan 1. Untuk Mengetahui tentang metabolisme karbohidrat 2. Untuk mampu menjelaskan jalur metabolisme karbohidrat mulai: Jalur glikolisis, siklus asam sitrat, jalur pentosa ,jalur transfer elektron dan jalur fosforilasi oksidatif. 3. Manfaat Penulisan 1. Untuk dapat memahami tentang metabolisme karbohidrat,serta 2. Menjelaskan tentang jalur metabolisme karbohidrat mulai dari glikolisis, jalur pentose fosfat, siklus asam sitrat, transfer elektron dan fosforilasi oksidatif. BAB II PEMBAHASAN A. Pengertian Metabolisme Karbohidrat Metabolisme merupakan reaksi dalam sel yang dikatalisis oleh enzimenzim. Pengertian secara luas metabolisme bukanlah suatu proses acak melainkan sangat terintegrasi dan terkoordinasi. Mempunyai tujuan dan mencakup berbagai kerjasama banyak system multi enzim. Sedangkan karbohidrat merupakan zat padat berwarna putih, yang sukar larut dalam pelarut organic, tetapi larut dalam air (kecuali beberapa sakarida) dengan fungsi utama sebagai penghasil energi di dalam tubuh. Pada metabolisme karbohidarat salah satu substansi penting yang akan dipelajari adalah oksidasi glukosa yang disebut dengan respirasi sel. Proses oksidasi ini terjadi secara aerobik. Diamana glukosa ini akan dioksidasi secara lebih lanjut menjadi energi. Karena glukosa termasuk ke dalam senyawa organic yang mengandung karbon, hydrogen dan oksigen, maka dapat di oksidasi secara sempurna menjadi CO2 dan H2O. Okisidasi aerobic glukosa ini memrlukan partisipasi dari lima proses metabolisme yang terkait (glikolisis,siklus asam sitrat,jalur pentosa,transfer elektron,dan fosforilasi oksidatif). Berikut ini jalur metabolisme karbohidrat yaitu: 1. Glikolisis Glikolisis merupakan sebuah rangakaian reaksi biokimia dimana glukosa dioksidaisi menjadi asam piruvat. Dalam keadaaan anaerob piruvat dikonversi menjadi asetil KoA yang kemudian masuk dalam jalur asam trikarboksilat.Sedangkan serangkaian reaksi yang trjadi berurutann dalam jalur EMP untuk mengkonversi glukosa untuk menjadi asam piruvat yang secara garis besar dapat dikelompokkan dalam dua tahap yaitu, perubahan glukosa menjadi triosa fosfat (yang memerlukan energi kimia) dan tahap perubahan triofosfat menjadi asam piruvat sambil melepaskan energi kimia ke lingkungannya. (Gambar proses glikolisis). a. Isomerase Glukosa 6-Fosfat Reaksi berikutnya adalah reaksi isomerase glukosa menjadi fruktosa 6fosfat. Reaksi dan sebaliknya dikatalisis enzim fosfat glukoisomerase (ΔG±1400 kalori,Ph) b. Fosforealasi Fruktosa -6-Fosfat Menjadi Fruktosa 1.6 Difosfat. Pada reaks tahap ketiga ini dikatalisis oleh fosfat- fruktosakinase. Tahap ini merupakan tahap reaksi penting untuk pengendalian metabolisme karena enzim ini adalah enzim alkosterik yang dapat dipengaruhi oleh beberapa metabolit umum. Kelebihan ATP ataupun asam sitrat dapat menghambat enzim fosfofruktokinas ini. Sebaliknya AMP,ADP, dan Fruktosa 6-P dapat menstimulasi enzim.Enzim ini memerlukan ion Mg sebagai kofaktor dan memiliki berat miolekul yang sangat tinggi (±360.000 dan terdiri dari 4 sub unit). c. Pembentukan Trio Fosfa Reaksi berikutnya menyangkut pemotongan glukosa 1,6- difosfat dengan membentuk dua triosa fosfat: dihidroksi aseton fosfat dan Dgliseraldehida-3-fosfat. Enzim yang memngkatalisis rekasi ini adalah aldolase,yang diisolasi pertama kali oleh “Warburg” kini diketahui banyak ditemukan di alam. Garapan yang didapat dari oksidasi aldehida menjadi asam karboksilat disimpan dalam bentuk gugus hasil fosfat 1-3 difosfogliserat. Enzim yang berperan adalah gliseraldehida-3-fosfatdehidrogenase. Berat molekul enzim ini 145.000 dan terdiri atas suatu tetramer dengan berat molekul maisnng-masimg sebanyak 35.000 dan terikat erat dengan NAD+, jadi seluruhnya ada 4 NAD+. d. Interkonversi Asam 3-Fosfogliserat Menjadi 2-Fosfogliserat Fosfogliseril mutase mengkatalisis interkonvensi dua macam asam fosfogliserat. e. Pembentukan Asam Fosfoenol Piruvat Reaksi berikutnya dikatalisis oleh enzim enolase, Tetapan Kesetimbangan rekasi ini sama dengan 3. Hal ini berarti bahwa reaksi diatas berjalan secara reversible. Asam fosfoenol piruvat (PED) merupakan molekul berenergi tinggi. Hidrolisis molekul ini menghasilkan ΔG-14.800 kalori. f. Hidrolisis Asam Fosfoenol Piruvat Menjadi Piruvat Gugus fosfat dari PEP dipindahkan kepada ADP sehingga terbebntuk ATP. Reaksi ini di katalisis oleh enzim piruvat kinase dan menghasilkan energi sebesar 61.000 kalori. Taoutomerisasi dari brntuk enol menjajdi keton dapat memberikan cukup energi untuk membentuk ATP. Satu molekul asam piruvat yang dihasilkan memiliki total energi tersimpan kira-kira sebesar 564 kcal, seadangkan energi yang tersimpan dalam satu molekul glukosa sekitar 686 kcal. Sehingga energy yang tersimpan dalam dua molekul asam piruvat lebih besar daripada satu molekul glukosa sebagai bahan dasarnya. Singkatnya seluruh proses glikolisis melibatkan pemecahan satu molekul glukosa dan menghasilkan 2 molekul NADH, 2 molekul ATP, 2 molekul H2O dan 2 molekul asam piruvat.Produk tersebut selanjutnya digunakan dalam siklus/jalus asam sitrat yang merupakan bagian dari respirasi selular. 2. Jalur Asam Sitrat (Siklus Krebs) Siklus asam sitrat atau siklus asam trikarboksilat atau disebut juga dengan siklus krebs merupakan tahapan kedua dari respirasi aerob. Sesuai dengan namanya penemu siklus ini adalah Sir Hans Krebs (1937), pada kondisi aerob glukosa akan dioksidasi secara sempurna menjadi H2O dan CO2 melalui siklus asam sitrat. Sebelum memasuki siklus asam sitrat, asam piruvat (3 atom karbon) harus dioksidas terlebih dahulu menajdi asetil koenzim A atau asetil Ko-A (2 atom karbon). Reaksi ini terjadi di dalam mitokondria dan dikatalisis oleh enzim piruvat dehydrogenase. (Gambar jalur siklus krebs) Sebagai jalur metabolisme, siklus krebs sangat berperan penting pada ketersediaan ATP yang dieperlukan jaringan. Beberapa fungsi dar siklus krebs diantaranya yaitu sebagai jalur terakhir oksidasi karbohidrat,protein dan lipid yang akan dimetabolisme menjadi Asetil Koenzim-A, menghasilkan sebgaian besar CO2 dengan mengoksidasi glukosa; dan menghasilkan sejumlah koenzim tereduksi yang menggerakan rantai pernapasan untuk memproduksi ATP (Adenosin Trfosfat) serta menyediakan sejumlah bahan untuk kebutuhan sintesis protein dan asam nukleat mengkonversi sejumlah energi zat yang berlebihan untuk digunakan pada sintesis asam lemak sebelum pembentukan trigleserida untuk penimbunan lemak dan juga bertindak sebagai pengendalian langsung maupun tidak langsung terhadap system enzim melalui komponen siklus. Proses dan Tahapan Siklus Krebs Siklus krebs merupakan tahapan kedua setelah glikoslis dari respirasi seluler, siklus ini terjadi di dalam mitokondria sedangkan glikolisis terjadi di dalam sitoplasma. Asam piruvat yang dihasilkan dari proses glikolisis masuk kedalam mitokondria terlebih dulu melalui proses dekarbosilasi oksidatif agar proses siklus jrebs dapat berlangsung. Pada tahap dekarbosilasi oksidatif, asam piruvat akan diubah menjadi Asetil Koenzim-A. Proses pengubahan ini diperantai oleh enzim piruvat dehydrogenase yang ada pada mitokondria sel eukariotik. Berikut adalah tahapan pengubahan asam piruvat menjadi Asetil Ko-A atau disebut dengan dekarbosilasi oksidatif: Pelepas gugus karboksilat (-COO) dari asam piruvat menjadi CO2; Sisa dua atom karbon (Ch3COO-) dari piruvat akan mentrasfer kelebihan elektronnya ke NAD+ sehingga akan terbentuk NADH dan dua molekul tadi akan menjadi asetat. Selanjutnya, Koenzim-A (Ko-A) akan diikatkan pada asetat yang telah terbentuk pada asetat yang telah terbentuk sebelumnya sehingga dihasilkan Asetil Koenzim-A (Ko-A). Asetil Ko-A inilah yang menjadi bahan baku dalam siklus kreb yang berlangsung di mitokondria untuk menghasilkan ATP, NADH, FADH2, dan CO2. Terdapat 8 tahapan dalam siklus krebs diantaranya yaitu sebagai berikut: Tahap I : Sitrat Sintesa Proses yang berlangsung ditahap ini disebut dengan hidrolisis. Pada tahap ini terjadi penggabungan molekul Asetil Ko-A dengan oksaloasetat membentuk asam sitrat dibantu oleh enzim asam sitrat sintase. Tahap II : Isomerase Sitrat Pada tahap ini, asam sitrat yang telah terbentuk diubah menjadi isositrat dengan bantuan enzim akotinase yang mengandung Fe2+. Tahap III : Isositrat Dehidrogenase Pada tahap ini, berlangsung proses dekarboksilai oksidatif atau perombakan pertama. Isositrat yang terbentuk pada tahap sebelumnya dioksidasi menjadi oksalosuksinat yang terikat enzim oleh enzim isositrat Zdehydrogenase. Pada tahap ini pula, isositrat diubah menjadi αketoglutarat oleh enzim isositrat dehydrogenase dan dibantu NADH. Tahap IV : α-Ketoglutarat Dehidrogenase Kompleks Pada tahap ini terjadi proses pengubahan α-ketoglutarat menjadi suksinil Ko-A oleh enzim α-ketoglutarat dehydrogenase kompleks. Tahap V : Suksinat Thikonase Pada tahap ini, terjadinya konversi suksinil Ko-A menjadi suksinat. Pada tahap ini proses konversi atau pengubahan tidak hanya dibantu oleh enzim namun juga memerlukan Mg2+ dan GPD dengan Pi (fosfat) akan membentuk GTP. GTP inilah yang nantinya akan diubah menjadi ATP. Tahap VI : Suksinat Dehidrogenase Suksinat yang telah dihasilkan pada tahap ini akan di dehidregenase menjadi fumarat dengan bantuan enzim suksinat dehydrogenase. Tahap VII : Hidrasi Pada tahap ini terjadi proses hidrasi, yaitu proses penambahan atom hydrogen pada ikatan ganda karbon (C=C) yang ada pada fumarat sehingga menghasilkan malat. Tahap VIII : Regenerasi Oksaloasetat Pada tahap ini, terjadi pengubahan malat oleh enzim malat dehydrogenase membentuk oksaloasetat. Oksaloasetat ini berperan menangkap Asetil Ko-A sehingga proses siklus kreb dapat berlangsung kembali. Untuk mencukupi kebutuhan energi, siklus krebs harus berlangsung sebnayak dua kali. Hal tersebut karena reaksi oksidasi pada molekul glukosa untuk sekali proses siklus kreb hanya menghasilkan 2 molekul Asetil Ko-A. Dalam proses satu kali siklus krebs, menghasilkan 12 ATP dengan perhitungan 1 molekul GTP yang akan secara diproduksi menjadi ATP; 3 molekul ATP NADH yang akan dioksidasi melalui transport elektron mengahsilkan 3 ATP per molekul; 1 molekul FADH yang akan di oksidasi melalui transport elektron mengahsilkan 2 ATP per molekul; 1 molekul CO2 yang dilepaskan. Sehingga, unntuk dua kali siklus krebs akan dihasilkan energy sebanyak 24 ATP dan molekul CO2. 3. Jalur Pentosa Fosfat Jalur pentosa fosfat merupakan jalur metabolisme alternatif untuk oksidasi glukosa di mana tidak ada ATP yang dihasilkan. Produk utamanya adalah NADPH, suatu pereduksi yang diperlukan dalam beberapa proses anabolisme (untuk biosintesis asam lemak,kolesterol, dan steroid lain) dan ribosa-5 fosfat yang merupakan komponen struktural nukleotida dan asam nukleat (Ribosa untuk biosintesis asam nukleat). Jalur pentosa fosfat merupakan jalur untuk sintesis tiga fosfat pentosa : ribulosa 5 - fosfat, ribose 5 - fosfat, dan xylulose 5 - fosfat. Ribosa 5 – fosfat diperlukan untuk sintesis RNA dan DNA.Jalur pentosa fosfat/heksosa monofosfat menghasilkan NADPH dan ribosa di luar mitokondria.Kepentingan lain jalur pentosa fosfat berlangsung dalam jaringan hepar, lemak, korteks adrenal, tiroid, eritrosit, kelenjar mammae.NADPH juga penting dalam detoksifikasi obat oleh monooksigenase, reduksiglutation. Lintasan pentosa fosfat merupakan jalur alternatif untuk metabolisme glukosa. Lintasan ini tidak menghasilkan ATP, tetapi mempunyai dua fungsi utama, yaitu : a. Produksi NADPH untuk sintesis reduktif seperti biosintesis asam lemak serta steroid. b. Mencegah stress oksidatif dengan mengubah H2O2 menjadi H2O dan jika tidak terdapat NADPH, H2O2akan di ubah menjadi radikal bebas hidroksin yang akan menyerang sel. Pada sel darah merah, kegunaan pertama dari NADPH adalah untuk mereduksi bentuk disulfid dari glutathione menjadi bentuk sulfhydril, reduksi glutathione ini adalah untuk mempertahankan struktur normal dari sel darah merah dan untuk menjaga bentuk hemoglobin dalam bentuk Fe2+.NADPH pada hati dan payudara digunakan untuk biosintesis asam lemak. Reaksi pentosa fosfat terjadi dalam sitosol.Enzim pada lintasan pentosa fosfat seperti pada glikolisis ditemukan di dalam sitosol.Seperti pada glikolisis, oksidasi dicapai lewat reaksi dehidrogenasi, tetapi dalam hal lintasan pentosa fosfat, sebagai akseptor hidrogen digunakan NADP+ dan bukan NAD+.Tidak ada ATP yang digunakan ataupun diproduksi pada jalur ini. Terdapat 2 fase pada pentosa fosfat : 1. Fase oksidatif yang menghasilkan NADPH Pada fase yang pertama, glukosa 6-phosphate menjalani proses dehidroginase dan dekarboksilase untuk memberikan sebuah senyawa pentosa, yaitu ribosa 5-phosphate. 2. Fase nonoksidatif yang menghasilkan prekursor ribosa Pada fase yang kedua, ribulosa 5-fosfat dikonversi kembali menjadi glukosa 6-fosfat oleh serangkaian reaksi yang terutama melibatkan dua enzim yaitu transketolase dan transaldolase. I. Fase Oksidatif yang menghasilkan NADPH Reaksi dehydrogenase glukosa 6-fosfat menjadi 6-fosfoglukonat trejadi lewat pembentukan 6-fosfoglukonolakton yang dikatalisis oleh enzim glukosa-6-fosfat dehydrogenase, suatu enzim yang bergantung pada NADP. Hidrolisis 6-fosfoglukonolakton dilaksankan oleh enzim glukonolakton hydrolase. Tahap oksidasi yang kedua dikatalisis oleh enzim 6-fosfoglukonat dehydrogenase, yang juga memerlukan NADP+ Sebagai akseptor hydrogen. Dekarbosilasi kemudia terjadi dengan pembentukan senyawa ketopentosa yaitu ribulosa 5-fosfat. Reaksi mungkin berlangsung dalam dua tahap melalui intermediate 3-keto-6-fosfoglukonat. II. Fase Nonoksidatif yang menghasilkan prekurso ribose. Pada fase yang kedua, ribulosa 5-fosfat dikonversi kembali menjadi glukosa 6-fosfat oleh serangkaian reaksi yang terutama melibatkan dua enzim yaitu transketolase dan transaldolase. Ribulosa 5fosfat kini berfungsi sebagai substrat bagi dua enzim yang berbeda. Ribulosa 5-fosfat 3-epimerase mengubah konfigurasi disekitar karbon 3 dari ribulosa 5-fosfat, dengan membentuk epimer xilulosa 5-fosfat, yaitu senyawa ketopentosa lainnya. Ribosa 5-fosfat ketoisomerase mengubah ribulosa 5-fosfat menjadi senyawa aldopentosa yang bersesuaian, yaitu ribose 5-fosfat yang merupakan precursor bagi residu ribose yang diperlukan dalam sintesis nukleotida dan asam nukleat. Transketolase memindahkan unut dua-karbon yang terdiri atas karbon aldehid pada gula aldosa. Oleh karena itu, enzim ini mempengaruhi konversi gula pentosa menjadi aldosa dengan berkurangnya dua karbon, dan sekaligus mengonversi gula aldosa menjadi ketosa dengan bertambahnya dua atom karbon. Reaksi tersebut memerlukam vitamin B, yaitu tiamin. Enzim transketolase mengkatalisis proses pemindahan unit dua karbon dari xilulosa 5 fosfat kepada ribulosa 5 fosfat yang menghasilkan ketosa sedoheptulosa 7-fosfat 7 karbon dan aldosa gliseraldehid 3-fosfat. Kedua produk ini kemudian memasuki reaksi lainnya yang dikenal sebagai reaksi transdolasi.Enzim transdolase memungkinka pemindahan moietas dihidroksiaseton 3 karbon (karbon 1-3), dari ketosa sedoheptulosa 7-fosfat kepda aldosa gliseraldehid 3-fosfat untuk membentuk ketosa fruktosa 6fosfat dan aldosa eritrosa 4-fosfat 4 karbon. Kemudian berlangsung reaksi selanjutnya yang sekali lagi melibatkan enzim transketolase dengan xilulosa 5-fosfat berfungsi sebagai donor glikoaldehid. Pada keadaan ini, eritrosit 4-fosfat yang terbentuk di atas bertindak sebagai akseptor, dan hasil reaksinya adalah fruktosa 6fosfat serat gliseraldehid 3-fosfat. 4. Jalur Tranfer Elektron Rantai transpor elektron adalah proses di mana NADH dan [FADH2] dihasilkan selama glikolisis, B-oksidasi, dan proses katabolik lainnyateroksidasi sehingga melepaskan energi dalam bentuk ATP. Mekanisme yang ATP terbentuk dalam ETC disebut phosphorolation kemiosmotik. Produk sampingan dari proses katabolik kebanyakan NADH dan [FADH2] yang merupakan bentuk pengurangan (reduksi). Proses metabolisme menggunakan NADH dan [FADH2] untuk mengangkut elektron dalam bentuk ion hidrida (H-). Elektron ini melewati dari NADH atau [FADH2] untuk pembawa elektron terikat membran yang kemudian diteruskan kepada pembawa elektron lain sampai mereka akhirnya diberikan kepada oksigen yang mengakibatkan produksi air. Saat elektron berlalu dari satu operator elektron ke yang lain ion hidrogen diangkut ke ruang antarmembran di tiga titik-titik tertentu dalam rantai. Pengangkutan ion hidrogen menciptakan konsentrasi yang lebih besar dari ion hidrogen dalam ruang antarmembran dibandingkan dalam matriks yang kemudian dapat digunakan untuk menggerakkan ATP menghasilkan ATP (molekul energi tinggi). Sintase dan Transfer Elektron I (NADH-ubiquinone oxidioreductase): Sebuah protein integral yang menerima elektron dalam bentuk ion hidrida dari NADH dan melewati mereka ke ubiquinone II (suksinat-ubiquinone oxidioreductase alias dehidrogenase suksinat dari siklus TCA): Sebuah protein perifer yang menerima elektron dari suksinat (perantara metabolit siklus TCA) untuk menghasilkan fumarat dan [FADH2]. Dari suksinat elektron diterima oleh [FAD] (kelompok prostetik protein) yang kemudian menjadi [FADH2]. Elektron tersebut kemudian dilewatkan ke ubiquinone. Q (Ubiquinone / ubiquinol): Ubiquinone (bentuk teroksidasi molekul) menerima elektron dari beberapa operator yang berbeda, dari I, II, dehidrogenase Gliserol-3-fosfat, dan ETF. Sekarang bentuk tereduksi (ubiquinol) yang melewati elektronnya pergi ke III. III (ubiquinol-sitokrom c oxidioreductase): Sebuah protein integral yang menerima elektron dari ubiquinol yang kemudian diteruskan ke sitokrom c IV (sitokrom c oksidase): Sebuah protein integral yang yang menerima elektron dari sitokrom c dan mentransfernya ke oksigen untuk menghasilkan air dalam matriks mitokondria. ATP Synthas: Sebuah protein terpisahkan terdiri dari beberapa subunit yang berbeda. Protein ini bertanggung jawab langsung untuk produksi ATP melalui phosphorolation kemiosmotik. Ia menggunakan gradien proton yang dibuat oleh beberapa operator lain di ETC untuk menggerakkan rotor mekanis. Energi dari rotor yang kemudian digunakan untuk phosphorolate ADT menjadi ATP. ETF (Elektron-transfer flavoprotein) dehidrogenase: Ini protein perifer terletak di sisi matriks dari membran dalam merupakan bagian siklus Boksidasi. Elektron dari asil-CoA yang disumbangkan ke flavoprotientransfer elektron yang kemudian ditransfer ke ETF (Elektron-transfer flavoprotein) dehidrogenase dalam bentuk [FADH2]. ETF dehidrogenase kemudian melewati mereka elektron dari [FADH2] untuk ubiquinone dan melalui RTE. Dehydrogenas gliserol-3-fosfat: Ini protein perifer terletak di sisi ruang antarmembran dari membran dalam merupakan bagian dari sistem transportasi gliserol-3-fosfat. Ia menerima sebuah proton dari gliserol-3fosfat ke [FAD] kelompok prostetik yang menghasilkan [FADH2]. Dari [FADH2] elektron tersebut kemudian diberikan kepada ubiquinone dan melalui RTE. 5. Jalur Fosforilasi Oksidatif Fosforilasi menggunakan oksidatif adalah energi yang suatu lintasan dilepaskan metabolisme yang oleh oksidasi nutrien untuk menghasilkan adenosina trifosfat (ATP).Walaupun banyak bentuk kehidupan di bumi menggunakan berbagai jenis nutrien, hampir semuanya menjalankan fosforilasi oksidatif untuk menghasilkan ATP. Lintasan ini sangat umum digunakan karena ia merupakan cara yang sangat efisien untuk melepaskan energi, dibandingkan dengan proses fermentasi alternatif lainnya seperti glikolisis anaerobik. Rantai transpor elektron dalam mitokondria merupakan tempat terjadinya fosforilasi oksidatif pada eukariota. NADH dan suksinat yang dihasilkan pada siklus asam sitrat dioksidasi, melepaskan energi untuk digunakan oleh ATP sintase. Selama fosforilasi oksidatif, elektron ditransfer dari pendonor elektron ke penerima elektron melalui reaksi redoks. Reaksi redoks ini melepaskan energi yang digunakan untuk membentuk ATP. Pada eukariota, reaksi redoks ini dijalankan oleh serangkaian kompleks protein di dalam mitokondria, manakala pada prokariota, protein-protein ini berada di membran dalam sel. Enzim-enzim yang saling berhubungan ini disebut sebagai rantai transpor elektron. Pada eukariota, lima kompleks protein utama terlibat dalam proses ini, manakala pada prokariota, terdapat banyak enzim-enzim berbeda yang terlibat. Energi yang dilepaskan oleh perpindahan elektron melalui rantai transpor elektron ini digunakan untuk mentranspor proton melewati membran dalam mitokondria. Proses ini disebut kemiosmosis. Transpor menghasilkan energi potensial dalam bentuk gradien pH dan potensial listrik di sepanjang membran ini. Energi yang tersimpan dalam bentuk ini dimanfaatkan dengan cara mengijinkan proton mengalir balik melewati membran melalui enzim yang disebut ATP sintase. Enzim ini menggunakan energi seperti ini untuk menghasilkan ATP dari adenosina difosfat (ADP) melalui reaksi fosforilasi. Reaksi ini didorong oleh aliran proton, yang mendorong rotasi salah satu bagian enzim. Walaupun fosforilasi oksidatif adalah bagian vital metabolisme, ia menghasilkan spesi oksigen reaktif seperti superoksida dan hidrogen peroksida. Hal ini dapat mengakibatkan pembentukan radikal bebas, merusak sel tubuh, dan kemungkinan juga menyebabkan penuaan. Enzimenzim yang terlibat dalam lintasan metabolisme ini juga merupakan target dari banyak obat dan racun yang dapat menghambat aktivitas enzim. Tinjauan Transfer Energi Melalui Kemiosmosis Rantai transpor elektron membawa baik proton maupun elektron, mengangkut proton dari donor ke akseptor, dan mengangkut proton melawati membran. Proses ini menggunakan molekul yang larut dan terikat pada molekul transfer. Pada mitokondria, elektron ditransfer dalam ruang antarmembran menggunakan protein transfer elektron sitokrom c yang larut dalam air. Ia hanya mengangkut elektron, dan elektron ini ditransfer menggunakan reduksi dan oksidasi atom besi yang terikat pada protein pada gugus heme strukturnya. Sitokrom c juga ditemukan pada beberapa bakteri, di mana ia berlokasi di dalam ruang periplasma. Dalam membran dalam mitokondria, koenzim Q10 pembawa elektron yang larut dalam lipid membawa baik elektron maupun proton menggunakan siklus redoks. Molekul benzokuinon yang kecil ini sangat hidrofobik, sehingga ia akan berdifusi dengan bebas ke dalam membran. Ketika Q menerima dua elektron dan dua proton, ia menjadi bentuk tereduksi ubikuinol (QH2); ketika QH2 melepaskan dua elektron dan dua proton, ia teroksidasi kembali menjadi bentuk ubikuinon (Q). Akibatnya, jika dua enzim disusun sedemikiannya Q direduksi pada satu sisi membran dan QH2 dioksidasi pada sisi lainnya, ubikuinon akan menggandengkan reaksi ini dan mengulang alik proton melewati membran. Beberapa rantai transpor elektron bakteri menggunakan kuinon yang berbeda, seperti menakuinon, selain ubikuinon Dalam protein, elektron ditransfer antar kofaktor flavin, gugus besi-sulfur, dan sitokrom. Terdapat beberapa jenis gugus besi-sulfur. Jenis paling sederhana yang ditemukan pada rantai transfer elektron terdiri dari dua atom besi yang dihubungkan oleh dua atom sulfur; ini disebut sebagai gugus [2Fe-2S]. Jenis kedua, disebut [4Fe-4S], mengandung sebua kubus empat atom besi dan empat atom sulfur. Tiap-tiap atom pada gugus ini berkoordinasi dengan asam amino, biasanya koordinasi antara atom sulfur dengan sisteina. Kofaktor ion logam menjalani reaksi redoks tanpa mengikat ataupun melepaskan proton, sehingga pada rantai transpor elektron ia hanya berfungsi sebagai pengangkut elektron. Elektron bergerak cukup jauh melalui protein-protein ini dengan cara meloncat disekitar rantai kofaktor ini. Hal ini terjadi melalui penerowongan kuantum, yang terjadi dengan cepat pada jarak yang lebih kecil daripada 1,4×10−9 m. Banyak proses-proses katabolik biokimia, seperti glikolisis, siklus asam sitrat, dan oksidasi beta, menghasilkan koenzim NADH yang tereduksi. Koenzim ini mengandung elektron yang memiliki potensial transfer yang tinggi. Dengan kata lain, ia akan melepaskan energi yang sangat besar semasa oksidasi. Namun, sel tidak akan melepaskan semua energi ini secara bersamaan karena akan menjadi reaksi yang tidak terkontrol. Sebaliknya, elektron dilepaskan dari NADH dan dipindahkan ke oksigen melalui serangkaian enzim yang akan melepaskan sejumlah kecil energi pada tiap-tiap enzim tersebut. Rangkaian enzim yang terdiri dari kompleks I sampai dengan kompleks IV ini disebut sebagai rantai transpor elektron dan ditemukan dalam membran dalam mitokondria. 1. Kompleks I (NADH dehidrogenase) 2. Kompleks II: Suksinat-Q oksidoreduktase. 3. Komplek 3 (Q-sitokrom c oksidoreduktase) 4. Kompleks IV: Sitokrom C Oksidase. 5. Komplek V (ATP sintase) 2. Pengertian Metabolisme Metabolisme adalah semua reaksi kimia yang terjadi di dalam organisme, termasuk yang terjadi di tingkat sel. Dua tujuan utama metabolisme yaitu: Konversi makanan menjadi energi umtuk menjalankan proses seluler. Konversi makanan/bahan bakar menjadi bahan penyusun protein, lipid, asam nukleat dan karbohidrat. BAB III PENUTUP A. KESIMPULAN Setelah mempelajari tentang metabolisme karbohidrat kita dapat menyimpulkan bahwa semua proses kimia yang terjadi di dalam sel suatu organisme disebut metabolisme,dimana terjadi perubahan molekul-moleku kimiawi melalui jalur-jalur tahapan reaksi yang kompleks. Beberapa jalur metabolisme merombak molekul kompleks menjadi molekul sederhana untuk menghasilkan energi (katabolik) melalui proses respirasi dan fermentasi, jalur metabolisme lain yang sebaliknya menggunakan energy untuk membangun molekul kompleks (zat organik) dari molekul yang sederhana berupa zat anorganik (anabolik) melalui proses biosintesis. B. SARAN Dari uraian makalah ini disarankan kepada para pembaca khususnya mahasiswa yang memprogram mata kuliah yang terkait dengan isi makalah ini agar sebaiknya mencari literatur lain baik dari beberapa referensi buku maupun internet agar materi ini dapat dikembangkan lebih luas lagi dengan harapan wawasan dapat bertambah mengenai lima jalur dalam metabolisme karbohidrat. DAFTAR PUSTAKAs Campbel,N.A,. Reece.J.B.,dan Mitchel,L.G.2003.Biologi Edisi Kelima Jilid 2. Jakarta: Erlangga. Horton,R.H.,Moran,L.A.,Ochs,R.S.,Rawan,J.D.,dan Scrimgeour,K.G.2003. Principles of Biochemistry Third Edition New York:Prentice-Hall,Inc. Lehninger,L.A.1993.Dasar-dasar Biokimia.Jakarta:Erlangga. Strayer,L.1995.Biochemistry.W.H.Freeman and Company,New York and Basingstoke.